
The Influence of Symbiosis on the Proteome of the Exaiptasia Endosymbiont Breviolum minutum
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Organisms and Design
2.2. Protein Extraction
2.3. LC-ESI-MS/MS
2.4. Protein Identification and Quantification
2.5. Statistical Analyses
3. Results and Discussion
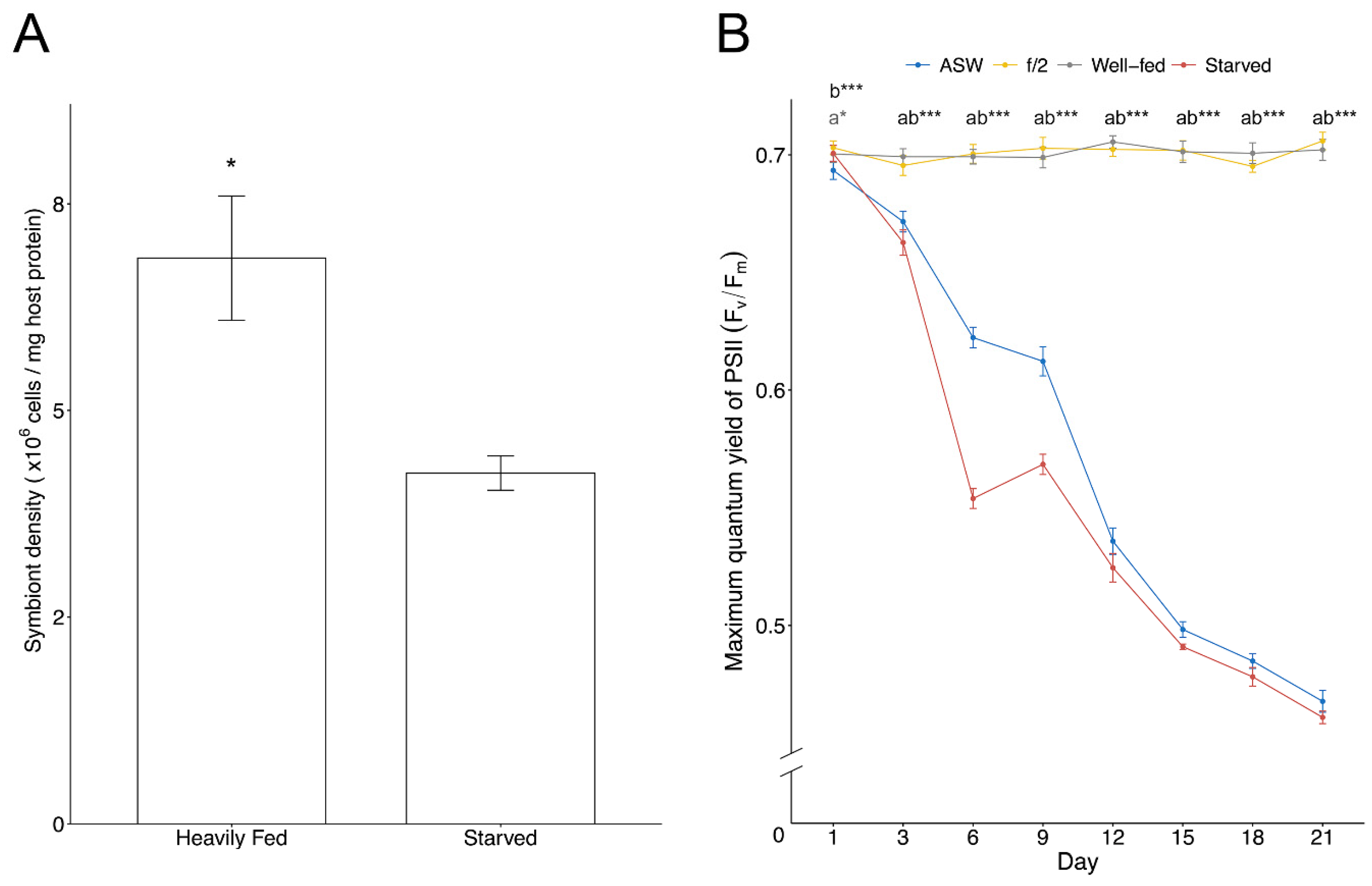
3.1. Endosymbiont Density and Photobiology
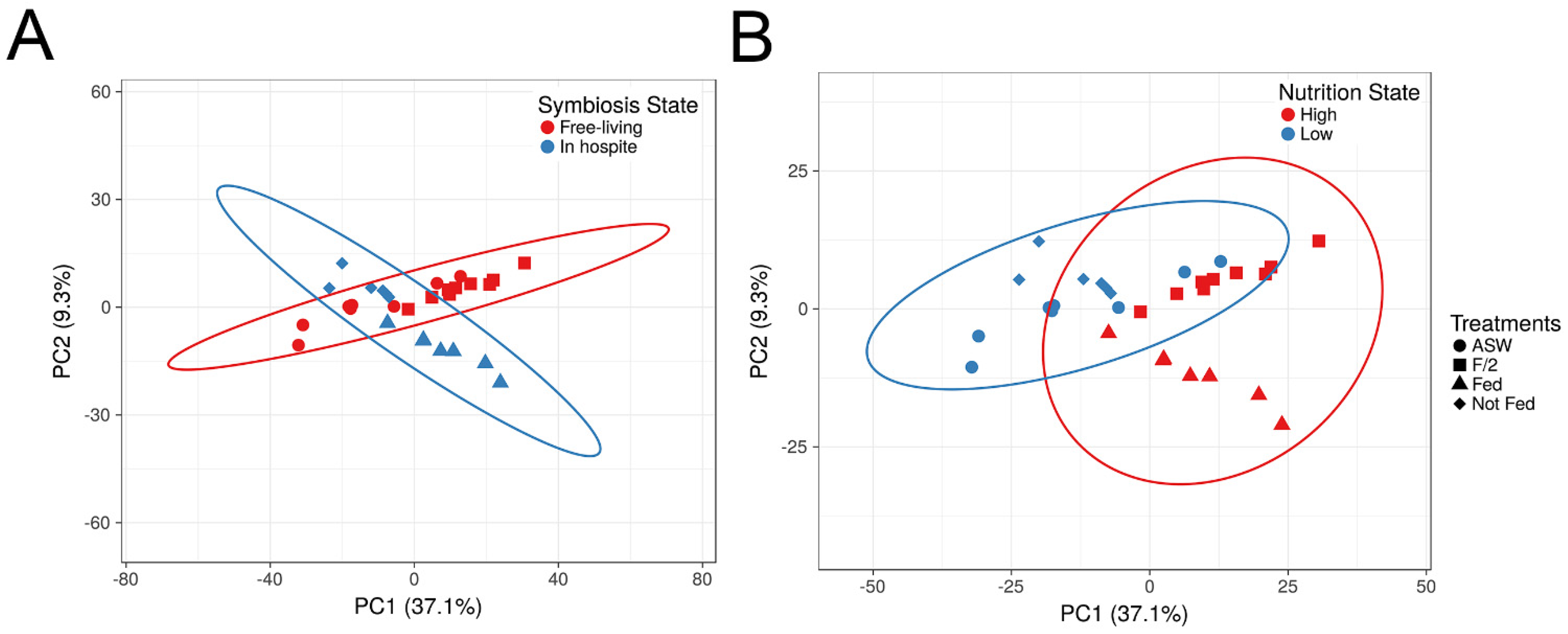
3.2. Proteins Affected by Symbiotic State and Nutritional Regime
3.3. Proteomic Shifts Associated with Symbiotic State
3.3.1. Response to the Symbiosome
3.3.2. Dissolved Inorganic Carbon Transport
3.3.3. Nitrogen Metabolism
3.3.4. Carbohydrate and Fatty Acid Metabolism
3.4. Proteomic Shifts Associated with Nutritional State
3.4.1. Photosynthesis
3.4.2. Oxidative Stress under High Nutrient Regimes
3.4.3. Iron Metabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Veron, J.E.N.; Hoegh-Guldberg, O.; Lenton, T.M.; Lough, J.M.; Obura, D.O.; Pearce-Kelly, P.; Sheppard, C.R.C.; Spalding, M.; Stafford-Smith, M.G.; Rogers, A.D. The Coral Reef Crisis: The Critical Importance of <350 Ppm CO2. Mar. Pollut. Bull. 2009, 58, 1428–1436. [Google Scholar] [CrossRef]
- Muscatine, L.; Porter, J.W. Reef Corals: Mutualistic Symbioses Adapted to Nutrient-Poor Environments. Bioscience 1977, 27, 454–460. [Google Scholar] [CrossRef]
- Matthews, J.L.; Crowder, C.M.; Oakley, C.A.; Lutz, A.; Roessner, U.; Meyer, E.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Optimal Nutrient Exchange and Immune Responses Operate in Partner Specificity in the Cnidarian-Dinoflagellate Symbiosis. Proc. Natl. Acad. Sci. USA 2017, 114, 13194–13199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burriesci, M.S.; Raab, T.K.; Pringle, J.R. Evidence That Glucose Is the Major Transferred Metabolite in Dinoflagellate-Cnidarian Symbiosis. J. Exp. Biol. 2012, 215, 3467–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillyer, K.E.; Dias, D.A.; Lutz, A.; Wilkinson, S.P.; Roessner, U.; Davy, S.K. Metabolite Profiling of Symbiont and Host during Thermal Stress and Bleaching in the Coral Acropora aspera. Coral Reefs 2017, 36, 105–118. [Google Scholar] [CrossRef]
- Dunn, S.R.; Thomas, M.C.; Nette, G.W.; Dove, S.G. A Lipidomic Approach to Understanding Free Fatty Acid Lipogenesis Derived from Dissolved Inorganic Carbon within Cnidarian-Dinoflagellate Symbiosis. PLoS ONE 2012, 7, e46801. [Google Scholar] [CrossRef] [Green Version]
- Hillyer, K.E.; Tumanov, S.; Villas-Bôas, S.; Davy, S.K. Metabolite Profiling of Symbiont and Host during Thermal Stress and Bleaching in a Model Cnidarian-Dinoflagellate Symbiosis. J. Exp. Biol. 2016, 219, 516–527. [Google Scholar] [CrossRef] [Green Version]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580.e6. [Google Scholar] [CrossRef] [Green Version]
- Nitschke, M.R.; Craveiro, S.C.; Brandão, C.; Fidalgo, C.; Serôdio, J.; Calado, A.J.; Frommlet, J.C. Description of Freudenthalidium Gen. Nov. and Halluxium Gen. Nov. to Formally Recognize Clades Fr3 and H as Genera in the Family Symbiodiniaceae (Dinophyceae). J. Phycol. 2020, 56, 923–940. [Google Scholar] [CrossRef]
- Pochon, X.; LaJeunesse, T.C. Miliolidium n. Gen, a New Symbiodiniacean Genus Whose Members Associate with Soritid Foraminifera or Are Free-Living. J. Eukaryot. Microbiol. 2021, 68, e12856. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Wiedenmann, J.; Casado-Amezua, P.; D’Ambra, I.; Turnham, K.E.; Nitschke, M.R.; Oakley, C.A.; Goffredo, S.; Spano, C.A.; Cubillos, V.M.; et al. Revival of Philozoon Geddes for Host-Specialized Dinoflagellates, ‘Zooxanthellae’, in Animals from Coastal Temperate Zones of Northern and Southern Hemispheres. Eur. J. Phycol. 2021, 57, 166–180. [Google Scholar] [CrossRef]
- Wakefield, T.S.; Kempf, S.C. Development of Host- and Symbiont-Specific Monoclonal Antibodies and Confirmation of the Origin of the Symbiosome Membrane in a Cnidarian-Dinoflagellate Symbiosis. Biol. Bull. 2001, 200, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, T.S.; Farmer, M.A.; Kempf, S.C. Revised Description of the Fine Structure of in Situ “Zooxanthellae” Genus Symbiodinium. Biol. Bull. 2000, 199, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Trench, R.K. Microalgal-Invertebrate Symbioses—A Review. Endocytobiosis Cell Res. 1993, 9, 135–175. [Google Scholar]
- Trench, R.K.; Blank, R.J. Symbiodinium microadriaticum Freudenthal, S. goreauii sp. Nov., S. kawagutii sp. Nov. and S. Pilosum sp. Nov.: Gymnodinioid Dinoflagellate Symbionts of Marine Invertebrates. J. Phycol. 1987, 23, 469–481. [Google Scholar] [CrossRef]
- Freudenthal, H.D. Symbiodinium Gen. Nov. and Symbiodinium microadriaticum sp. Nov., a Zooxanthella: Taxonomy, Life Cycle, and Morphology. J. Protozool. 1962, 9, 45–52. [Google Scholar] [CrossRef]
- Muscatine, L. Symbiosis of Hydra and Algae-III. Extracellular Products of the Algae. Comp. Biochem. Physiol. 1965, 16, 77–92. [Google Scholar] [CrossRef]
- Muscatine, L.; Falkowski, P.G.; Dubinsky, Z. Carbon Budgets in Symbiotic Associations BT—Endocytobiology II. In Intracellular Space as Oligogenetic Ecosystem; Schenk, H.E.A., Schwemmler, W., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 1983. [Google Scholar]
- Davy, S.K.; Allemand, D.; Weis, V.M. Cell Biology of Cnidarian-Dinoflagellate Symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76, 229–261. [Google Scholar] [CrossRef] [Green Version]
- Sproles, A.E.; Oakley, C.A.; Matthews, J.L.; Peng, L.; Owen, J.G.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Proteomics Quantifies Protein Expression Changes in a Model Cnidarian Colonised by a Thermally Tolerant but Suboptimal Symbiont. ISME J. 2019, 13, 2334–2345. [Google Scholar] [CrossRef]
- Oakley, C.A.; Durand, E.; Wilkinson, S.P.; Peng, L.; Weis, V.M.; Grossman, A.R.; Davy, S.K. Thermal Shock Induces Host Proteostasis Disruption and Endoplasmic Reticulum Stress in the Model Symbiotic Cnidarian Aiptasia. J. Proteome Res. 2017, 16, 2121–2134. [Google Scholar] [CrossRef]
- Lehnert, E.M.; Mouchka, M.E.; Burriesci, M.S.; Gallo, N.D.; Schwarz, J.A.; Pringle, J.R. Extensive Differences in Gene Expression between Symbiotic and Aposymbiotic Cnidarians. G3 Genes Genomes Genet. 2014, 4, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, E.M.; Burriesci, M.S.; Pringle, J.R. Developing the Anemone Aiptasia as a Tractable Model for Cnidarian-Dinoflagellate Symbiosis: The Transcriptome of Aposymbiotic A. pallida. BMC Genom. 2012, 13, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, R.A.; Palumbi, S.R. Rapid Acclimation Ability Mediated by Transcriptome Changes in Reef-Building Corals. Genome Biol. Evol. 2015, 7, 1602–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasaribu, B.; Weng, L.-C.; Lin, I.-P.; Camargo, E.; Tzen, J.T.C.; Tsai, C.-H.; Ho, S.-L.; Lin, M.-R.; Wang, L.-H.; Chen, C.-S.; et al. Morphological Variability and Distinct Protein Profiles of Cultured and Endosymbiotic Symbiodinium Cells Isolated from Exaiptasia pulchella. Sci. Rep. 2015, 5, 15353. [Google Scholar] [CrossRef] [Green Version]
- Maor-Landaw, K.; van Oppen, M.J.H.; McFadden, G.I. Symbiotic Lifestyle Triggers Drastic Changes in the Gene Expression of the Algal Endosymbiont Breviolum minutum (Symbiodiniaceae). Ecol. Evol. 2020, 10, 451–466. [Google Scholar] [CrossRef] [Green Version]
- Bellantuono, A.J.; Dougan, K.E.; Granados-Cifuentes, C.; Rodriguez-Lanetty, M. Free-Living and Symbiotic Lifestyles of a Thermotolerant Coral Endosymbiont Display Profoundly Distinct Transcriptomes under Both Stable and Heat Stress Conditions. Mol. Ecol. 2019, 28, 5265–5281. [Google Scholar] [CrossRef]
- Chen, H.K.; Song, S.N.; Wang, L.H.; Mayfield, A.B.; Chen, Y.J.; Chen, W.N.U.; Chen, C.S. A Compartmental Comparison of Major Lipid Species in a Coral-Symbiodinium Endosymbiosis: Evidence That the Coral Host Regulates Lipogenesis of Its Cytosolic Lipid Bodies. PLoS ONE 2015, 10, e0132519. [Google Scholar] [CrossRef]
- Tomanek, L. Proteomics to Study Adaptations in Marine Organisms to Environmental Stress. J. Proteom. 2014, 105, 92–106. [Google Scholar] [CrossRef]
- Oakley, C.A.; Newson, G.I.; Peng, L.; Davy, S.K. The Symbiodinium Proteome Response to Thermal and Nutrient Stresses. Plant Cell Physiol. 2022, pcac175. [Google Scholar] [CrossRef]
- Schubert, O.T.; Röst, H.L.; Collins, B.C.; Rosenberger, G.; Aebersold, R. Quantitative Proteomics: Challenges and Opportunities in Basic and Applied Research. Nat. Protoc. 2017, 12, 1289–1294. [Google Scholar] [CrossRef] [Green Version]
- Weis, V.M.; Davy, S.K.; Hoegh-Guldberg, O.; Rodriguez-Lanetty, M.; Pringle, J.R. Cell Biology in Model Systems as the Key to Understanding Corals. Trends Ecol. Evol. 2008, 23, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Grajales, A.; Rodríguez, E. Morphological Revision of the Genus Aiptasia and the Family Aiptasiidae (Cnidaria, Actiniaria, Metridioidea). Zootaxa 2014, 3826, 55–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, C.B.; D’Elia, C.F.; Muller-Parker, G. Host Feeding and Nutrient Sufficiency for Zooxanthellae in the Sea Anemone Aiptasia pallida. Mar. Biol. 1988, 98, 253–262. [Google Scholar] [CrossRef]
- Xiang, T.; Lehnert, E.; Jinkerson, R.E.; Clowez, S.; Kim, R.G.; DeNofrio, J.C.; Pringle, J.R.; Grossman, A.R. Symbiont Population Control by Host-Symbiont Metabolic Interaction in Symbiodiniaceae-Cnidarian Associations. Nat. Commun. 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.T.; Douglas, A.E. Nitrogen Recycling or Nitrogen Conservation in an Alga-Invertebrate Symbiosis? J. Exp. Biol. 1998, 201, 2445–2453. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Springer: Berlin/Heidelberg, Germany, 1975; pp. 29–60. [Google Scholar]
- Baker, A.C. The Symbiosis Ecology of Reef-Building Corals; Paper 3763; University of Miami: Coral Gables, FL, USA, 1999. [Google Scholar]
- Thornhill, D.J.; Lajeunesse, T.C.; Santos, S.R. Measuring RDNA Diversity in Eukaryotic Microbial Systems: How Intragenomic Variation, Pseudogenes, and PCR Artifacts Confound Biodiversity Estimates. Mol. Ecol. 2007, 16, 5326–5340. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Thornhill, D.J. Improved Resolution of Reef-Coral Endosymbiont (Symbiodinium) Species Diversity, Ecology, and Evolution through PsbA Non-Coding Region Genotyping. PLoS ONE 2011, 6, e29013. [Google Scholar] [CrossRef] [Green Version]
- Franklin, E.C.; Stat, M.; Pochon, X.; Putnam, H.M.; Gates, R.D. GeoSymbio: A Hybrid, Cloud-Based Web Application of Global Geospatial Bioinformatics and Ecoinformatics for Symbiodinium–Host Symbioses. Mol. Ecol. Resour. 2012, 12, 369–373. [Google Scholar] [CrossRef]
- Deutsch, E.W.; Csordas, A.; Sun, Z.; Jarnuczak, A.; Perez-Riverol, Y.; Ternent, T.; Campbell, D.S.; Bernal-Llinares, M.; Okuda, S.; Kawano, S.; et al. The ProteomeXchange Consortium in 2017: Supporting the Cultural Change in Proteomics Public Data Deposition. Nucleic Acids Res. 2017, 45, D1100–D1106. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE Database and Related Tools and Resources in 2019: Improving Support for Quantification Data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate Proteome-Wide Label-Free Quantification by Delayed Normalization and Maximal Peptide Ratio Extraction, Termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, J.E.; Baumgarten, S.; Michell, C.T.; Baums, I.B.; LaJeunesse, T.C.; Voolstra, C.R. Gene Expression Variation Resolves Species and Individual Strains among Coral-Associated Dinoflagellates within the Genus Symbiodinium. Genome Biol. Evol. 2016, 8, 665–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgarten, S.; Simakov, O.; Esherick, L.Y.; Liew, Y.J.; Lehnert, E.M.; Michell, C.T.; Li, Y.; Hambleton, E.A.; Guse, A.; Oates, M.E.; et al. The Genome of Aiptasia, a Sea Anemone Model for Coral Symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 11893–11898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Tyanova, S.; Cox, J. Perseus: A Bioinformatics Platform for Integrative Analysis of Proteomics Data in Cancer Research. In Methods in Molecular Biology; von Stechow, L., Ed.; Springer: New York, NY, USA, 2018; Volume 1711, pp. 133–148. ISBN 978-1-4939-7493-1. [Google Scholar]
- Schwämmle, V.; Hagensen, C.E.; Rogowska-Wrzesinska, A.; Jensen, O.N. PolySTest: Robust Statistical Testing of Proteomics Data with Missing Values Improves Detection of Biologically Relevant Features. Mol. Cell. Proteom. 2020, 19, 1396–1408. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2014, 12, 59–60. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Muller-parker, G.; Lee, K.W.; Cook, C.B. Changes in the Ultrastructure of Symbiotic Zooxanthellae (Symbiodinium sp., Dinophyceae) in Fed and Starved Sea Anemones Maintained under High and Low Light. Cell 1996, 32, 987–994. [Google Scholar] [CrossRef]
- Barott, K.L.; Venn, A.A.; Perez, S.O.; Tambutteeé, S.; Tresguerres, M.; Somero, G.N. Coral Host Cells Acidify Symbiotic Algal Microenvironment to Promote Photosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Dunn, S.R.; Schnitzler, C.E.; Weis, V.M. Apoptosis and Autophagy as Mechanisms of Dinoflagellate Symbiont Release during Cnidarian Bleaching: Every Which Way You Lose. Proc. R. Soc. B Biol. Sci. 2007, 274, 3079–3085. [Google Scholar] [CrossRef] [Green Version]
- Weber, S.S.; Ragaz, C.; Hilbi, H. Pathogen Trafficking Pathways and Host Phosphoinositide Metabolism. Mol. Microbiol. 2009, 71, 1341–1352. [Google Scholar] [CrossRef]
- Hilbi, H. Modulation of Phosphoinositide Metabolism by Pathogenic Bacteria. Cell. Microbiol. 2006, 8, 1697–1706. [Google Scholar] [CrossRef]
- Almaguer, C.; Cheng, W.; Nolder, C.; Patton-Vogt, J. Glycerophosphoinositol, a Novel Phosphate Source Whose Transport Is Regulated by Multiple Factors in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 31937–31942. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, P.; Nikiforov, A.; Robinson, B.; Bakalinsky, A.; O’Donnell, A.; Patton-Vogt, J. Elevated Uptake of Glycerophosphoinositol Through the Git1 Permease Causes Cell Growth Inhibition in Saccharomyces cerevisiae. FASEB J. 2017, 31, 781.16. [Google Scholar] [CrossRef]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The Many Roles of the Eukaryotic Elongation Factor 1 Complex. Wiley Interdiscip. Rev. RNA 2012, 3, 543–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, K.L.; Jarocki, V.M.; Charles, I.G.; Djordjevic, S.P. The Diverse Functional Roles of Elongation Factor Tu (Ef-Tu) in Microbial Pathogenesis. Front. Microbiol. 2019, 10, 2351. [Google Scholar] [CrossRef] [PubMed]
- De Medina-Redondo, M.; Arnáiz-Pita, Y.; Fontaine, T.; Del Rey, F.; Latgé, J.P.; De Aldana, C.R.V. The β-1,3-Glucanosyltransferase Gas4p Is Essential for Ascospore Wall Maturation and Spore Viability in Schizosaccharomyces pombe. Mol. Microbiol. 2008, 68, 1283–1299. [Google Scholar] [CrossRef] [PubMed]
- Colley, N.J.; Trench, R.K. Selectivity in Phagocytosis and Persistence of Symbiotic Algae in the Scyphistoma Stage of the Jellyfish Cassiopeia xamachana. Proc. R. Soc. Lond. Ser. B Contain. Pap. A Biol. Character. R. Soc. 1983, 219, 61–82. [Google Scholar] [CrossRef]
- Schoenberg, D.A.; Trench, R.K.; Smith, D.C. Genetic Variation in Symbiodinium (= Gymnodinium) Microadriaticum Freudenthal, and Specificity in Its Symbiosis with Marine Invertebrates. III. Specificity and Infectivity of Symbiodinium Microadriaticum. Proc. R. Soc. Lond. B Biol. Sci. 1980, 207, 445–460. [Google Scholar] [CrossRef]
- Furla, P.; Galgani, I.; Durand, I.; Allemand, D. Sources and Mechanisms of Inorganic Carbon Transport for Coral Calcification and Photosynthesis. J. Exp. Biol. 2000, 203, 3445–3457. [Google Scholar] [CrossRef]
- Oakley, C.A.; Ameismeier, M.F.; Peng, L.; Weis, V.M.; Grossman, A.R.; Davy, S.K. Symbiosis Induces Widespread Changes in the Proteome of the Model Cnidarian Aiptasia. Cell. Microbiol. 2016, 18, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Weis, V.M. The Induction of Carbonic Anhydrase in the Symbiotic Sea Anemone Aiptasia pulchella. Biol. Bull. 1991, 180, 496–504. [Google Scholar] [CrossRef]
- Rai, A.K.; Chen, T.; Moroney, J. Mitochondrial Carbonic Anhydrases Are Needed for Optimal Photosynthesis at Low CO2 Levels in Chlamydomonas. Plant Physiol. 2021, 187, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, A.; Tambutté, É.; Tambutté, S.; Allemand, D.; Zoccola, D. Symbiosis-Dependent Gene Expression in Coral-Dinoflagellate Association: Cloning and Characterization of a P-Type H+-ATPase Gene. Proc. R. Soc. B Biol. Sci. 2010, 277, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mies, M.; Voolstra, C.R.; Castro, C.B.; Pires, D.O.; Calderon, E.N.; Sumida, P.Y.G. Expression of a Symbiosis-Specific Gene in Symbiodinium Type A1 Associated with Coral, Nudibranch and Giant Clam Larvae. R. Soc. Open Sci. 2017, 4, 170253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.J.; Yellowlees, D. Inorganic Nitrogen Uptake by Symbiotic Marine Cnidarians: A Critical Review. Proc.—R. Soc. Lond. B 1989, 237, 109–125. [Google Scholar] [CrossRef]
- Rädecker, N.; Raina, J.B.; Pernice, M.; Perna, G.; Guagliardo, P.; Kilburn, M.R.; Aranda, M.; Voolstra, C.R. Aiptasia as a Model to Study Metabolic Diversity and Specificity in Cnidarian-Dinoflagellate Symbioses. bioRxiv 2017, 9, 214. [Google Scholar]
- Grover, R.; Maguer, J.-F.F.; Allemand, D.; Ferrier-Pagès, C. Uptake of Dissolved Free Amino Acids by the Scleractinian Coral Stylophora pistillata. J. Exp. Biol. 2008, 211, 860–865. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Fixter, L.M.; Davies, P.S. Ammonium Metabolism in the Symbiotic Sea Anemone Anemonia viridis. Hydrobiologia 2001, 461, 25–35. [Google Scholar] [CrossRef]
- Grover, R.; Maguer, J.F.; Reynaud-Vaganay, S.; Ferrier-Pagès, C. Uptake of Ammonium by the Scleractinian Coral Stylophora pistillata: Effect of Feeding, Light, and Ammonium Concentrations. Limnol. Oceanogr. 2002, 47, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Pernice, M.; Meibom, A.; Van Den Heuvel, A.; Kopp, C.; Domart-Coulon, I.; Hoegh-Guldberg, O.; Dove, S. A Single-Cell View of Ammonium Assimilation in Coral-Dinoflagellate Symbiosis Microbe-Microbe and Microbe-Host Interactions. ISME J. 2012, 6, 1314–1324. [Google Scholar] [CrossRef] [Green Version]
- Yellowlees, D.; Rees, T.; Fitt, W. Effect of Ammonium-Supplemented Seawater on Glutamine Synthetase and Glutamate Dehydrogenase Activities in Host Tissue and Zooxanthellae of Pocillopora damicornis and on Ammonium Uptake Rates of the Zooxanthellae. Pac. Sci. 1994, 48, 291–295. [Google Scholar]
- Wang, J.T.; Douglas, A.E. Essential Amino Acid Synthesis and Nitrogen Recycling in an Alga-Invertebrate Symbiosis. Mar. Biol. 1999, 135, 219–222. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Dubinsky, Z.; Muscatine, L.; McCloskey, L. Population Control in Symbiotic Corals. Bioscience 1993, 43, 606–611. [Google Scholar] [CrossRef]
- Rahav, O.; Dubinsky, Z.; Achituv, Y.; Falkowski, P.G. Ammonium Metabolism in the Zooxanthellate Coral, Stylophora pistillata. Proc. R. Soc. Lond. B Biol. Sci. 1989, 236, 325–337. [Google Scholar] [CrossRef]
- Kopp, C.; Pernice, M.; Domart-Coulon, I.; Djediat, C.; Spangenberg, J.E.; Alexander, D.T.L.; Hignette, M.; Meziane, T.; Meibom, A. Highly Dynamic Cellular-Level Response of Symbiotic Coral to a Sudden Increase in Environmental Nitrogen. mBio 2013, 4, e00052-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howden, A.J.M.; Preston, G.M. Nitrilase Enzymes and Their Role in Plant-Microbe Interactions. Microb. Biotechnol. 2009, 2, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; De Luca, D.; Amoroso, M.; Benfatto, S.; Maestri, S.; Racioppi, C.; Esposito, F.; Ianora, A. New Molecular Insights on the Response of the Green Alga Tetraselmis suecica to Nitrogen Starvation. Sci. Rep. 2019, 9, 3336. [Google Scholar] [CrossRef] [Green Version]
- Shinzato, C.; Inoue, M.; Kusakabe, M. A Snapshot of a Coral “Holobiont”: A Transcriptome Assembly of the Scleractinian Coral, Porites, Captures a Wide Variety of Genes from Both the Host and Symbiotic Zooxanthellae. PLoS ONE 2014, 9, e85182. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, R.; Wakayama, M.; Nagano, Y.; Kawamura, T.; Sakai, K.; Moriguchi, M. Overexpression of Salt-Tolerant Glutaminase from Micrococcus Luteus K-3 in Escherichia coli and Its Purification. Protein. Expr. Purif. 1999, 15, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Nakamura, J.; Straub, K.; Wu, J.; Lou, L. The Glutamine Hydrolysis Function of Human GMP Synthetase. Identification of an Essential Active Site Cysteine. J. Biol. Chem. 1995, 270, 23450–23455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, N.J.; Von Schaewen, A. The Oxidative Pentose Phosphate Pathway: Structure and Organisation. Curr. Opin. Plant Biol. 2003, 6, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.C.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.M.; Krüger, A.; Tauqeer Alam, M.; et al. The Return of Metabolism: Biochemistry and Physiology of the Pentose Phosphate Pathway. Biol. Rev. 2015, 90, 927–963. [Google Scholar] [CrossRef]
- Huang, N.; Ledyard Stebbins, G.; Rodriguez, R.L. Classification and Evolution of α-Amylase Genes in Plants. Proc. Natl. Acad. Sci. USA 1992, 89, 7526–7530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labourel, A.; Frandsen, K.E.H.; Zhang, F.; Brouilly, N.; Grisel, S.; Haon, M.; Ciano, L.; Ropartz, D.; Fanuel, M.; Martin, F.; et al. A Fungal Family of Lytic Polysaccharide Monooxygenase-like Copper Proteins. Nat. Chem. Biol. 2020, 16, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Nutt, A.; Sild, V.; Pettersson, G.; Johansson, G. A Mean for Functional Classification of Cellulases. Eur. J. Biochem. 1998, 258, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Markell, D.; Trench, R.; Iglesias-prieto, R. Macromolecules Associated with the Cell Walls of Symbiotic Dinoflagellates. Symbiosis 1992, 12, 19–31. [Google Scholar]
- Wang, L.H.; Lee, H.H.; Fang, L.S.; Mayfield, A.B.; Chen, C.S. Fatty Acid and Phospholipid Syntheses Are Prerequisites for the Cell Cycle of Symbiodinium and Their Endosymbiosis within Sea Anemones. PLoS ONE 2013, 8, e72486. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.N.U.; Kang, H.J.; Weis, V.M.; Mayfield, A.B.; Jiang, P.L.; Fang, L.S.; Chen, C.S. Diel Rhythmicity of Lipid-Body Formation in a Coral-Symbiodinium Endosymbiosis. Coral Reefs 2012, 31, 521–534. [Google Scholar] [CrossRef]
- Wang, L.H.; Chen, H.K.; Jhu, C.S.; Cheng, J.O.; Fang, L.S.; Chen, C.S. Different Strategies of Energy Storage in Cultured and Freshly Isolated Symbiodinium sp. J. Phycol. 2015, 51, 1127–1136. [Google Scholar] [CrossRef]
- Matthews, J.L.; Oakley, C.A.; Lutz, A.; Hillyer, K.E.; Roessner, U.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Partner Switching and Metabolic Flux in a Model Cnidarian–Dinoflagellate Symbiosis. Proc. R. Soc. B Biol. Sci. 2018, 285, 20182336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosset, S.; Koster, G.; Brandsma, J.; Hunt, A.N.; Postle, A.D.; D’Angelo, C. Lipidome Analysis of Symbiodiniaceae Reveals Possible Mechanisms of Heat Stress Tolerance in Reef Coral Symbionts. Coral Reefs 2019, 38, 1241–1253. [Google Scholar] [CrossRef] [Green Version]
- Beedessee, G.; Hisata, K.; Roy, M.C.; Satoh, N.; Shoguchi, E. Multifunctional Polyketide Synthase Genes Identified by Genomic Survey of the Symbiotic Dinoflagellate, Symbiodinium minutum. BMC Genom. 2015, 16, 941. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Barua, A.; Ruvindy, R.; Savela, H.; Ajani, P.A.; Murray, S.A. The Genetic Basis of Toxin Biosynthesis in Dinoflagellates. Microorganisms 2019, 7, 222. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Buensanteai, N.; Moran-Diez, M.E.; Druzhinina, I.S.; Kenerley, C.M. Functional Analysis of Non-Ribosomal Peptide Synthetases (NRPSs) in Trichoderma Virens Reveals a Polyketide Synthase (PKS)/NRPS Hybrid Enzyme Involved in the Induced Systemic Resistance Response in Maize. Microbiology 2012, 158, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Muñoz, J.C.; Collemare, J. Evolutionary Histories of Type III Polyketide Synthases in Fungi. Front. Microbiol. 2020, 10, 3018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, T.; Hambleton, E.A.; Denofrio, J.C.; Pringle, J.R.; Grossman, A.R. Isolation of Clonal Axenic Strains of the Symbiotic Dinoflagellate Symbiodinium and Their Growth and Host Specificity. J. Phycol. 2013, 49, 447–458. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, H.; Orf, G.S.; Lu, Y.; Xu, W.; Harrington, L.B.; Liu, H.; Lo, C.S.; Blankenship, R.E. Evidence of Functional Trimeric Chlorophyll a/C2-Peridinin Proteins in the Dinoflagellate Symbiodinium. Biochim. Biophys. Acta (BBA)—Bioenerg. 2014, 1837, 1904–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubinsky, Z.; Stambler, N.; Ben-Zion, M.; McCloskey, L.R.; Muscatine, L.; Falkowski, P.G. The Effect of External Nutrient Resources on the Optical Properties and Photosynthetic Efficiency of Stylophora pistillata. Proc. R. Soc. B Biol. Sci. 1990, 239, 231–246. [Google Scholar] [CrossRef]
- Roberty, S.; Béraud, E.; Grover, R.; Ferrier-Pagès, C. Coral Productivity Is Co-Limited by Bicarbonate and Ammonium Availability. Microorganisms 2020, 8, 640. [Google Scholar] [CrossRef] [PubMed]
- Borell, E.M.; Bischof, K. Feeding Sustains Photosynthetic Quantum Yield of a Scleractinian Coral during Thermal Stress. Oecologia 2008, 157, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Fitt, W.K.; Cook, C.B. The Effects of Feeding or Addition of Dissolved Inorganic Nutrients in Maintaining the Symbiosis between Dinoflagellates and a Tropical Marine Cnidarian. Mar. Biol. 2001, 139, 507–517. [Google Scholar] [CrossRef]
- Venn, A.A.; Loram, J.E.; Douglas, A.E. Photosynthetic Symbioses in Animals. J. Exp. Bot. 2008, 59, 1069–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wietheger, A.; Starzak, D.E.; Gould, K.S.; Davy, S.K. Differential ROS Generation in Response to Stress in Symbiodinium spp. Biol. Bull. 2018, 234, 11–21. [Google Scholar] [CrossRef] [PubMed]
- McGinty, E.S.; Pieczonka, J.; Mydlarz, L.D. Variations in Reactive Oxygen Release and Antioxidant Activity in Multiple Symbiodinium Types in Response to Elevated Temperature. Microb. Ecol. 2012, 64, 1000–1007. [Google Scholar] [CrossRef]
- Roberty, S.; Furla, P.; Plumier, J.C. Differential Antioxidant Response between Two Symbiodinium Species from Contrasting Environments. Plant Cell Environ. 2016, 39, 2713–2724. [Google Scholar] [CrossRef]
- Krueger, T.; Becker, S.; Pontasch, S.; Dove, S.; Hoegh-Guldberg, O.; Leggat, W.; Fisher, P.L.; Davy, S.K. Antioxidant Plasticity and Thermal Sensitivity in Four Types of Symbiodinium sp. J. Phycol. 2014, 50, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.U.; Szabó, M.; Deák, Z.; Sass, L.; Larkum, A.; Ralph, P.; Vass, I. Symbiodinium sp. Cells Produce Light-Induced Intra- and Extracellular Singlet Oxygen, Which Mediates Photodamage of the Photosynthetic Apparatus and Has the Potential to Interact with the Animal Host in Coral Symbiosis. New Phytol. 2016, 212, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrier-Pagès, C.; Gattuso, J.P.; Dallot, S.; Jaubert, J. Effect of Nutrient Enrichment on Growth and Photosynthesis of the Zooxanthellate Coral Stylophora pistillata. Coral Reefs 2000, 19, 103–113. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding Oxidative Stress and Antioxidant Functions to Enhance Photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Davy, S.K.; Withers, K.J.T.; Hinde, R. Effects of Host Nutritional Status and Seasonality on the Nitrogen Status of Zooxanthellae in the Temperate Coral Plesiastrea versipora (Lamarck). J. Exp. Mar. Biol. Ecol. 2006, 335, 256–265. [Google Scholar] [CrossRef]
- Segovia, M. Programmed Cell Death in Dinoflagellates. In Programmed Cell Death in Protozoa; Springer: Berlin/Heidelberg, Germany, 2008; pp. 126–142. [Google Scholar]
- Dunn, S.R.; Thomason, J.C.; Le Tissier, M.D.A.; Bythell, J.C. Heat Stress Induces Different Forms of Cell Death in Sea Anemones and Their Endosymbiotic Algae Depending on Temperature and Duration. Cell Death Differ. 2004, 11, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Franklin, D.J.; Hoegh-Guldberg, O.; Jones, R.J.; Berges, J.A. Cell Death and Degeneration in the Symbiotic Dinoflagellates of the Coral Stylophora pistillata during Bleaching. Mar. Ecol. Prog. Ser. 2004, 272, 117–130. [Google Scholar] [CrossRef]
- Richier, S.; Sabourault, C.C.; Courtiade, J.; Zucchini, N.; Allemand, D.; Furla, P. Oxidative Stress and Apoptotic Events during Thermal Stress in the Symbiotic Sea Anemone, Anemonia viridis. FEBS J. 2006, 273, 4186–4198. [Google Scholar] [CrossRef]
- Watanabe, N.; Lam, E. Arabidopsis Metacaspase 2d Is a Positive Mediator of Cell Death Induced during Biotic and Abiotic Stresses. Plant J. 2011, 66, 969–982. [Google Scholar] [CrossRef]
- Hawkins, T.D.; Bradley, B.J.; Davy, S.K. Nitric Oxide Mediates Coral Bleaching through an Apoptotic-like Cell Death Pathway: Evidence from a Model Sea Anemone-Dinoflagellate Symbiosis. FASEB J. 2013, 27, 4790–4798. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, L.; Chen, M.; Chen, J. The Acute Transcriptomic Response of Coral-Algae Interactions to PH Fluctuation. Mar. Genom. 2018, 42, 32–40. [Google Scholar] [CrossRef]
- Gierz, S.L.; Forêt, S.; Leggat, W. Transcriptomic Analysis of Thermally Stressed Symbiodinium Reveals Differential Expression of Stress and Metabolism Genes. Front. Plant Sci. 2017, 8, 271. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A.; Evans, M.C.W.; Korb, R.E. The Role of Trace Metals in Photosynthetic Electron Transport in O2-Evolving Organisms. Photosynth. Res. 1999, 60, 111–150. [Google Scholar] [CrossRef]
- Sunda, W.G. Feedback Interactions between Trace Metal Nutrients and Phytoplankton in the Ocean. Front. Microbiol. 2012, 3, 204. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, I.B.; Lin, S.; Ho, J.; Ho, T.Y. Effects of Trace Metal Concentrations on the Growth of the Coral Endosymbiont Symbiodinium kawagutii. Front. Microbiol. 2016, 7, 82. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.B.; Ho, T.Y. Trace Metal Requirements and Interactions in Symbiodinium kawagutii. Front. Microbiol. 2018, 9, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, H.G.; Rodriguez, I.B.; LaJeunesse, T.C.; Ho, T.Y. Endosymbiotic Dinoflagellates Pump Iron: Differences in Iron and Other Trace Metal Needs among the Symbiodiniaceae. Coral Reefs 2020, 39, 915–927. [Google Scholar] [CrossRef]
- Groussman, R.D.; Parker, M.S.; Armbrust, E.V. Diversity and Evolutionary History of Iron Metabolism Genes in Diatoms. PLoS ONE 2015, 10, e0129081. [Google Scholar] [CrossRef]
- Herbik, A.; Bölling, C.; Buckhout, T.J. The Involvement of a Multicopper Oxidase in Iron Uptake by the Green Algae Chlamydomonas reinhardtii. Plant Physiol. 2002, 130, 2039–2048. [Google Scholar] [CrossRef] [Green Version]
- Ferrier-Pagès, C.; Sauzéat, L.; Balter, V. Coral Bleaching Is Linked to the Capacity of the Animal Host to Supply Essential Metals to the Symbionts. Glob. Chang. Biol. 2018, 24, 3145–3157. [Google Scholar] [CrossRef]
- Arosio, P.; Elia, L.; Poli, M. Ferritin, Cellular Iron Storage and Regulation. IUBMB Life 2017, 69, 414–422. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| In vs. Ex hospite Fold Change | Uniprot ID | Uniprot Protein Names |
|---|---|---|
| Unique, in hospite | A0A1Q9DR34 | Polyphosphatidylinositol phosphatase (INP53) |
| Unique, in hospite | A0A1Q9F6G5 | Nitrilase |
| Unique, in hospite | Q59Q30 | Glycerophosphoinositol permease 1 (GIT1) |
| Unique, in hospite | Q04634 | Elongation factor 1-alpha (eEF1) |
| Unique, in hospite | Q06135 | 1,3-beta-glucanosyltransferase (GAS2) |
| 3.44 | B0Y8K2 | 1,4-beta-D-glucan cellobiohydrolase B (CBHB) |
| 1.78 | A0A1Q9CUD0 | Putative glucose-6-phosphate 1-epimerase |
| 1.76 | Q9SAU2 | Ribulose-5-phosphate-3-epimerase (R5P3E) |
| 1.16 | B4R9R7 | Guanosine monophosphate synthetase (GMP) |
| −0.92 | P11029 | Acetyl-CoA carboxylase (ACC) |
| −1.03 | Q9Y7Y7 | 1,3-beta-glucanosyltransferase (GAS4) |
| −1.10 | Q54WR9 | Type-3 glutamine synthetase |
| −1.49 | Q12613 | Glutamine synthetase (GS) |
| −1.50 | Q00955 | Acetyl-CoA carboxylase (ACC) |
| −1.56 | A0A1Q9CUK2 | GDP-mannose transporter GONST5 |
| −2.04 | Q9LV03 | Glutamate synthase 1, chloroplastic |
| −3.21 | Q76NM6 | V-type proton ATPase catalytic subunit A |
| −3.90 | A0A1Q9D9X1 | Carbonic anhydrase 2 |
| −3.95 | Q9I262 | Carbonic anhydrase |
| Unique, ex hospite | P74572 | Pyrroline-5-carboxylate reductase (P5CR) |
| Unique, ex hospite | Q9LV03 | Glutamate synthase 1, chloroplastic |
| Unique, ex hospite | A5CZ28 | Gamma-glutamyl phosphate reductase (GPR) |
| Unique, ex hospite | A0A1Q9CS65 | Carbonic anhydrase 2 |
| Unique, ex hospite | Q9I262 | Carbonic anhydrase |
| Unique, ex hospite | G4N296 | Highly reducing polyketide synthase 19 (PKS19) |
| Unique, ex hospite | P9WQE6 | Phenolphthiocerol/phthiocerol polyketide synthase (PpsA) |
| Unique, ex hospite | P21567 | Alpha-amylase |
| Unique, ex hospite | G2Q9T3 | Polysaccharide monooxygenase |
| High vs. Low Nutrient Regime Fold Change | Uniprot ID | Uniprot Protein Names |
|---|---|---|
| Unique, High | A0A1Q9CJD4 | Glutathione S-transferase |
| Unique, High | A0A1Q9CL40 | Multicopper oxidase (MCO) |
| 3.54 | O09452 | Glyceraldehyde-3-phosphate dehydrogenase |
| 3.01 | P26302 | Phosphoribulokinase (PRK) |
| 2.88 | P00110 | Cytochrome c6 (Cytochrome c-553) |
| 2.87 | Q40296 | Chlorophyll a-chlorophyll c2-peridinin-protein (apcPC) |
| 2.84 | P00110 | Cytochrome c6 (Cytochrome c-553) |
| 2.78 | Q40297 | Chlorophyll a-chlorophyll c2-peridinin-protein (apcPC) |
| 2.77 | Q95AG0 | Cytochrome f |
| 2.76 | A0T0C6 | Cytochrome c-550 (Cytochrome c550) |
| 2.68 | Q9XQV2 | Photosystem I P700 chlorophyll a apoprotein A2 |
| 2.58 | Q85FP8 | Photosystem I reaction centre subunit XI |
| 2.53 | Q9SDM1 | Chlorophyll a-b binding protein |
| 2.53 | Q00598 | Ferredoxin--NADP reductase |
| 2.5 | P55738 | Caroteno-chlorophyll a-c-binding protein |
| 2.39 | Q41406 | Ribulose bisphosphate carboxylase (RuBisCO) |
| 2.35 | A0A1Q9EQB8 | Metacaspase-1B |
| 2.34 | P49472 | Photosystem II CP43 reaction centre protein |
| 2.32 | P25851 | Fructose-1,6-bisphosphatase 1, (FBPase1) (ELECTRON FLOW 1) |
| 2.31 | P54375 | Superoxide dismutase (SOD) |
| 2.23 | A0T0T0 | Photosystem II D2 protein |
| 1.69 | Q9MSC2 | Photosystem II protein D1 |
| 1.64 | P20136 | Glutathione S-transferase 2 (GST-CL2) |
| 1.47 | Q7XJ02 | Probable L-ascorbate peroxidase 7 (APX) |
| 1.39 | P46434 | Glutathione S-transferase 1 |
| 1.21 | P42770 | Glutathione reductase, chloroplastic (GR) |
| 0.92 | P46429 | Glutathione S-transferase 2 |
| −1.07 | P42577 | Soma ferritin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mashini, A.G.; Oakley, C.A.; Beepat, S.S.; Peng, L.; Grossman, A.R.; Weis, V.M.; Davy, S.K. The Influence of Symbiosis on the Proteome of the Exaiptasia Endosymbiont Breviolum minutum. Microorganisms 2023, 11, 292. https://doi.org/10.3390/microorganisms11020292
Mashini AG, Oakley CA, Beepat SS, Peng L, Grossman AR, Weis VM, Davy SK. The Influence of Symbiosis on the Proteome of the Exaiptasia Endosymbiont Breviolum minutum. Microorganisms. 2023; 11(2):292. https://doi.org/10.3390/microorganisms11020292
Chicago/Turabian StyleMashini, Amirhossein Gheitanchi, Clinton A. Oakley, Sandeep S. Beepat, Lifeng Peng, Arthur R. Grossman, Virginia M. Weis, and Simon K. Davy. 2023. "The Influence of Symbiosis on the Proteome of the Exaiptasia Endosymbiont Breviolum minutum" Microorganisms 11, no. 2: 292. https://doi.org/10.3390/microorganisms11020292