
Genetic, Antigenic, and Pathobiological Characterization of H9 and H6 Low Pathogenicity Avian Influenza Viruses Isolated in Vietnam from 2014 to 2018
 , , , , and
, , , , and 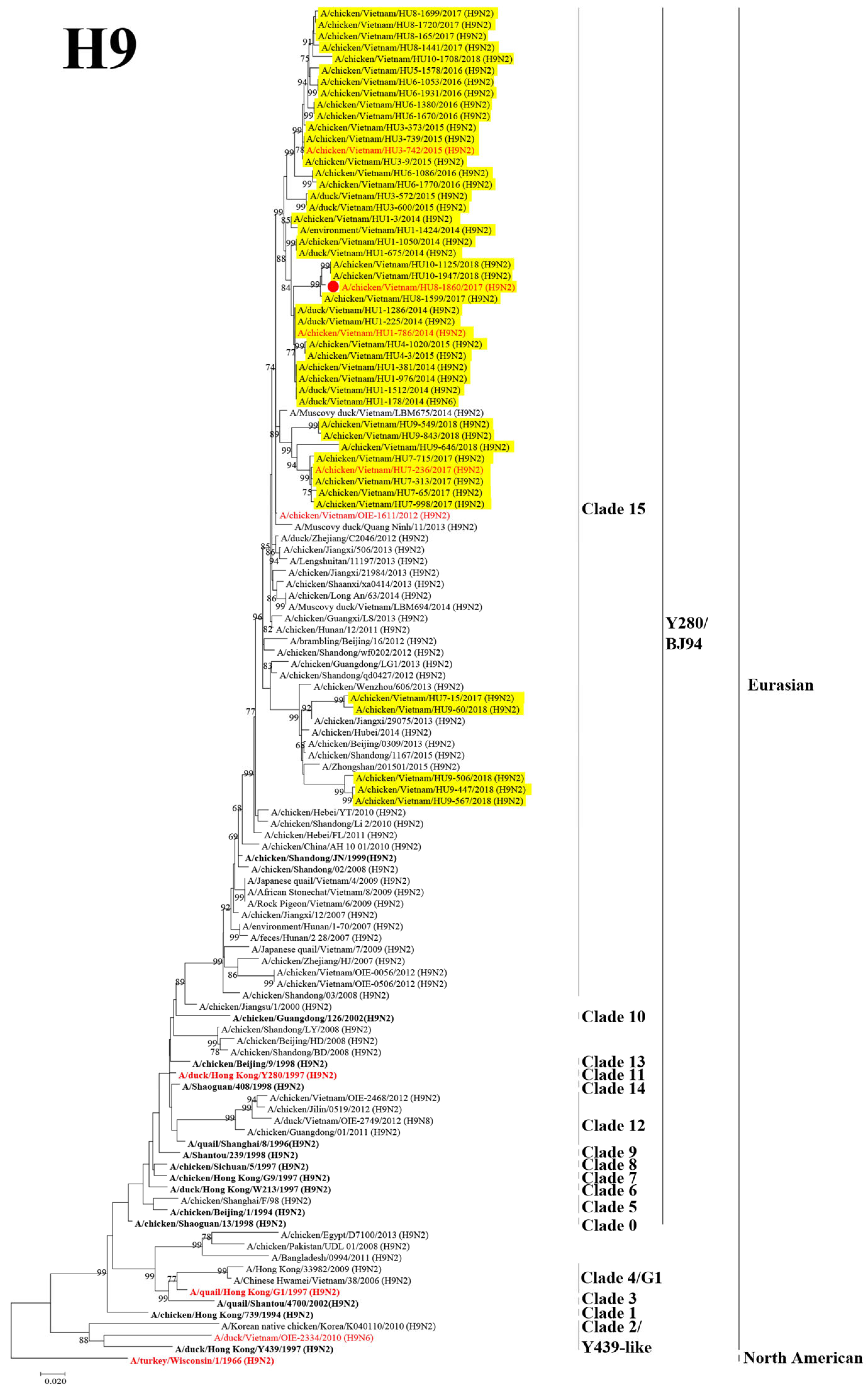{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification of AIVs
2.3. Sequencing and Phylogenetic Analysis
2.4. Cross-HI Test and Antigenic Cartography
2.5. Isolation and Identification of E. coli
2.6. Animal Experiments
2.7. Virus Titration
2.8. Statistical Analysis
3. Results
3.1. Identification of the AIVs Circulating in Poultry
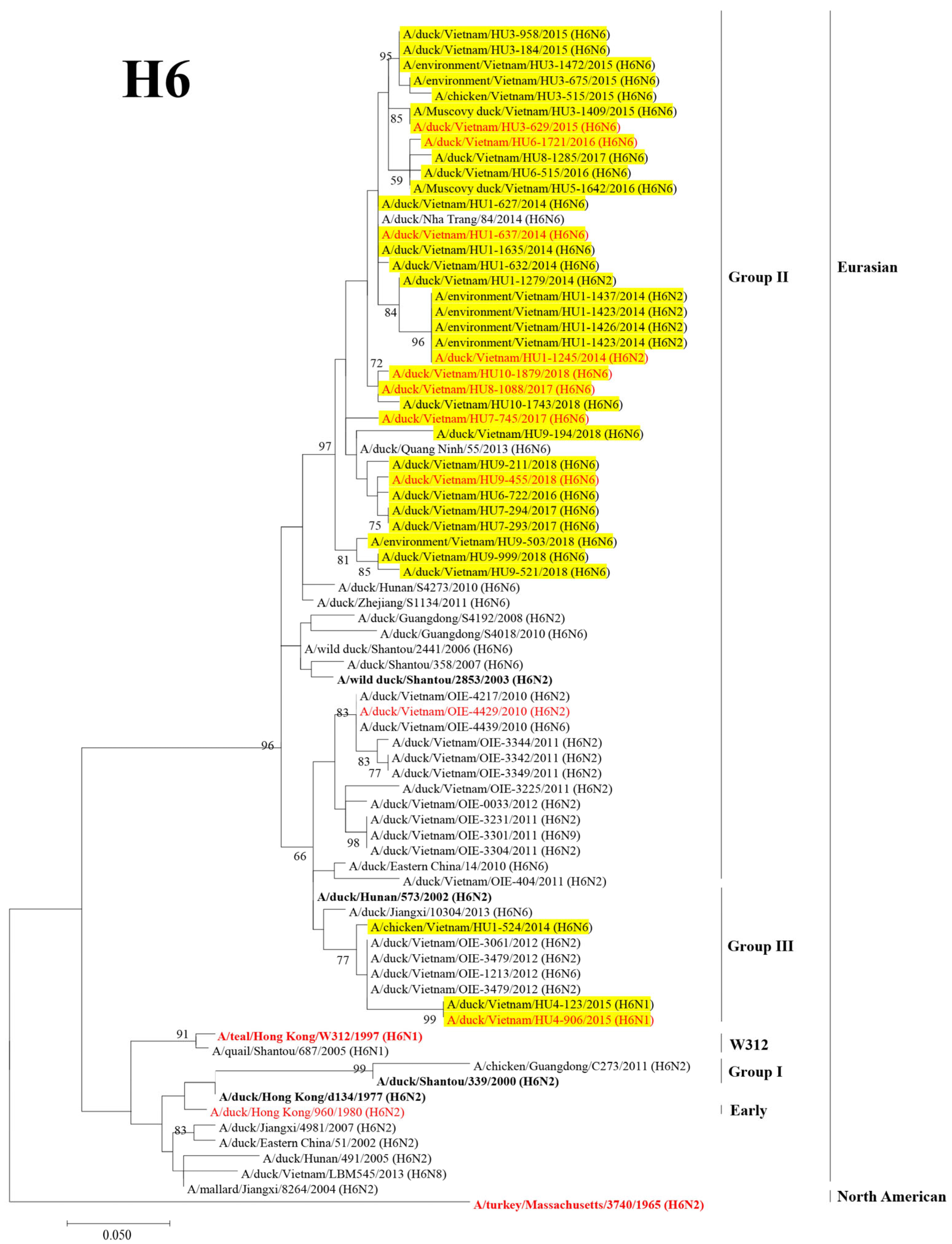
3.2. Phylogenetic Analysis of the HA and NA Genes of H9 and H6 LPAIVs
3.3. Genotyping the Isolated H9 and H6 LPAIVs
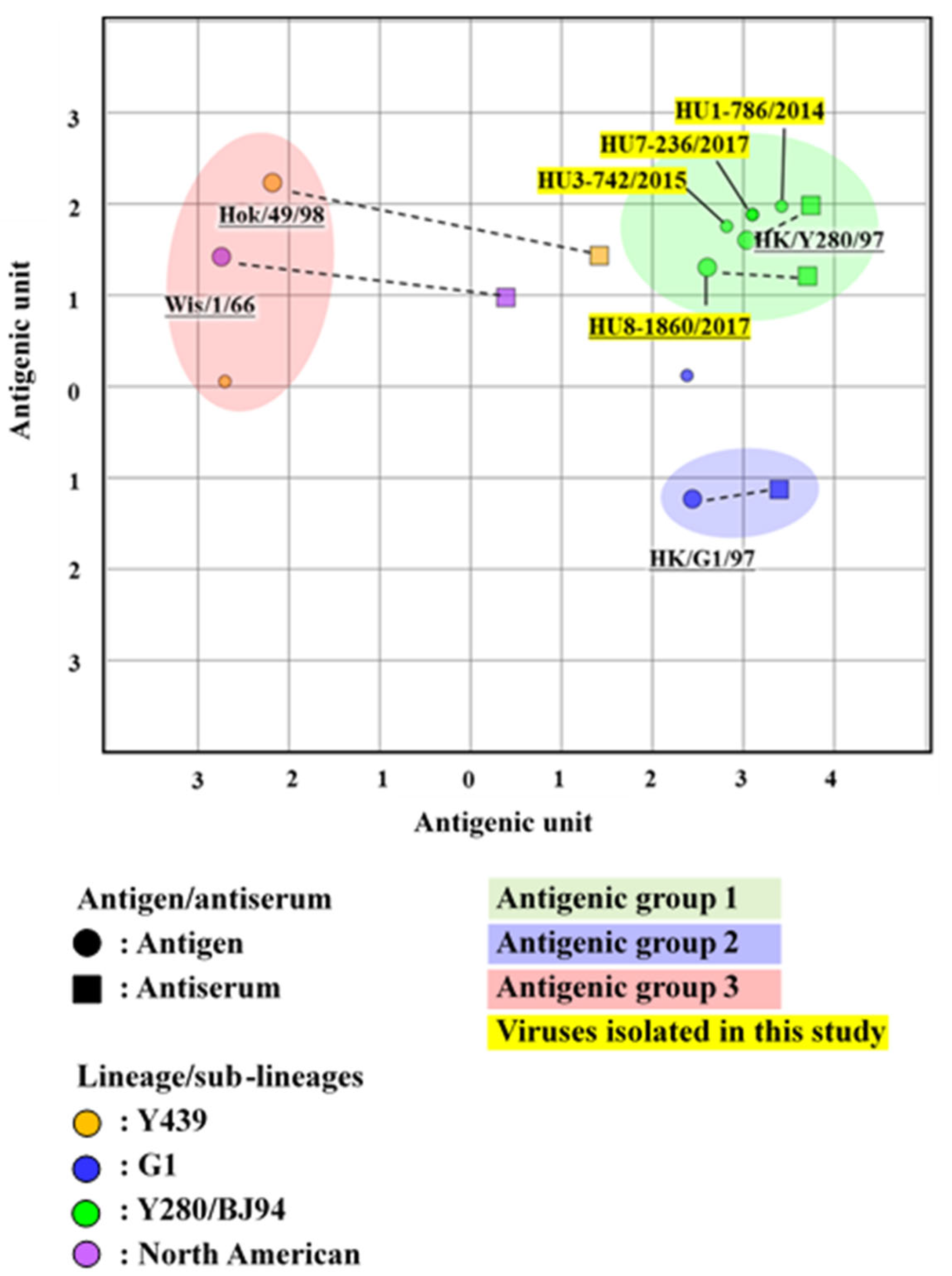
3.4. Antigenic Analysis of the H9 LPAIVs
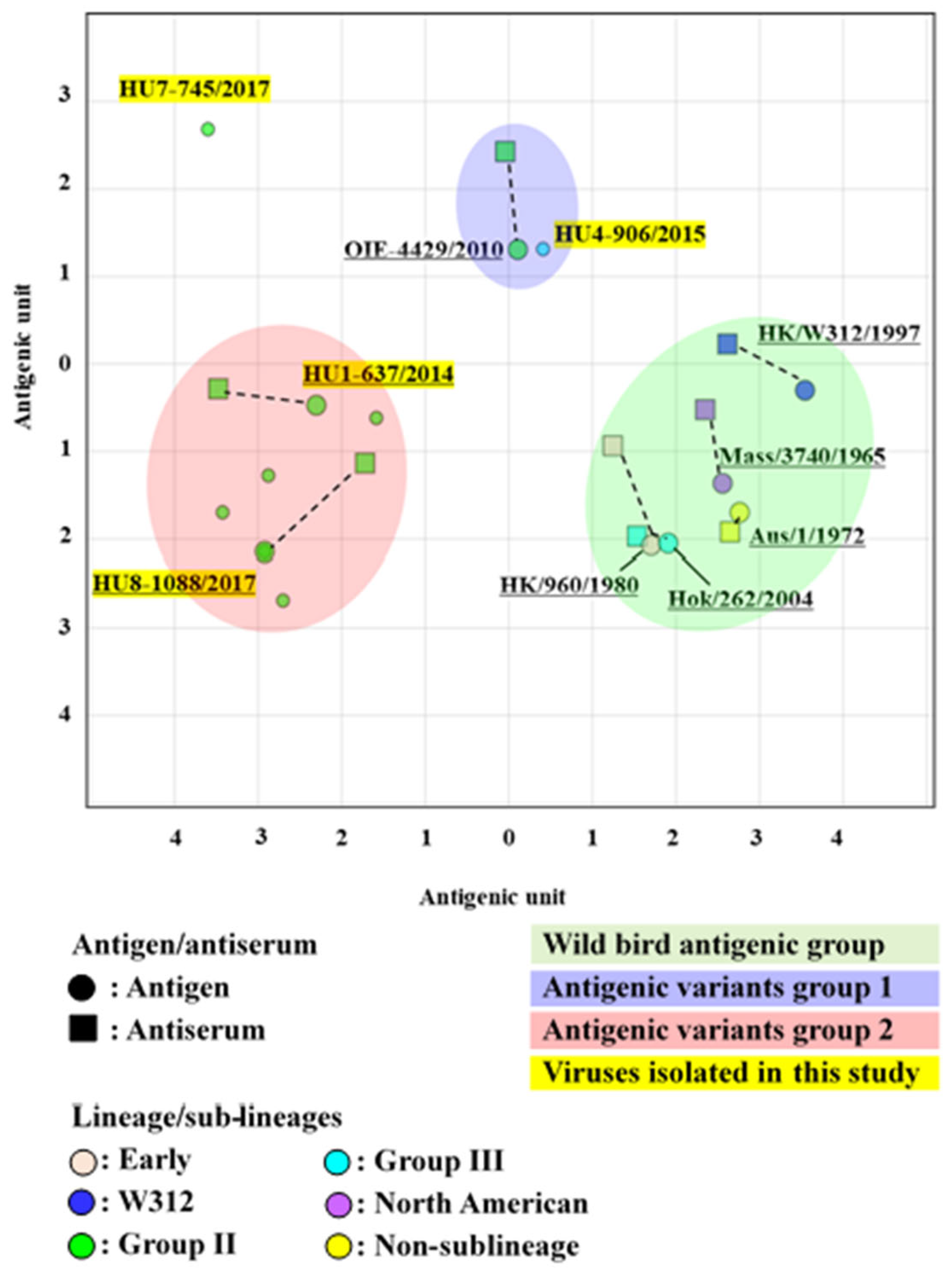
3.5. Antigenic Analysis of the H6 LPAIVs
3.6. Virulence-Associated Genes of APEC
3.7. Pathogenicity of H9N2 LPAIVs in the Coinfection with E. coli in Chickens
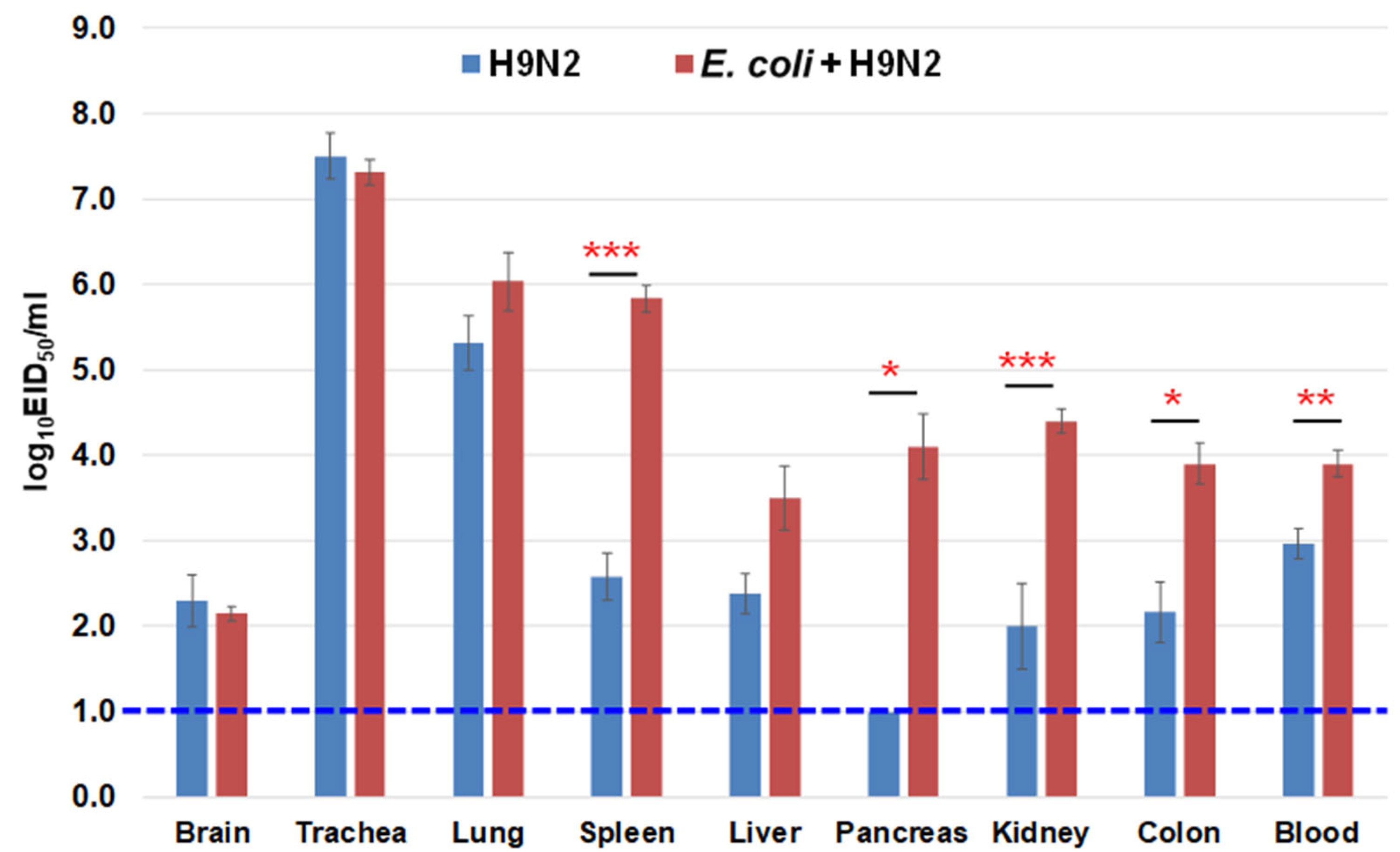
3.8. Virus Recovery from the Chicken Organs
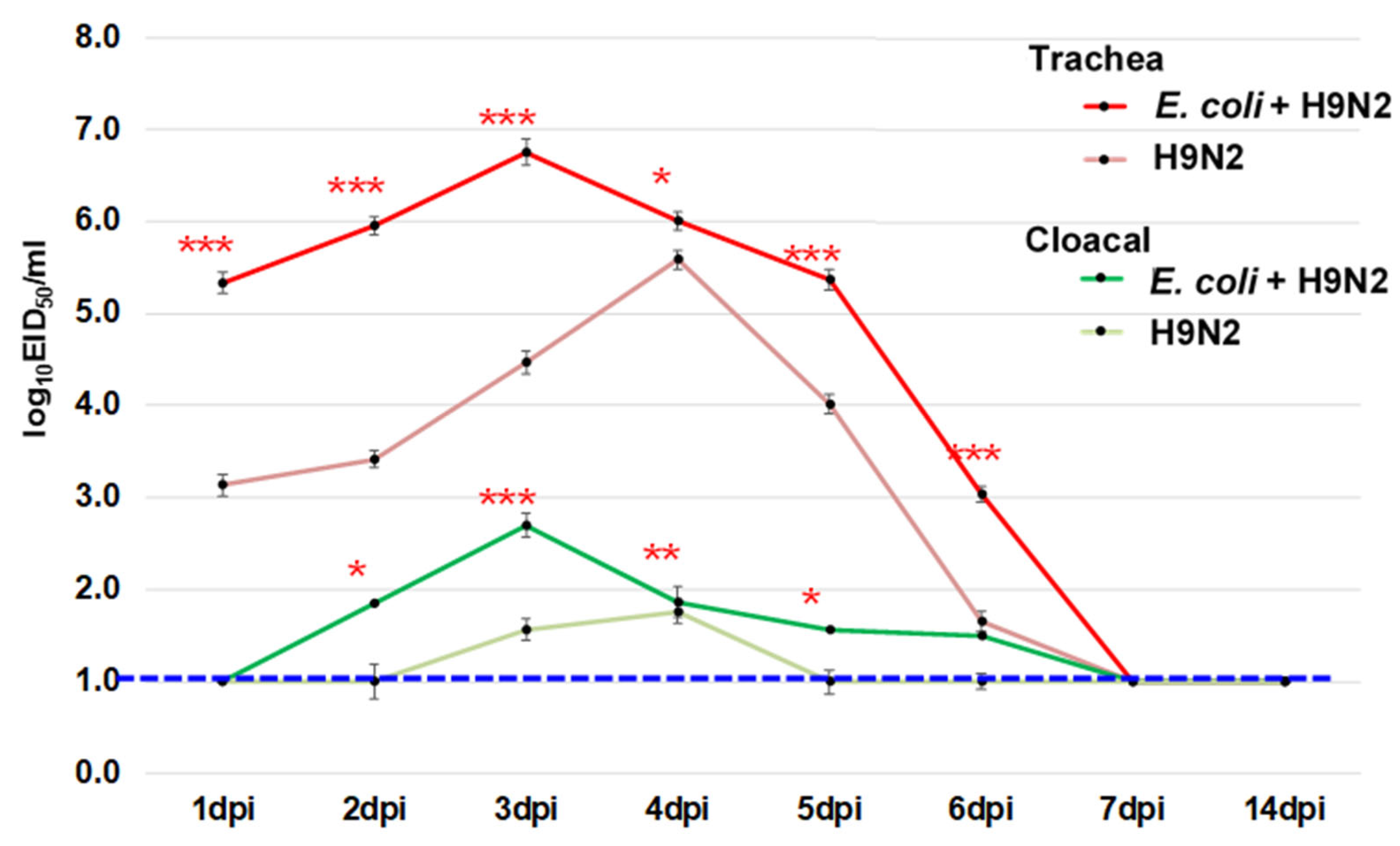
3.9. Virus Shedding during the Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howley, M.; Knipe, D. Orthomyxoriruses. In Fields Virology: Emerging Viruses, 7th ed.; Wolters Kluwer: Philadelphia, PA, USA, 2020; pp. 649–673. [Google Scholar]
- Yewdell, J.W.; Webster, R.G.; Gerhard, W.U. Antigenic variation in three distinct determinants of an influenza type A haemagglutinin molecule. Nature 1979, 279, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.K.; Seo, S.H.; Kim, J.A.; Webby, R.J.; Webster, R.G. Avian influenza viruses in Korean live poultry markets and their pathogenic potential. Virology 2005, 332, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinde, H.; Read, D.H.; Daft, B.M.; Hammarlund, M.; Moore, J.; Uzal, F.; Mukai, J.; Woolcock, P. The occurrence of avian influenza A subtype H6N2 in commercial layer flocks in southern California (2000–02): Clinicopathologic findings. Avian Dis. 2003, 47, 1214–1218. [Google Scholar] [CrossRef]
- Huang, K.; Bahl, J.; Fan, X.H.; Vijaykrishna, D.; Cheung, C.L.; Webby, R.J.; Webster, R.G.; Chen, H.; Smith, G.J.D.; Peiris, J.S.M.; et al. Establishment of an H6N2 Influenza virus lineage in domestic ducks in southern China. J. Virol. 2010, 84, 6978–6986. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.J.; Song, M.S.; Cruz, D.J.M.; Park, K.J.; Pascua, P.N.Q.; Lee, J.H.; Baek, Y.H.; Choi, D.H.; Choi, Y.K.; Kim, C.J. Active reassortment of H9 influenza viruses between wild birds and live-poultry markets in Korea. Arch. Virol. 2009, 155, 229–241. [Google Scholar] [CrossRef]
- Gowthaman, V.; Singh, S.D.; Dhama, K.; Ramakrishnan, M.A.; Malik, Y.P.S.; Murthy, T.R.G.K.; Chitra, R.; Munir, M. Co-infection of Newcastle disease virus genotype XIII with low pathogenic avian influenza exacerbates clinical outcome of Newcastle disease in vaccinated layer poultry flocks. Virusdisease 2019, 30, 441–452. [Google Scholar] [CrossRef]
- Pan, Q.; Liu, A.; Zhang, F.; Ling, Y.; Ou, C.; Hou, N.; He, C. Co-infection of broilers with Ornithobacterium rhinotracheale and H9N2 avian influenza virus. BMC Vet. Res. 2012, 8, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Kishida, N.; Sakoda, Y.; Eto, M.; Sunaga, Y.; Kida, H. Co-infection of Staphylococcus aureus or Haemophilus paragallinarum exacerbates H9N2 influenza A virus infection in chickens. Arch. Virol. 2004, 149, 2095–2104. [Google Scholar] [CrossRef]
- Quan, C.; Shi, W.; Yang, Y.; Yang, Y.; Liu, X.; Xu, W.; Li, H.; Li, J.; Wang, Q.; Tong, Z.; et al. New threats from H7N9 influenza virus: Spread and evolution of high- and low-pathogenicity variants with high genomic diversity in wave five. J. Virol. 2018, 92, e00301–e00318. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Shi, Y.; Wu, Y.; Liu, D.; Gao, G.F. Origin and molecular characterization of the human-infecting H6N1 influenza virus in Taiwan. Protein Cell 2013, 4, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Um, S.; Siegers, J.Y.; Sar, B.; Chin, S.; Patel, S.; Bunnary, S.; Hak, M.; Sor, S.; Sokhen, O.; Heng, S.; et al. Human infection with avian influenza A(H9N2) virus, Cambodia, February 2021. Emerg. Infect. Dis. 2021, 27, 2742–2745. [Google Scholar] [CrossRef]
- He, J.; Wu, Q.; Yu, J.-L.; He, L.; Sun, Y.; Shi, Y.-L.; Chen, Q.-Q.; Ge, Y.-L.; Zhang, Z.-H.; Li, W.-W.; et al. Sporadic occurrence of H9N2 avian influenza infections in human in Anhui province, eastern China: A notable problem. Microb. Pathog. 2019, 140, 103940. [Google Scholar] [CrossRef]
- Okamatsu, M.; Nishi, T.; Nomura, N.; Yamamoto, N.; Sakoda, Y.; Sakurai, K.; Chu, H.D.; Thanh, L.P.; Van Nguyen, L.; Van Hoang, N.; et al. The genetic and antigenic diversity of avian influenza viruses isolated from domestic ducks, muscovy ducks, and chickens in northern and southern Vietnam, 2010–2012. Virus Genes 2013, 47, 317–329. [Google Scholar] [CrossRef]
- Nomura, N.; Sakoda, Y.; Endo, M.; Yoshida, H.; Yamamoto, N.; Okamatsu, M.; Sakurai, K.; Van Hoang, N.; Van Nguyen, L.; Chu, H.D.; et al. Characterization of avian influenza viruses isolated from domestic ducks in Vietnam in 2009 and 2010. Arch. Virol. 2011, 157, 247–257. [Google Scholar] [CrossRef]
- Chu, D.-H.; Okamatsu, M.; Matsuno, K.; Hiono, T.; Ogasawara, K.; Nguyen, L.T.; Van Nguyen, L.; Nguyen, T.N.; Nguyen, T.T.; Van Pham, D.; et al. Genetic and antigenic characterization of H5, H6 and H9 avian influenza viruses circulating in live bird markets with intervention in the center part of Vietnam. Vet. Microbiol. 2016, 192, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Le, K.T.; Stevenson, M.A.; Isoda, N.; Nguyen, L.T.; Chu, D.; Nguyen, T.N.; Van Nguyen, L.; Tien, T.N.; Le, T.T.; Matsuno, K.; et al. A systematic approach to illuminate a new hot spot of avian influenza virus circulation in South Vietnam, 2016–2017. Transbound. Emerg. Dis. 2021, 69, e831–e844. [Google Scholar] [CrossRef]
- Le, K.T.; Isoda, N.; Nguyen, L.T.; Chu, D.-H.; Van Nguyen, L.; Phan, M.Q.; Nguyen, D.T.; Nguyen, T.N.; Tien, T.N.; Le, T.T.; et al. Risk profile of low pathogenicity avian influenza virus infections in farms in southern Vietnam. J. Vet. Med. Sci. 2022, 84, 860–868. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Mehat, J.W.; van Vliet, A.H.M.; La Ragione, R.M. The avian pathogenic Escherichia coli (APEC) pathotype is comprised of multiple distinct, independent genotypes. Avian Pathol. 2021, 50, 402–416. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2018; WOAH: Paris, France, 2018; Chapter 3.3.4. [Google Scholar]
- Kida, H.; Yanagawa, R. Isolation and characterization of influenza A virus from wild free-flying ducks in Hokkaido, Japan. Zentralbl. Bakteriol. Orig. A. 1979, 244, 135–143. [Google Scholar] [PubMed]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D.R. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakabe, S.; Sakoda, Y.; Haraguchi, Y.; Isoda, N.; Soda, K.; Takakuwa, H.; Saijo, K.; Sawata, A.; Kume, K.; Hagiwara, J.; et al. A vaccine prepared from a non-pathogenic H7N7 virus isolated from natural reservoir conferred protective immunity against the challenge with lethal dose of highly pathogenic avian influenza virus in chickens. Vaccine 2008, 26, 2127–2134. [Google Scholar] [CrossRef]
- Le, K.T.; Okamatsu, M.; Nguyen, L.T.; Matsuno, K.; Chu, D.-H.; Tien, T.N.; Le, T.T.; Kida, H.; Sakoda, Y. Genetic and antigenic characterization of the first H7N7 low pathogenic avian influenza viruses isolated in Vietnam. Infect. Genet. Evol. 2020, 78, 104117–104125. [Google Scholar] [CrossRef]
- Smith, D.J.; Lapedes, A.S.; De Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Mapping the antigenic and genetic evolution of influenza virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Meng, Q.; Dai, J.; Han, X.; Han, Y.; Ding, C.; Liu, H.; Yu, S. Development of an allele-specific PCR assay for simultaneous sero-typing of avian pathogenic Escherichia coli predominant O1, O2, O18 and O78 strains. PLoS ONE 2014, 9, e96904. [Google Scholar] [CrossRef] [Green Version]
- Jansen, T. Virulence-associated genes in avian pathogenic Escherichia coli (APEC) isolated from internal organs of poultry having died from colibacillosis. Int. J. Med. Microbiol. 2001, 291, 371–378. [Google Scholar] [CrossRef]
- Subedi, M.; Luitel, H.; Devkota, B.; Bhattarai, R.K.; Phuyal, S.; Panthi, P.; Shrestha, A.; Chaudhary, D.K. Antibiotic resistance pattern and virulence genes content in avian pathogenic Escherichia coli (APEC) from broiler chickens in Chitwan, Nepal. BMC Vet. Res. 2018, 14, 113–119. [Google Scholar] [CrossRef]
- Prioste, F.; Cunha, M.; Teixeira, R.; Zwargg, T.; Di Chiacchio, R.; Melville, P.; Benites, N.; Sinhorini, J.; Matushima, E.; Knöbl, T. Genetic similarity between APEC and Escherichia coli strains isolated from Guaruba Guarouba in a survey on healthy captive psittacine birds. Braz. J. Vet. Res. Anim. Sci. 2013, 50, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Horne, S.M.; Pfaff-McDonough, S.; Giddings, C.; Nolan, L. Cloning and sequencing of the iss gene from a virulent avian Escherichia coli. Avian Dis. 2000, 44, 179–184. [Google Scholar] [CrossRef]
- El-Sukhon, S.N.; Musa, A.; Al-Attar, M. Studies on the bacterial etiology of airsacculitis of broilers in northern and middle Jordan with special reference to Escherichia coli, Ornithobacterium rhinotracheale, and Bordetella avium. Avian Dis. 2002, 46, 605–612. [Google Scholar] [CrossRef]
- Stokholm, N.M.; Permin, A.; Bisgaard, M.; Christensen, J.P. Causes of Mortality in Commercial Organic Layers in Denmark. Avian Dis. 2010, 54, 1241–1250. [Google Scholar] [CrossRef]
- Nolan, L.K.; Barnes, H.J.; Vaillancourt, J.-P.; Abdul-Aziz, T.; Logue, C.M. Colibacillosis. In Diseases of Poultry; Wiley: New York, NY, USA, 2013; pp. 751–805. [Google Scholar] [CrossRef]
- R Foundation for statistical computing. R: A Language and Environment for Statistical Computing; R Foundation for statistical computing: Vienna, Austria, 2013. [Google Scholar]
- Nguyen, L.T.; Firestone, S.M.; Stevenson, M.A.; Young, N.D.; Sims, L.D.; Chu, D.H.; Nguyen, T.N.; Van Nguyen, L.; Le, T.T.; Van Nguyen, H.; et al. A systematic study towards evolutionary and epidemiological dynamics of currently predominant H5 highly pathogenic avian influenza viruses in Vietnam. Sci. Rep. 2019, 9, 7723–7736. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.-H.; Stevenson, M.A.; Nguyen, L.V.; Isoda, N.; Firestone, S.M.; Nguyen, T.N.; Matsuno, K.; Okamatsu, M.; Kida, H.; Sakoda, Y. A cross-sectional study to quantify the prevalence of avian influenza viruses in poultry at intervention and non-intervention live bird markets in central Vietnam, 2014. Transbound. Emerg. Dis. 2017, 64, 1991–1999. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Jin, X.; Hu, J.; Wu, Y.; Zhang, M.; Li, X.; Chen, J.; Xie, S.; Liu, J.; Qi, W.; et al. Genetic evolution characteristics of genotype G57 virus, a dominant genotype of H9N2 avian influenza virus. Front. Microbiol. 2021, 12, 633835. [Google Scholar] [CrossRef]
- Cui, J.; Cui, P.; Shi, J.; Fan, W.; Xing, X.; Gu, W.; Zhang, Y.; Zhang, Y.; Zeng, X.; Jiang, Y.; et al. Continued evolution of H6 avian influenza viruses isolated from farms in China between 2014 and 2018. Transbound. Emerg. Dis. 2021, 69, 2156–2172. [Google Scholar] [CrossRef]
- Koshikawa, N.; Hasegawa, S.; Nagashima, Y.; Mitsuhashi, K.; Tsubota, Y.; Miyata, S.; Miyagi, Y.; Yasumitsu, H.; Miyazaki, K. Expression of trypsin by epithelial cells of various tissues, leukocytes, and neurons in human and mouse. Am. J. Pathol. 1998, 153, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Swayne, D.E. Understanding the complex pathobiology of high pathogenicity avian influenza viruses in birds. Avian Dis. 2007, 51, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Tse, L.V.; Hamilton, A.M.; Friling, T.; Whittaker, G.R. A novel activation mechanism of avian influenza virus H9N2 by Furin. J. Virol. 2014, 88, 1673–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laporte, M.; Naesens, L. Airway proteases: An emerging drug target for influenza and other respiratory virus infections. Curr. Opin. Virol. 2017, 24, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, Y.; Chen, L.; Zhang, B.; Zhang, M.; Wang, J.; Jiang, Y.; Yang, C.; Jiang, T. Genetic characteristics and pathogenicity analysis in chickens and mice of three H9N2 avian influenza viruses. Viruses 2019, 11, 1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.-H.; Li, P.-Y.; Xu, M.-J.; Wang, C.-L.; Li, C.-H.; Gao, J.-P.; Wang, X.-J.; Xu, T.; Zhang, H.-L.; Tian, S.-F. Molecular characterization and pathogenesis of H9N2 avian influenza virus isolated from a racing pigeon. Vet. Microbiol. 2020, 246, 108747. [Google Scholar] [CrossRef]
- Kye, S.-J.; Park, M.-J.; Kim, N.-Y.; Lee, Y.-N.; Heo, G.-B.; Baek, Y.-K.; Shin, J.-I.; Lee, M.-H.; Lee, Y.-J. Pathogenicity of H9N2 low pathogenic avian influenza viruses of different lineages isolated from live bird markets tested in three animal models: SPF chickens, Korean native chickens, and ducks. Poult. Sci. 2021, 100, 101318. [Google Scholar] [CrossRef]
- El Sayes, M.; Kandeil, A.; Moatasim, Y.; El Taweel, A.; Rubrum, A.; Kutkat, O.; Kamel, M.N.; Badra, R.; Barakat, A.B.; McKenzie, P.P.; et al. Insights into genetic characteristics and virological features of endemic avian influenza A (H9N2) viruses in Egypt from 2017–2021. Viruses 2022, 14, 1484. [Google Scholar] [CrossRef]
- Baron, J.; Tarnow, C.; Mayoli-Nüssle, D.; Schilling, E.; Meyer, D.; Hammami, M.; Schwalm, F.; Steinmetzer, T.; Guan, Y.; Garten, W.; et al. Matriptase, HAT, and TMPRSS2 activate the hemagglutinin of H9N2 influenza A viruses. J. Virol. 2013, 87, 1811–1820. [Google Scholar] [CrossRef] [Green Version]
- Jaleel, S.; Younus, M.; Idrees, A.; Arshad, M.; Khan, A.U.; Ehtisham-Ul-Haque, S.; Zaheer, M.I.; Tanweer, M.; Towakal, F.; Munibullah, K.A.; et al. Pathological alterations in respiratory system during co-infection with low pathogenic avian influenza virus (H9N2) and Escherichia coli in broiler chickens. J. Vet. Res. 2017, 61, 253–258. [Google Scholar] [CrossRef]
- Costa-Hurtado, M.; Afonso, C.L.; Miller, P.J.; Spackman, E.; Kapczynski, D.R.; E Swayne, D.; Shepherd, E.; Smith, D.; Zsak, A.; Pantin-Jackwood, M. Virus interference between H7N2 low pathogenic avian influenza virus and lentogenic Newcastle disease virus in experimental co-infections in chickens and turkeys. Vet. Res. 2014, 45, 1. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, S.I.A.; Zyan, K.A.; Hamoud, M.M.; Khalifa, E.; Dardir, S.; Khalifa, R.; Kilany, W.H.; Elfeil, W.K. Effect of co-infection of low pathogenic avian influenza H9N2 virus and avian pathogenic E. coli on H9N2-vaccinated commercial broiler chickens. Front. Vet. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Mosleh, N.; Dadras, H.; Asasi, K.; Taebipour, M.J.; Tohidifar, S.S.; Farjanikish, G. Evaluation of the timing of the Escherichia coli co-infection on pathogenecity of H9N2 avian influenza virus in broiler chickens. Iran. J. Vet. Res. 2017, 18, 86–91. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, K.T.; Nguyen, L.T.; Huynh, L.T.; Chu, D.-H.; Nguyen, L.V.; Nguyen, T.N.; Tien, T.N.; Matsuno, K.; Okamatsu, M.; Hiono, T.; et al. Genetic, Antigenic, and Pathobiological Characterization of H9 and H6 Low Pathogenicity Avian Influenza Viruses Isolated in Vietnam from 2014 to 2018. Microorganisms 2023, 11, 244. https://doi.org/10.3390/microorganisms11020244
Le KT, Nguyen LT, Huynh LT, Chu D-H, Nguyen LV, Nguyen TN, Tien TN, Matsuno K, Okamatsu M, Hiono T, et al. Genetic, Antigenic, and Pathobiological Characterization of H9 and H6 Low Pathogenicity Avian Influenza Viruses Isolated in Vietnam from 2014 to 2018. Microorganisms. 2023; 11(2):244. https://doi.org/10.3390/microorganisms11020244
Chicago/Turabian StyleLe, Kien Trung, Lam Thanh Nguyen, Loc Tan Huynh, Duc-Huy Chu, Long Van Nguyen, Tien Ngoc Nguyen, Tien Ngoc Tien, Keita Matsuno, Masatoshi Okamatsu, Takahiro Hiono, and et al. 2023. "Genetic, Antigenic, and Pathobiological Characterization of H9 and H6 Low Pathogenicity Avian Influenza Viruses Isolated in Vietnam from 2014 to 2018" Microorganisms 11, no. 2: 244. https://doi.org/10.3390/microorganisms11020244