
Exploring Acetogenesis in Firmicutes: From Phylogenetic Analysis to Solid Medium Cultivation with Solid-Phase Electrochemical Isolation Equipments

Abstract
:1. Introduction
2. Materials and Methods
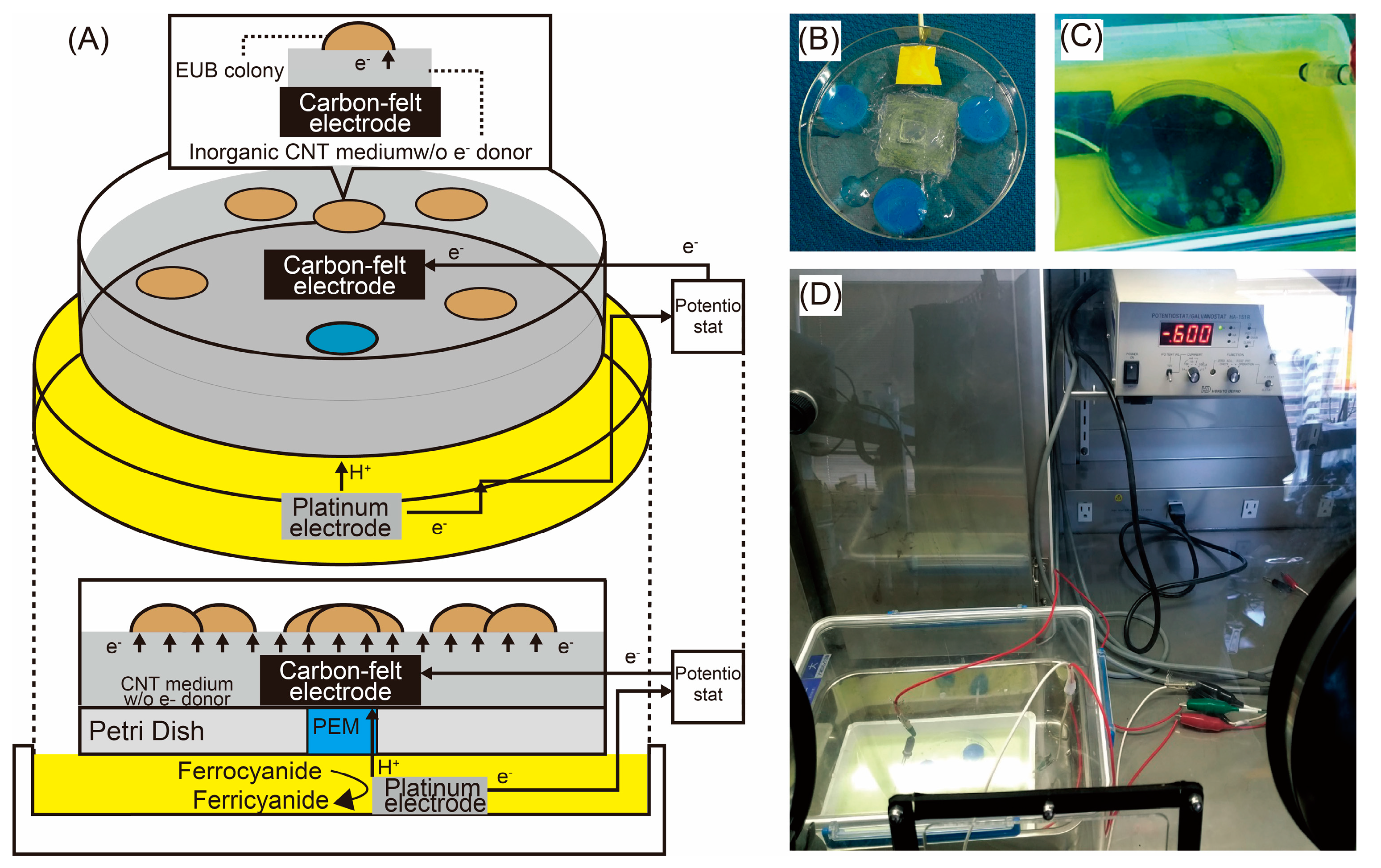
2.1. SPECIEs: A Novel Approach for Electrotrophic Acetogen Isolation
2.2. Genomic DNA Extraction and PCR Conditions
2.3. Sequencing Library Preparation, Sequencing, and Bioinformatics Analysis
2.4. Phylogenetic Tree of Electrotrophic Acetogens and Their Characterization
2.5. Selection and Culture of Electrotrophic Acetogen Candidates
2.6. Electrochemical Culture of Selected Bacteria
2.7. Lowry Method for Protein Quantitation
3. Results and Discussion
3.1. Electrochemical Cultivation of Potential Acetogens on Solid Medium
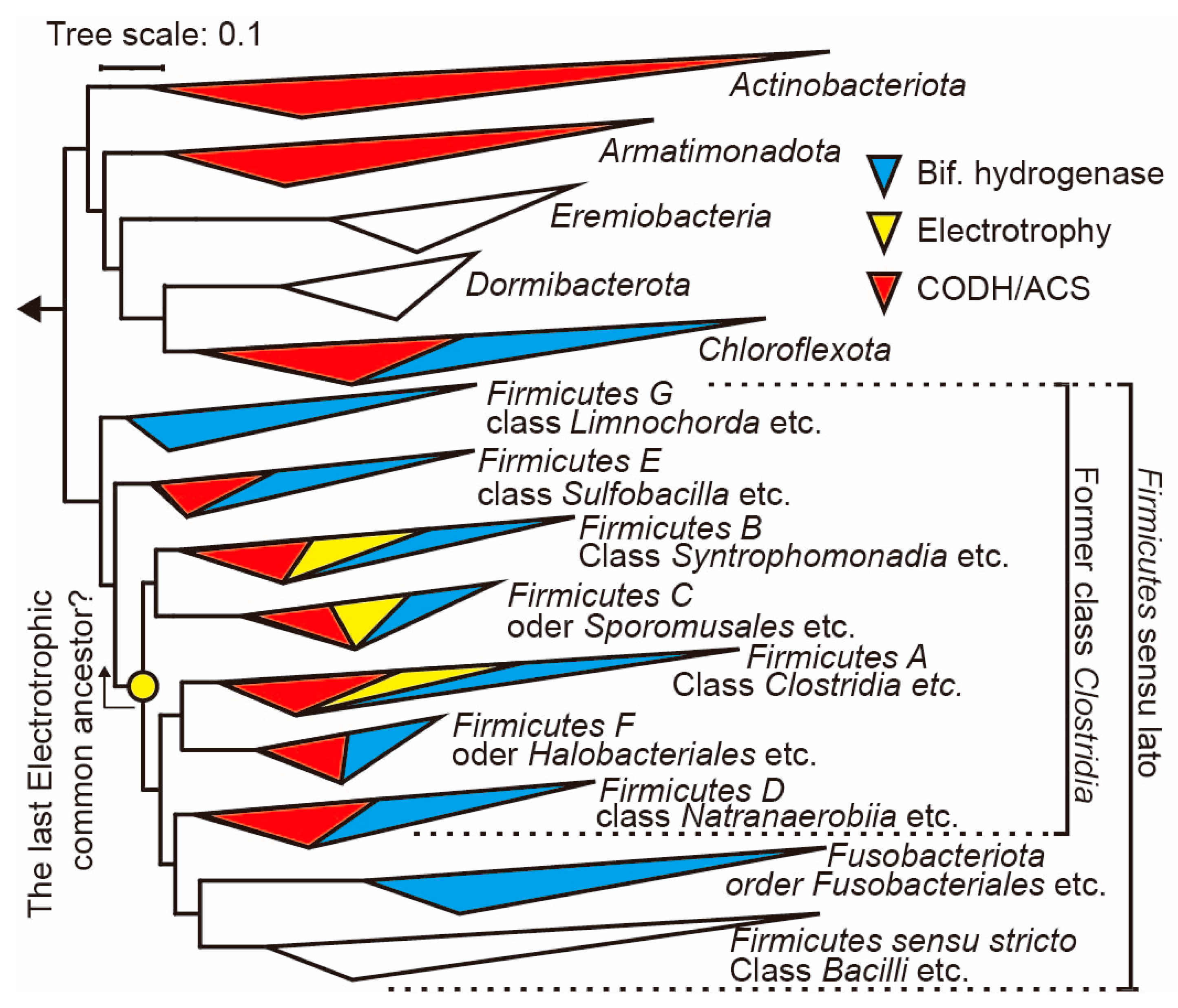
3.2. Phylogenetic Analysis of Acetogens: Elucidating Latent Electrotrophic Traits within Firmicutes Sensu Lato
3.3. Evaluating Latent Electrotrophic Potential in Selected Firmicutes Strains: Uncovering New MES Candidates
3.4. Development Potential and Limitations of SPECIEs and Future Challenges

{kind=link}
{kind=link}
{kind=link}
| Activity | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Phylum | Class | CODH/ACS Genes | pta Gene | Electron Utilizing | Bifurcating Hydrogenase Gene | Species Name/MAG Source | Assembly/Protein Accession | Reference of E-Utilizing Activity | |
| Actinobacteriota | Coriobacteriia | + | + | N.R. | - | Rumen | GCA_900314665 | ||
| Aromatimonadota | Abditibacteria | + | + | N.R. | - | groundwater filtered through a 3.0 um filter | GCA_001872605 | ||
| Eremiobacteria | - | - | - | N.R. | - | - | - | ||
| Dormibacterota | - | - | - | N.R. | - | - | - | ||
| Chloroflexota | Dehaloccocoidia | + | + | N.R. | + | wastewater | GCA_002436065 | ||
| Firmicutes sensu lato | Firmicutes G | UBA4882 | - | - | N.R. | + | Hydrogenispora ethanolica | GCF_004340685 | |
| Firmicutes E | DTU015 | + | + | N.R. | + | Hydrothermal vent biofilm | GCA_002291985 | ||
| Firmicutes B | Moorellia | + | + | + | + | Moorella thermoacetica | GCF_001267405/WP_011393219 | Nevin et al., 2011 [13] | |
| Firmicutes C | Negativicutes | + | + | + | + | Sporomusa ovata | GCF_000445445/WP_021167212 | Nevin et al., 2011 [13] | |
| Firmicutes A | Clostridia | + | + | + | + | Clostridium ljungdahli | GCF_000143685/WP_013238135 | Im et al., 2022 [51] | |
| Firmicutes F | Halanaerobiia | + | + | N.R. | + | Acetohalanaerobium arabaticum | GCF_000144695/WP_013277223 | ||
| Firmicutes D | Dethiobacteria | + | + | N.R. | + | Hydrothermal vent biofilm | GCA_002292015 | ||
| Fusobacteriota | Fusobacteriia | - | - | N.R. | + | Fusobacterium varium | WP_005950651 | ||
| Firmicutes sensu stricto | - | - | - | N.R. | - | - | - | ||
| Strain Name | Sampling Conditions | Protein Concentration (mg-BSA/mL) [A] | Total Volume of Sampling Locations (mL) [B] | Total Protein Weight at Sampling Location (mg) [A × B] | Net Protein Increase (mg) | Potential for Growth under Electrode-Oxidation Conditions |
|---|---|---|---|---|---|---|
| Syntriphomonas erecta JCM 13344 | Planktonic cells post-cultivation | 0.272 ± 0.079 | 200 | 68.733 | −14.400 | Negative |
| Planktonic cells immediately after cell addition | 0.344 ± 0.065 | 200 | 54.333 | |||
| Electrode post-cultivation | 0.278 ± 0.055 | 10 | 2.780 | 0.66 | Negative | |
| Protein weight of the electrode immediately after cell addition (calculated value) | 0.344 × 10 = 3.44 | |||||
| Halanaerobacter jenidensis JCM 16696 | Planktonic cells post-cultivation | 0.692 ± 0.091 | 200 | 138.400 | 47.067 | Positive |
| Planktonic cells immediately after cell addition | 0.457 ± 0.129 | 200 | 91.333 | |||
| Electrode post-cultivation | 1.227 ± 0.157 | 10 | 12.270 | 7.700 | Positive | |
| Protein weight of the electrode immediately after cell addition (calculated value) | 0.457 × 10 = 4.57 | |||||
| Desulfotomaculum defulvii JCM 14036 | Planktonic cells post-cultivation | 0.395 ± 0.044 | 200 | 84.467 | −5.533 | Negative |
| Planktonic cells immediately after cell addition | 0.422 ± 0.052 | 200 | 78.933 | |||
| Electrode post-cultivation | 0.203 ± 0.086 | 10 | 2.027 | −2.193 | Negative | |
| Protein weight of the electrode immediately after cell addition (calculated value) | 0.422 × 10 = 4.22 |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Twidell, J. Renewable Energy Resources; Routledge: London, UK, 2021. [Google Scholar]
- Olah, G.A.; Prakash, G.S.; Goeppert, A. Anthropogenic chemical carbon cycle for a sustainable future. J. Am. Chem. Soc. 2011, 133, 12881–12898. [Google Scholar] [CrossRef]
- Zhang, Z.; Pan, S.-Y.; Li, H.; Cai, J.; Olabi, A.G.; Anthony, E.J.; Manovic, V. Recent advances in carbon dioxide utilization. Renew. Sustain. Energy Rev. 2020, 125, 109799. [Google Scholar] [CrossRef]
- Pavan, M.; Reinmets, K.; Garg, S.; Mueller, A.P.; Marcellin, E.; Köpke, M.; Valgepea, K. Advances in systems metabolic engineering of autotrophic carbon oxide-fixing biocatalysts towards a circular economy. Metab. Eng. 2022, 71, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Dessì, P.; Sánchez, C.; Mills, S.; Cocco, F.G.; Isipato, M.; Ijaz, U.Z.; Collins, G.; Lens, P.N. Carboxylic acids production and electrosynthetic microbial community evolution under different CO2 feeding regimens. Bioelectrochemistry 2021, 137, 107686. [Google Scholar] [CrossRef] [PubMed]
- Modestra, J.A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Prospects and trends in bioelectrochemical systems: Transitioning from CO2 towards a low-carbon circular bioeconomy. Bioresour. Technol. 2022, 364, 128040. [Google Scholar] [CrossRef] [PubMed]
- Borrel, G.; Adam, P.S.; Gribaldo, S. Methanogenesis and the Wood–Ljungdahl pathway: An ancient, versatile, and fragile association. Genome Biol. Evol. 2016, 8, 1706–1711. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Electrotrophy: Other microbial species, iron, and electrodes as electron donors for microbial respirations. Bioresour. Technol. 2022, 345, 126553. [Google Scholar] [CrossRef] [PubMed]
- Obi, C.N.; Asogwa, G.C. Electromicrobiology: An Emerging Reality—A Review. Open Access Libr. J. 2015, 2, 1–10. [Google Scholar] [CrossRef]
- Wang, H.; Ren, Z.J. A comprehensive review of microbial electrochemical systems as a platform technology. Biotechnol. Adv. 2013, 31, 1796–1807. [Google Scholar] [CrossRef]
- Tsui, T.-H.; Zhang, L.; Zhang, J.; Dai, Y.; Tong, Y.W. Engineering interface between bioenergy recovery and biogas desulfurization: Sustainability interplays of biochar application. Renew. Sustain. Energy Rev. 2022, 157, 112053. [Google Scholar] [CrossRef]
- Kim, M.; Li, S.; Song, Y.E.; Lee, D.-Y.; Kim, J.R. Electrode-attached cell-driven biogas upgrading of anaerobic digestion effluent CO2 to CH4 using a microbial electrosynthesis cell. Chem. Eng. J. 2022, 446, 137079. [Google Scholar] [CrossRef]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011, 77, 2882–2886. [Google Scholar] [CrossRef] [PubMed]
- LaBelle, E.V.; May, H.D. Energy efficiency and productivity enhancement of microbial electrosynthesis of acetate. Front. Microbiol. 2017, 8, 756. [Google Scholar] [CrossRef] [PubMed]
- Mock, J.; Zheng, Y.; Mueller, A.P.; Ly, S.; Tran, L.; Segovia, S.; Nagaraju, S.; Köpke, M.; Dürre, P.; Thauer, R.K. Energy conservation associated with ethanol formation from H2 and CO2 in Clostridium autoethanogenum involving electron bifurcation. J. Bacteriol. 2015, 197, 2965–2980. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Kato, S. Extracellular electron transfer in acetogenic bacteria and its application for conversion of carbon dioxide into organic compounds. Appl. Microbiol. Biotechnol. 2017, 101, 6301–6307. [Google Scholar] [CrossRef] [PubMed]
- Madjarov, J.; Soares, R.; Paquete, C.M.; Louro, R.O. Sporomusa ovata as catalyst for bioelectrochemical carbon dioxide reduction: A review across disciplines from microbiology to process engineering. Front. Microbiol. 2022, 13, 913311. [Google Scholar] [CrossRef]
- Boto, S.T.; Bardl, B.; Harnisch, F.; Rosenbaum, M.A. Microbial electrosynthesis with Clostridium ljungdahlii benefits from hydrogen electron mediation and permits a greater variety of products. Green Chem. 2023, 25, 4375–4386. [Google Scholar] [CrossRef]
- Mills, S.; Dessì, P.; Pant, D.; Farràs, P.; Sloan, W.T.; Collins, G.; Ijaz, U.Z. A meta-analysis of acetogenic and methanogenic microbiomes in microbial electrosynthesis. NPJ Biofilms Microbiomes 2022, 8, 73. [Google Scholar] [CrossRef]
- Overmann, J.; Abt, B.; Sikorski, J. Present and future of culturing bacteria. Annu. Rev. Microbiol. 2017, 71, 711–730. [Google Scholar] [CrossRef]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A Toolkit to Classify genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef]
- Mayumi, D.; Mochimaru, H.; Yoshioka, H.; Sakata, S.; Maeda, H.; Miyagawa, Y.; Ikarashi, M.; Takeuchi, M.; Kamagata, Y. Evidence for syntrophic acetate oxidation coupled to hydrogenotrophic methanogenesis in the high-temperature petroleum reservoir of Yabase oil field (Japan). Environ. Microbiol. 2011, 13, 1995–2006. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I. Phylot: Phylogenetic Tree Generator. 2015. Available online: www.letunic.com (accessed on 11 November 2023).
- Coleman, G.A.; Davín, A.A.; Mahendrarajah, T.A.; Szánthó, L.L.; Spang, A.; Hugenholtz, P.; Szöllősi, G.J.; Williams, T.A. A rooted phylogeny resolves early bacterial evolution. Science 2021, 372, eabe0511. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Mendler, K.; Chen, H.; Parks, D.H.; Lobb, B.; Hug, L.A.; Doxey, A.C. AnnoTree: Visualization and exploration of a functionally annotated microbial tree of life. Nucleic Acids Res. 2019, 47, 4442–4448. [Google Scholar] [CrossRef]
- Søndergaard, D.; Pedersen, C.N.; Greening, C. HydDB: A web tool for hydrogenase classification and analysis. Sci. Rep. 2016, 6, 34212. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; ter Heijne, A.; Benetton, X.D.; Vanbroekhoven, K.; Buisman, C.J.; Strik, D.P.; Pant, D. Carbon dioxide reduction by mixed and pure cultures in microbial electrosynthesis using an assembly of graphite felt and stainless steel as a cathode. Bioresour. Technol. 2015, 195, 14–24. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Puniya, A.K.; Callaghan, T.M.; Edwards, J.E.; Youssef, N.; Dagar, S.S.; Fliegerova, K.; Griffith, G.W.; Forster, R.; Tsang, A. Anaerobic fungi (phylum Neocallimastigomycota): Advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiol. Ecol. 2014, 90, 1–17. [Google Scholar] [CrossRef]
- Deutzmann, J.r.S.; Kracke, F.; Gu, W.; Spormann, A.M. Microbial electrosynthesis of acetate powered by intermittent electricity. Environ. Sci. Technol. 2022, 56, 16073–16081. [Google Scholar] [CrossRef]
- Arends, J.B.; Patil, S.A.; Roume, H.; Rabaey, K. Continuous long-term electricity-driven bioproduction of carboxylates and isopropanol from CO2 with a mixed microbial community. J. CO2 Util. 2017, 20, 141–149. [Google Scholar] [CrossRef]
- Park, S.S.; DeCicco, B. Autotrophic growth with hydrogen of Mycobacterium gordonae and another scotochromogenic mycobacterium. Int. J. Syst. Evol. Microbiol. 1974, 24, 338–345. [Google Scholar] [CrossRef]
- Pett-Ridge, J.; Firestone, M. Redox fluctuation structures microbial communities in a wet tropical soil. Appl. Environ. Microbiol. 2005, 71, 6998–7007. [Google Scholar] [CrossRef] [PubMed]
- Philips, J. Extracellular electron uptake by acetogenic bacteria: Does H2 consumption favor the H2 evolution reaction on a cathode or metallic iron? Front. Microbiol. 2020, 10, 2997. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-M.; Wang, L.; Chen, L.; Tian, L.-J.; Zhu, T.-T.; Wu, Q.-Z.; Hu, Y.-R.; Zheng, L.-R.; Li, W.-W. AQDS activates extracellular synergistic biodetoxification of copper and selenite via altering the coordination environment of outer-membrane proteins. Environ. Sci. Technol. 2022, 56, 13786–13797. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, H.; Kahnt, J.; Thauer, R.K. A reversible electron-bifurcating ferredoxin-and NAD-dependent [FeFe]-hydrogenase (HydABC) in Moorella thermoacetica. J. Bacteriol. 2013, 195, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Schuchmann, K.; Müller, V. Energetics and application of heterotrophy in acetogenic bacteria. Appl. Environ. Microbiol. 2016, 82, 4056–4069. [Google Scholar] [CrossRef] [PubMed]
- Tanner, R.S.; Woese, C.R. A phylogenetic assessment of the acetogens. In Acetogenesis; Springer: Berlin/Heidelberg, Germany, 1994; pp. 254–269. [Google Scholar]
- Xavier, J.C.; Hordijk, W.; Kauffman, S.; Steel, M.; Martin, W.F. Autocatalytic chemical networks at the origin of metabolism. Proc. R. Soc. B 2020, 287, 20192377. [Google Scholar] [CrossRef] [PubMed]
- Basen, M.; Müller, V. Acetogens-from the origin of life to biotechnological applications. Front. Microbiol. 2023, 14, 1186930. [Google Scholar] [CrossRef]
- Tremblay, P.-L.; Faraghiparapari, N.; Zhang, T. Accelerated H2 evolution during microbial electrosynthesis with Sporomusa ovata. Catalysts 2019, 9, 166. [Google Scholar] [CrossRef]
- Holmes, D.E.; Zhou, J.; Ueki, T.; Woodard, T.; Lovley, D.R. Mechanisms for electron uptake by Methanosarcina acetivorans during direct interspecies electron transfer. MBio 2021, 12, e0234421. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef]
- Friedrich, T.; Abelmann, A.; Brors, B.; Guénebaut, V.; Kintscher, L.; Leonard, K.; Rasmussen, T.; Scheide, D.; Schlitt, A.; Schulte, U. Redox components and structure of the respiratory NADH: Ubiquinone oxidoreductase (complex I). Biochim. Biophys. Acta (BBA) Bioenerg. 1998, 1365, 215–219. [Google Scholar] [CrossRef]
- Watanabe, T.; Shima, S. MvhB-type polyferredoxin as an electron-transfer chain in putative redox-enzyme complexes. Chem. Lett. 2021, 50, 353–360. [Google Scholar] [CrossRef]
- Downing, B.E.; Gupta, D.; Nayak, D.D. The dual role of a multi-heme cytochrome in methanogenesis: MmcA is important for energy conservation and carbon metabolism in Methanosarcina acetivorans. Mol. Microbiol. 2023, 119, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Poudel, S.; Dunham, E.C.; Lindsay, M.R.; Amenabar, M.J.; Fones, E.M.; Colman, D.R.; Boyd, E.S. Origin and evolution of flavin-based electron bifurcating enzymes. Front. Microbiol. 2018, 9, 1762. [Google Scholar] [CrossRef] [PubMed]
- Mohanakrishna, G.; Reesh, I.M.A.; Vanbroekhoven, K.; Pant, D. Microbial electrosynthesis feasibility evaluation at high bicarbonate concentrations with enriched homoacetogenic biocathode. Sci. Total Environ. 2020, 715, 137003. [Google Scholar] [CrossRef]
- Aryal, N.; Tremblay, P.-L.; Lizak, D.M.; Zhang, T. Performance of different Sporomusa species for the microbial electrosynthesis of acetate from carbon dioxide. Bioresour. Technol. 2017, 233, 184–190. [Google Scholar] [CrossRef]
- Im, C.; Valgepea, K.; Modin, O.; Nygård, Y. Clostridium ljungdahlii as a biocatalyst in microbial electrosynthesis–Effect of culture conditions on product formation. Bioresour. Technol. Rep. 2022, 19, 101156. [Google Scholar]



| Run No. | Run-1E | Run-1N | Run-2E | Run-2N | Run-3E | Run-3N |
|---|---|---|---|---|---|---|
| Addition of voltage (mV) | −600 | Not added | −600 | Not added | −400 | Not added |
| Final conc. of CNT | 1% | 0.10% | 0.10% | |||
| Gelling agent | Gellan gum | Sodium silicate (Adj. pH 7.0) | Sodium silicate (Adj. pH 7.0) | |||
| Colony formation | + | + | − | − | + | − |
| Fungal colony formation | + | + | − | − | − | − |
| Cultivation period (weeks) | 4 | 4 | 12 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimura, Z.-i.; Kuriyama, H.; Iwasaki, Y. Exploring Acetogenesis in Firmicutes: From Phylogenetic Analysis to Solid Medium Cultivation with Solid-Phase Electrochemical Isolation Equipments. Microorganisms 2023, 11, 2976. https://doi.org/10.3390/microorganisms11122976
Kimura Z-i, Kuriyama H, Iwasaki Y. Exploring Acetogenesis in Firmicutes: From Phylogenetic Analysis to Solid Medium Cultivation with Solid-Phase Electrochemical Isolation Equipments. Microorganisms. 2023; 11(12):2976. https://doi.org/10.3390/microorganisms11122976
Chicago/Turabian StyleKimura, Zen-ichiro, Hiroki Kuriyama, and Yuki Iwasaki. 2023. "Exploring Acetogenesis in Firmicutes: From Phylogenetic Analysis to Solid Medium Cultivation with Solid-Phase Electrochemical Isolation Equipments" Microorganisms 11, no. 12: 2976. https://doi.org/10.3390/microorganisms11122976