
VT-1161—A Tetrazole for Management of Mono- and Dual-Species Biofilms
 , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganism Strains and Culture Conditions
Cell Cultures
2.3. Antimicrobial Assay (MIC)
2.4. Biofilm Formation and Biofilm Mass Quantification
2.5. Activity of VT-1161 against Mono- and Dual-Species Biofilms
2.6. Cell Cytotoxicity Assay
2.7. Adhesion to PNT1A Cells
2.8. Toxicity Assays in the Nematode Model
2.9. C. elegans Infection Assay
2.10. Antivirulence Assays in the Nematode Model
2.11. Statistical Analysis
3. Results
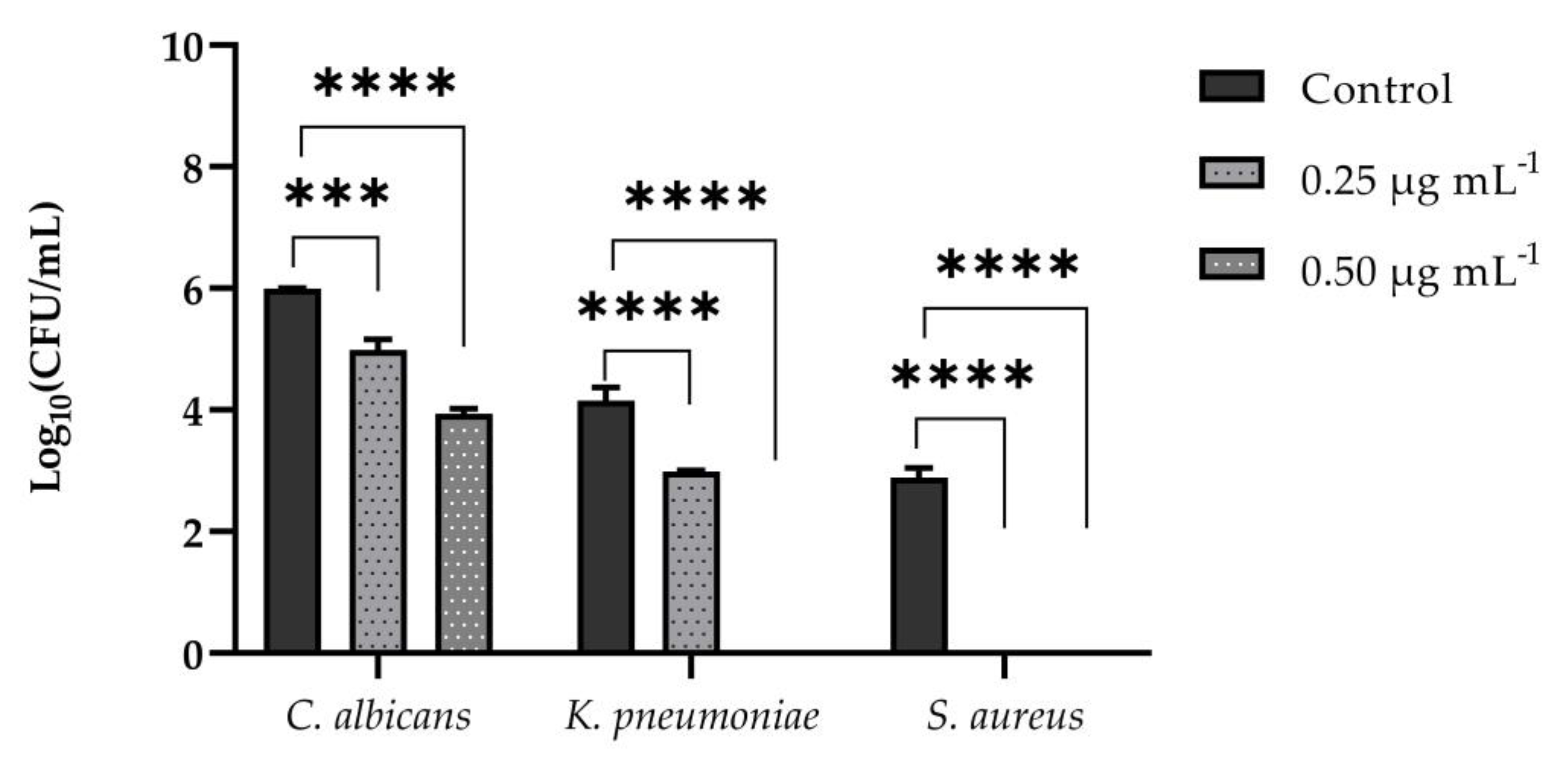
3.1. Determination of Minimum Inhibitory Concentration (MIC)
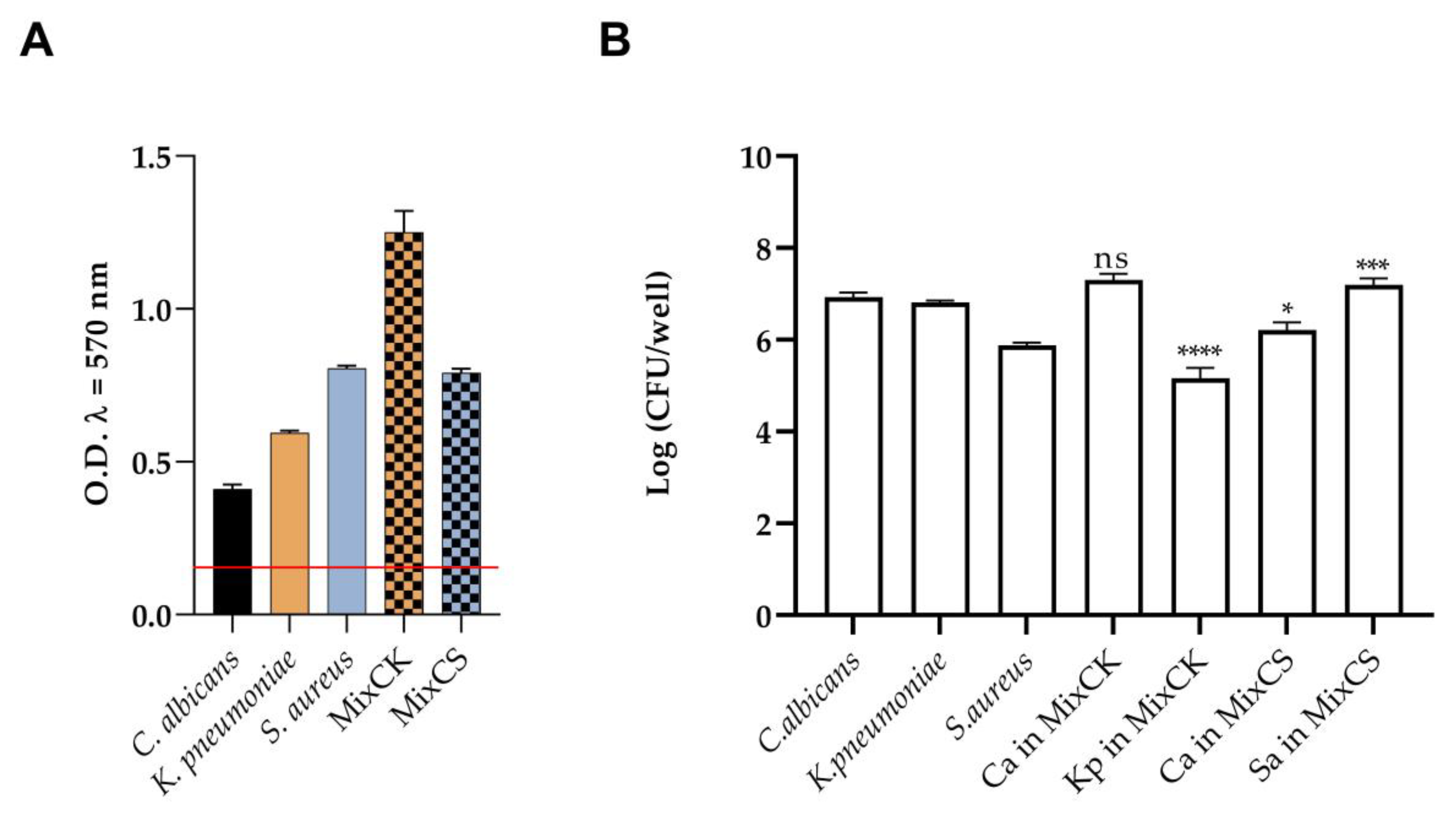
3.2. Quantitative Biofilm Production
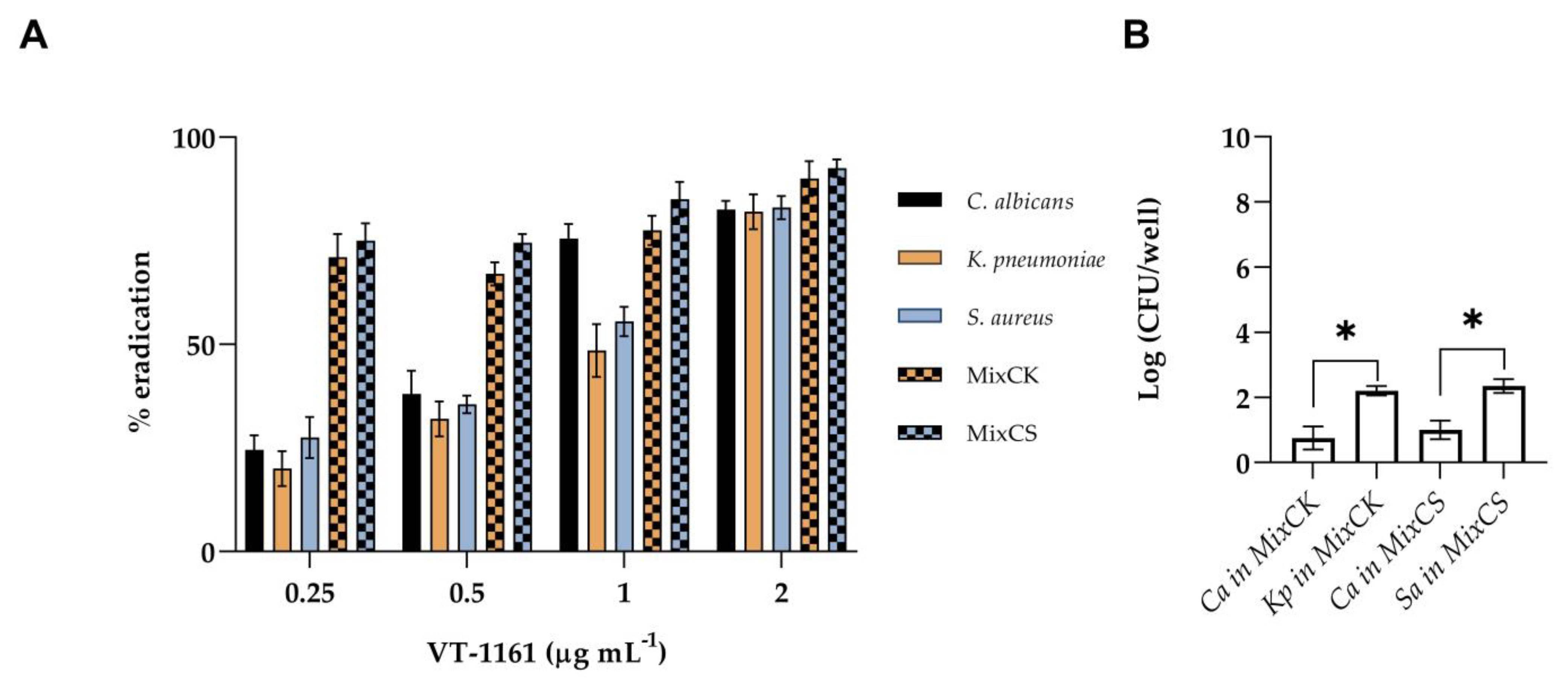
3.3. Inhibition and Eradication of Monomicrobial and Polymicrobial Biofilms
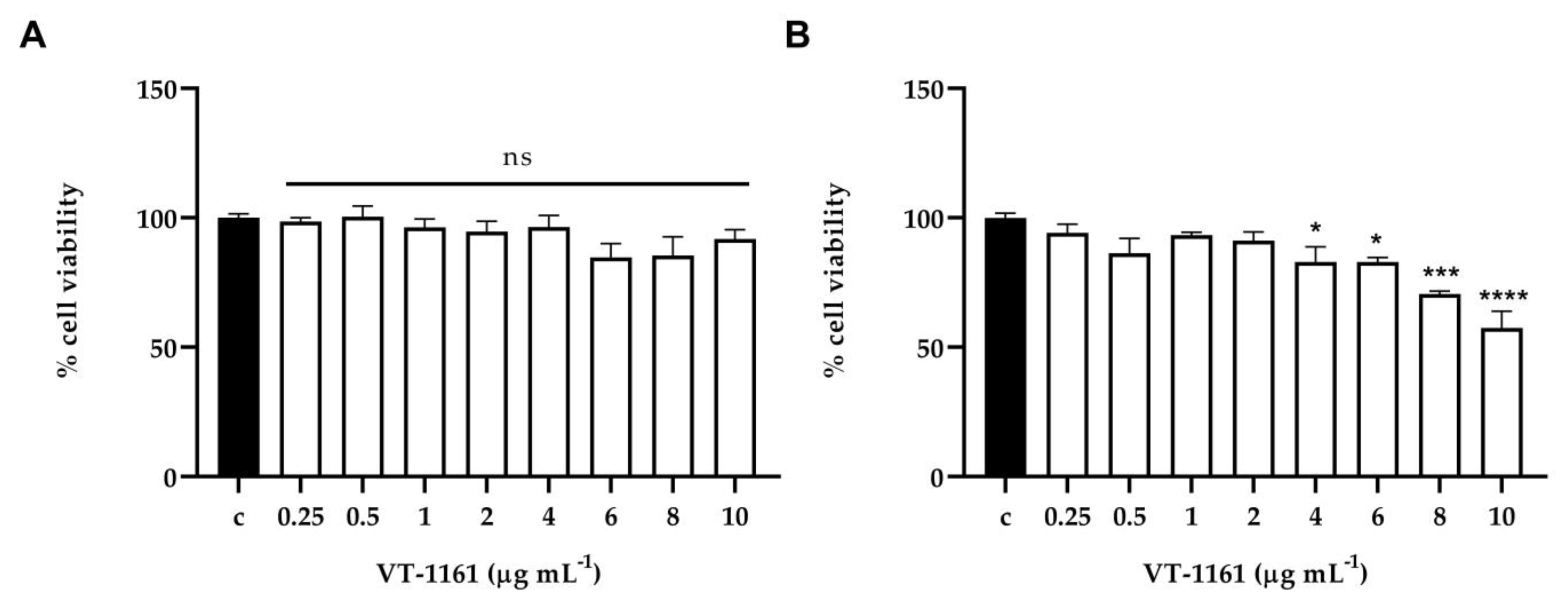
3.4. Toxicity Assay
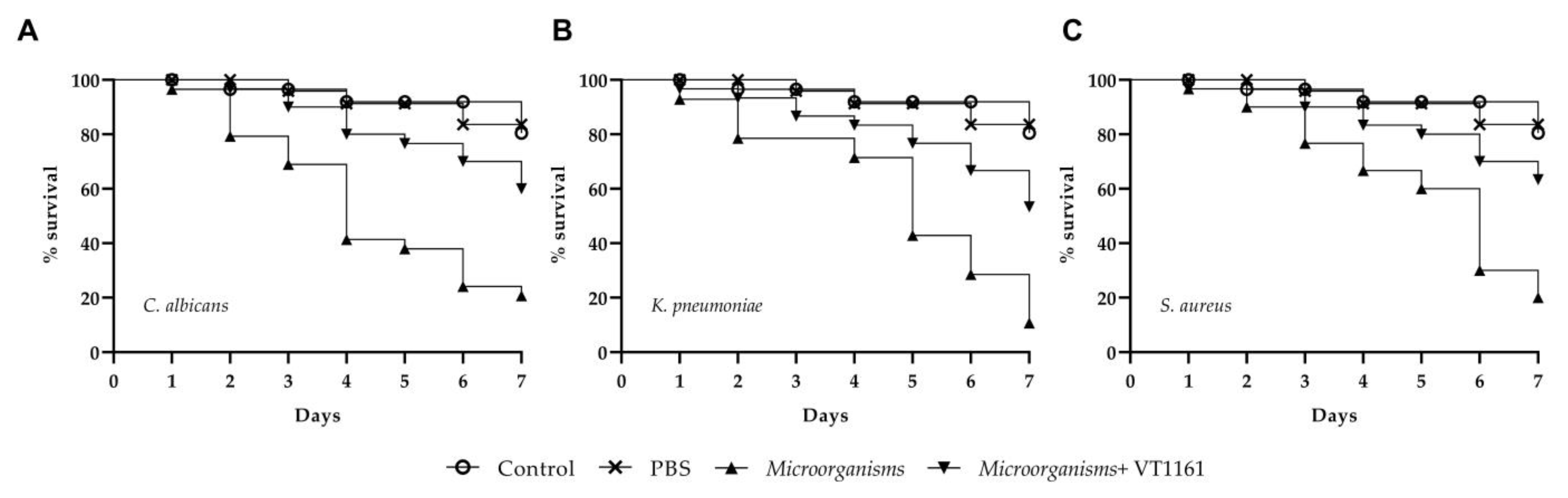
3.5. In Vitro and In Vivo Infection Assay
3.5.1. Adhesion Assay on PNT1A Cells
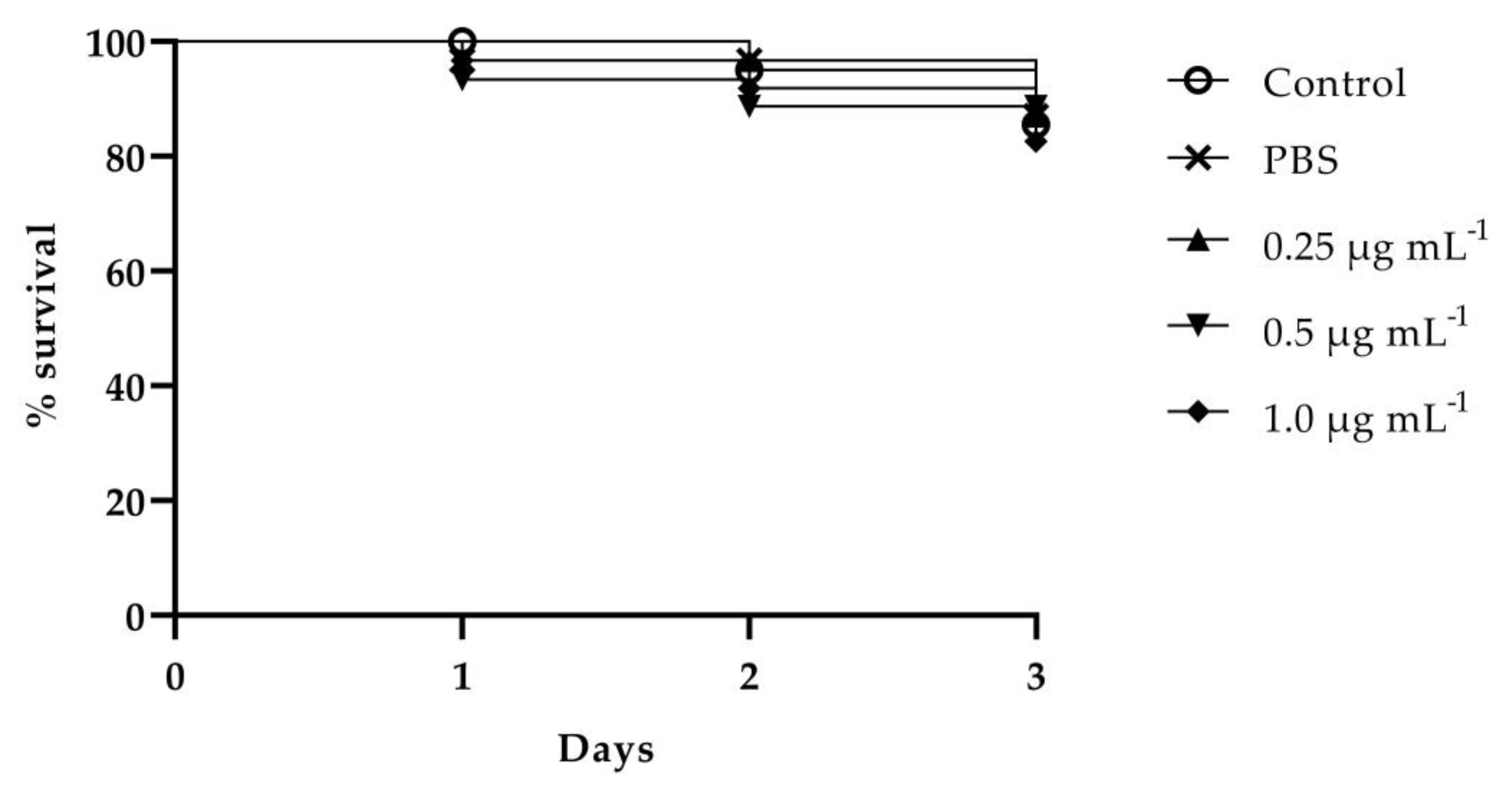
3.5.2. C. elegans Infection Assay and Effect of VT-1161 on C. elegans Survival
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kovács, R.; Majoros, L. Fungal Quorum-Sensing Molecules: A Review of Their Antifungal Effect against Candida Biofilms. J. Fungi 2020, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.; Pandian, S.K. Piperine Impedes Biofilm Formation and Hyphal Morphogenesis of Candida albicans. Front. Microbiol. 2020, 11, 756. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hu, X.; Wu, Y.; Zhang, W.; Chen, X.; You, X.; Hu, L. Synthesis and structure-activity relationship of novel bisindole amidines active against MDR Gram-positive and Gram-negative bacteria. Eur. J. Med. Chem. 2018, 150, 771–782. [Google Scholar] [CrossRef]
- Förster, T.M.; Mogavero, S.; Dräger, A.; Graf, K.; Polke, M.; Jacobsen, I.D.; Hube, B. Enemies and brothers in arms: Candida albicans and gram-positive bacteria. Cell. Microbiol. 2016, 18, 1709–1715. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef]
- Galdiero, E.; Ricciardelli, A.; D’Angelo, C.; de Alteriis, E.; Maione, A.; Albarano, L.; Casillo, A.; Corsaro, M.; Tutino, M.; Parrilli, E. Pentadecanoic acid against Candida albicans-Klebsiella pneumoniae biofilm: Towards the development of an anti-biofilm coating to prevent polymicrobial infections. Res. Microbiol. 2021, 172, 103880. [Google Scholar] [CrossRef]
- Galdiero, E.; Salvatore, M.M.; Maione, A.; de Alteriis, E.; Andolfi, A.; Salvatore, F.; Guida, M. GC-MS-based metabolomics study of single-and dual-species biofilms of Candida albicans and Klebsiella pneumoniae. Int. J. Mol. Sci. 2021, 22, 3496. [Google Scholar] [CrossRef]
- Collins, L.M.; Moore, R.; Sobel, J.D. Prognosis and long-term outcome of women with idiopathic recurrent vulvovaginal candidiasis caused by Candida albicans. J. Low. Genit. Tract Dis. 2020, 24, 48–52. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef]
- Hoekstra, W.J.; Garvey, E.P.; Moore, W.R.; Rafferty, S.W.; Yates, C.M.; Schotzinger, R.J. Design and optimization of highly-selective fungal CYP51 inhibitors. Bioorganic Med. Chem. Lett. 2014, 24, 3455–3458. [Google Scholar] [CrossRef]
- Warrilow, A.G.; Hull, C.M.; Parker, J.E.; Garvey, E.P.; Hoekstra, W.J.; Moore, W.R.; Schotzinger, R.J.; Kelly, D.E.; Kelly, S.L. The clinical candidate VT-1161 is a highly potent inhibitor of Candida albicans CYP51 but fails to bind the human enzyme. Antimicrob. Agents Chemother. 2014, 58, 7121–7127. [Google Scholar] [CrossRef] [Green Version]
- FDA Approves Mycovia Pharmaceuticals’ VIVJOA™ (oteseconazole), the First and Only FDA-Approved Medication for Recurrent Vulvovaginal Candidiasis (Chronic Yeast Infection). Available online: https://www.fda.gov/drugs/new-drugs-fda-cders-new-molecular-entities-and-new-therapeutic-biological-products/novel-drug-approvals-2022 (accessed on 28 November 2022).
- Clinical & Laboratory Standards Institute. Reference method for broth dilution antifungal susceptibility testing of yeasts. In Interactions of Yeasts, Moulds, and Antifungal Agents; Springer: Berlin/Heidelberg, Germany, 2012; pp. 65–74. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard, Pennsylvania, 19087–11898; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Falanga, A.; Maione, A.; La Pietra, A.; de Alteriis, E.; Vitale, S.; Bellavita, R.; Carotenuto, R.; Turrà, D.; Galdiero, S.; Galdiero, E.; et al. Competitiveness during Dual-Species Biofilm Formation of Fusarium oxysporum and Candida albicans and a Novel Treatment Strategy. Pharmaceutics 2022, 14, 1167. [Google Scholar] [CrossRef]
- Maione, A.; La Pietra, A.; de Alteriis, E.; Mileo, A.; De Falco, M.; Guida, M.; Galdiero, E. Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida auris and Klebsiella pneumoniae. Microorganisms 2022, 10, 1773. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Forte, M.; Di Lorenzo, M.; Iachetta, G.; Mita, D.G.; Laforgia, V.; De Falco, M. Nonylphenol acts on prostate adenocarcinoma cells via estrogen molecular pathways. Ecotoxicol. Environ. Saf. 2019, 180, 412–419. [Google Scholar] [CrossRef]
- Escobar, I.E.; Possamai Rossatto, F.C.; Kim, S.M.; Kang, M.H.; Kim, W.; Mylonakis, E. Repurposing Kinase Inhibitor Bay 11-7085 to Combat Staphylococcus aureus and Candida albicans Biofilms. Front. Pharm. 2021, 12, 675300. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef] [Green Version]
- Němeček, J.; Sychra, P.; Macháček, M.; Benková, M.; Karabanovich, G.; Konečná, K.; Kavková, V.; Stolaříková, J.; Hrabálek, A.; Vávrová, K.; et al. Structure-activity relationship studies on 3,5-dinitrophenyl tetrazoles as antitubercular agents. Eur. J. Med. Chem. 2017, 130, 419–432. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Gan, L.L.; Wang, H.; Zhou, C.H. New Progress in Azole Compounds as Antimicrobial Agents. Mini Rev. Med. Chem. 2017, 17, 122–166. [Google Scholar] [CrossRef] [PubMed]
- Farrokhi, Y.; Al-Shibli, B.; Al-Hameedawi, D.F.; Neshati, Z.; Makhdoumi, A. Escherichia coli enhances the virulence factors of Candida albicans, the cause of vulvovaginal candidiasis, in a dual bacterial/fungal biofilm. Res. Microbiol. 2021, 172, 103849. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Lewis, R.E.; Kontoyiannis, D.P. Investigational Antifungal Agents for Invasive Mycoses: A Clinical Perspective. Clin. Infect. Dis. 2022, 75, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Vallières, C.; Singh, N.; Alexander, C.; Avery, S.V. Repurposing Nonantifungal Approved Drugs for Synergistic Targeting of Fungal Pathogens. ACS Infect. Dis. 2020, 6, 2950–2958. [Google Scholar] [CrossRef]
- Gao, F.; Johnson, D.L.; Ekins, S.; Janiszewski, J.; Kelly, K.G.; Meyer, R.D.; West, M. Optimizing higher throughput methods to assess drug-drug interactions for CYP1A2, CYP2C9, CYP2C19, CYP2D6, rCYP2D6, and CYP3A4 in vitro using a single point IC50. J. Biomol. Screen. 2002, 7, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Brand, S.R.; Degenhardt, T.P.; Person, K.; Sobel, J.D.; Nyirjesy, P.; Schotzinger, R.J.; Tavakkol, A. A phase 2, randomized, double-blind, placebo-controlled, dose-ranging study to evaluate the efficacy and safety of orally administered VT-1161 in the treatment of recurrent vulvovaginal candidiasis. Am. J. Obs. Gynecol 2018, 218, 624.e621–624.e629. [Google Scholar] [CrossRef]
- Nishimoto, A.T.; Wiederhold, N.P.; Flowers, S.A.; Zhang, Q.; Kelly, S.L.; Morschhäuser, J.; Yates, C.M.; Hoekstra, W.J.; Schotzinger, R.J.; Garvey, E.P.; et al. In Vitro Activities of the Novel Investigational Tetrazoles VT-1161 and VT-1598 Compared to the Triazole Antifungals against Azole-Resistant Strains and Clinical Isolates of Candida albicans. Antimicrob. Agents Chemother. 2019, 63, e00341-19. [Google Scholar] [CrossRef] [Green Version]
- Schell, W.A.; Jones, A.M.; Garvey, E.P.; Hoekstra, W.J.; Schotzinger, R.J.; Alexander, B.D. Fungal CYP51 Inhibitors VT-1161 and VT-1129 Exhibit Strong In Vitro Activity against Candida glabrata and C. krusei Isolates Clinically Resistant to Azole and Echinocandin Antifungal Compounds. Antimicrob. Agents Chemother. 2017, 61, e01817-16. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Xiao, J.; Huang, G. Current scenario of tetrazole hybrids for antibacterial activity. Eur. J. Med. Chem. 2019, 184, 111744. [Google Scholar] [CrossRef]
- Nobre, L.S.; Todorovic, S.; Tavares, A.F.; Oldfield, E.; Hildebrandt, P.; Teixeira, M.; Saraiva, L.M. Binding of azole antibiotics to Staphylococcus aureus flavohemoglobin increases intracellular oxidative stress. J. Bacteriol. 2010, 192, 1527–1533. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC (µgmL−1) | ||||
|---|---|---|---|---|
| VT-1161 | FLC | MEM | VAN | |
| C. albicans | 2.0 | 16.0 | - | - |
| K. pneumoniae | 0.5 | - | 1.0 | - |
| S. aureus | 0.5 | - | - | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maione, A.; Mileo, A.; Pugliese, S.; Siciliano, A.; Cirillo, L.; Carraturo, F.; de Alteriis, E.; De Falco, M.; Guida, M.; Galdiero, E. VT-1161—A Tetrazole for Management of Mono- and Dual-Species Biofilms. Microorganisms 2023, 11, 237. https://doi.org/10.3390/microorganisms11020237
Maione A, Mileo A, Pugliese S, Siciliano A, Cirillo L, Carraturo F, de Alteriis E, De Falco M, Guida M, Galdiero E. VT-1161—A Tetrazole for Management of Mono- and Dual-Species Biofilms. Microorganisms. 2023; 11(2):237. https://doi.org/10.3390/microorganisms11020237
Chicago/Turabian StyleMaione, Angela, Aldo Mileo, Stefano Pugliese, Antonietta Siciliano, Luigi Cirillo, Federica Carraturo, Elisabetta de Alteriis, Maria De Falco, Marco Guida, and Emilia Galdiero. 2023. "VT-1161—A Tetrazole for Management of Mono- and Dual-Species Biofilms" Microorganisms 11, no. 2: 237. https://doi.org/10.3390/microorganisms11020237