
The Primary Mode of Action of Lippia graveolens Essential Oil on Salmonella enterica subsp. Enterica Serovar Typhimurium
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Essential Oil
2.2. GC-MS Analysis
2.3. Bacterial Strains and Growth Conditions
2.4. Antimicrobial Susceptibility Testing
2.4.1. Disc Diffusion Assays
2.4.2. Minimum Inhibitory Concentration and Minimum Bactericidal Concentration Assays
2.5. Time-Kill Studies
2.6. Cell Integrity Studies
2.7. Loss of Cytoplasmic Material
2.8. Determination of Released Proteins
2.9. Measurement of Intra- and Extracellular Adenosine 5′-Triphosphate (ATP) Concentrations
2.10. Membrane Depolarization Assay
2.11. Transmission Electron Microscopy (TEM)
2.12. Statistical Analysis
3. Results
3.1. Composition of Lippia graveolens EO of Plantas Cultivated in Cuba
3.2. Susceptibility of Salmonella ser. Typhimurium to Lippia graveolens Essential Oil
3.3. Time-Kill Studies
3.4. Cell Integrity Studies
3.5. Loss of Intracellular Material
3.6. Membrane Depolarization Assay
3.7. Measurement of Intra- and Extracellular Adenosine 5′-Triphosphate (ATP) Concentrations
3.8. Transmission Electron Microscopy (TEM)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WTO. International Forum on Food Safety and Trade. Available online: https://www.wto.org/english/tratop_e/sps_e/faowhowtoapril19_e.htm (accessed on 31 July 2023).
- Food Safety, Climate Change, and the Role of WHO. Available online: https://fctc.who.int/publications/i/item/food-safety-climate-change-and-the-role-of-who (accessed on 4 September 2023).
- WHO. Estimates of the Global Burden of Foodborne Diseases: Foodborne Diseases Burden Epidemiology Reference Group 2007–2015. Available online: https://www.who.int/publications-detail-redirect/9789241565165 (accessed on 4 September 2023).
- Lamas, A.; Miranda, J.M.; Regal, P.; Vázquez, B.; Franco, C.M.; Cepeda, A. A Comprehensive Review of Non-Enterica Subspecies of Salmonella Enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-Analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef]
- The Dark Side of the Salad: Salmonella Typhimurium Overcomes the Innate Immune Response of Arabidopsis Thaliana and Shows an Endopathogenic Lifestyle—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/18509467/ (accessed on 4 September 2023).
- Chen, Y.; Pouillot, R.; Santillana Farakos, S.M.; Duret, S.; Spungen, J.; Fu, T.-J.; Shakir, F.; Homola, P.A.; Dennis, S.; Van Doren, J.M. Risk Assessment of Salmonellosis from Consumption of Alfalfa Sprouts and Evaluation of the Public Health Impact of Sprout Seed Treatment and Spent Irrigation Water Testing. Risk Anal. 2018, 38, 1738–1757. [Google Scholar] [CrossRef]
- Cardoen, S.; Van Huffel, X.; Berkvens, D.; Quoilin, S.; Ducoffre, G.; Saegerman, C.; Speybroeck, N.; Imberechts, H.; Herman, L.; Ducatelle, R.; et al. Evidence-Based Semiquantitative Methodology for Prioritization of Foodborne Zoonoses. Foodborne Pathog. Dis. 2009, 6, 1083–1096. [Google Scholar] [CrossRef]
- Doyle, M.; Acheson, D.; Newland, J.; Dwelle, T.; Flynn, W.; Scott, H.M.; Singer, R.; Edge, M.S.; Flood, T. Enhancing Practitioner Knowledge about Antibiotic Resistance: Connecting Human and Animal Health. Food Prot. Trends 2016, 36, 390–394. [Google Scholar]
- Morganti, M.; Bolzoni, L.; Scaltriti, E.; Casadei, G.; Carra, E.; Rossi, L.; Gherardi, P.; Faccini, F.; Arrigoni, N.; Sacchi, A.R.; et al. Rise and Fall of Outbreak-Specific Clone inside Endemic Pulsotype of Salmonella 4,[5],12:I:-; Insights from High-Resolution Molecular Surveillance in Emilia-Romagna, Italy, 2012 to 2015. Euro Surveill 2018, 23, 17–00375. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Preedy, V.R. Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2015; p. 1013. [Google Scholar]
- Aziz, Z.A.A.; Ahmad, A.; Setapar, S.H.M.; Karakucuk, A.; Azim, M.M.; Lokhat, D.; Rafatullah, M.; Ganash, M.; Kamal, M.A.; Ashraf, G.M. Essential Oils: Extraction Techniques, Pharmaceutical and Therapeutic Potential—A Review. Curr. Drug Metab. 2018, 19, 1100–1110. [Google Scholar] [CrossRef]
- Guinoiseau, E.; Luciani, A.; de Rocca Serra, D.; Quilichini, Y.; Berti, L.; Lorenzi, V. Primary Mode of Action of Cistus Ladaniferus L. Essential Oil Active Fractions on Staphylococcus Aureus Strain. Adv. Microbiol. 2015, 5, 881. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential Oils of Oregano: Biological Activity beyond Their Antimicrobial Properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef]
- Almeida, M.C.; Pina, E.S.; Hernandes, C.; Zingaretti, S.M.; Taleb-Contini, S.H.; Salimena, F.R.G.; Slavov, S.N.; Haddad, S.K.; França, S.C.; Pereira, A.M.S.; et al. Genetic Diversity and Chemical Variability of Lippia spp. (Verbenaceae). BMC Res. Notes 2018, 11, 725. [Google Scholar] [CrossRef]
- Pascual, M.E.; Slowing, K.; Carretero, E.; Sánchez Mata, D.; Villar, A. Lippia: Traditional Uses, Chemistry and Pharmacology: A Review. J. Ethnopharmacol. 2001, 76, 201–214. [Google Scholar] [CrossRef]
- Hernández, T.; Canales, M.; Ávila, J.G.; García, A.; Meraz, S.; Caballero, J.; Lira, R. Composition and Antibacterial Activity of Essential Oil of Lippia Graveolens H.B.K. (Verbenaceae). Lat. Am. Caribb. Bull. Med. Aromat. Plants 2009, 8, 295–300. [Google Scholar]
- Arana-Sánchez, A.; Estarrón-Espinosa, M.; Obledo-Vázquez, E.N.; Padilla-Camberos, E.; Silva-Vázquez, R.; Lugo-Cervantes, E. Antimicrobial and Antioxidant Activities of Mexican Oregano Essential Oils (Lippia Graveolens H.B.K.) with Different Composition When Microencapsulated in Beta-Cyclodextrin. Lett. Appl. Microbiol. 2010, 50, 585–590. [Google Scholar] [CrossRef]
- Benachour, H.; Ramdani, M.; Lograda, T.; Chalard, P.; Figueredo, G. Chemical Composition and Antibacterial Activities of Capparis Spinosa Essential Oils from Algeria. Biodiversitas J. Biol. Divers. 2020, 21. [Google Scholar] [CrossRef]
- Klepser, M.E.; Ernst, E.J.; Lewis, R.E.; Ernst, M.E.; Pfaller, M.A. Influence of Test Conditions on Antifungal Time-Kill Curve Results: Proposal for Standardized Methods. Antimicrob. Agents Chemother. 1998, 42, 1207–1212. [Google Scholar] [CrossRef]
- Viljoen, A.; van Vuuren, S.; Ernst, E.; Klepser, M.; Demirci, B.; Başer, H.; van Wyk, B.E. Osmitopsis Asteriscoides (Asteraceae)-the Antimicrobial Activity and Essential Oil Composition of a Cape-Dutch Remedy. J. Ethnopharmacol. 2003, 88, 137–143. [Google Scholar] [CrossRef]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of Action of Melaleuca Alternifolia (Tea Tree) Oil on Staphylococcus Aureus Determined by Time-Kill, Lysis, Leakage, and Salt Tolerance Assays and Electron Microscopy. Antimicrob. Agents Chemother. 2002, 46, 1914–1920. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Gill, A.O.; Holley, R.A. Inhibition of Membrane Bound ATPases of Escherichia Coli and Listeria Monocytogenes by Plant Oil Aromatics. Int. J. Food Microbiol. 2006, 111, 170–174. [Google Scholar] [CrossRef]
- Turgis, M.; Han, J.; Caillet, S.; Lacroix, M. Antimicrobial Activity of Mustard Essential Oil against Escherichia Coli O157:H7 and Salmonella Typhi. Food Control 2009, 20, 1073–1079. [Google Scholar] [CrossRef]
- Chehimi, S.; Pons, A.-M.; Sablé, S.; Hajlaoui, M.-R.; Limam, F. Mode of Action of Thuricin S, a New Class IId Bacteriocin from Bacillus Thuringiensis. Can. J. Microbiol. 2010, 56, 162–167. [Google Scholar] [CrossRef]
- Mazzarrino, G.; Paparella, A.; Chaves-López, C.; Faberi, A.; Sergi, M.; Sigismondi, C.; Compagnone, D.; Serio, A. Salmonella Enterica and Listeria Monocytogenes Inactivation Dynamics after Treatment with Selected Essential Oils. Food Control 2015, 50, 794–803. [Google Scholar] [CrossRef]
- M26AE: Bactericidal Activity of Antimicrobial Agents. Available online: https://clsi.org/standards/products/microbiology/documents/m26/ (accessed on 4 September 2023).
- Chauhan, A.K.; Kang, S.C. Thymol Disrupts the Membrane Integrity of Salmonella Ser. Typhimurium in Vitro and Recovers Infected Macrophages from Oxidative Stress in an Ex Vivo Model. Res. Microbiol. 2014, 165, 559–565. [Google Scholar] [CrossRef]
- Saad, N.Y.; Muller, C.D.; Lobstein, A. Major Bioactivities and Mechanism of Action of Essential Oils and Their Components. Flavour Fragr. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Baek, K.-H.; Kang, S.C. Control of Salmonella in Foods by Using Essential Oils: A Review. Food Res. Int. 2012, 45, 722–734. [Google Scholar] [CrossRef]
- Salgueiro, L.R.; Cavaleiro, C.; Gonçalves, M.J.; da Cunha, A.P. Antimicrobial Activity and Chemical Composition of the Essential Oil of Lippia Graveolens from Guatemala. Planta Med. 2003, 69, 80–83. [Google Scholar] [CrossRef]
- Arias, J.; Mejía, J.; Córdoba, Y.; Martínez, J.R.; Stashenko, E.; del Valle, J.M. Optimization of Flavonoids Extraction from Lippia Graveolens and Lippia Origanoides Chemotypes with Ethanol-Modified Supercritical CO2 after Steam Distillation. Ind. Crops Prod. 2020, 146, 112170. [Google Scholar] [CrossRef]
- Hemaiswarya, S.; Doble, M. Synergistic Interaction of Eugenol with Antibiotics against Gram Negative Bacteria. Phytomedicine 2009, 16, 997–1005. [Google Scholar] [CrossRef]
- Yang, X.-N.; Khan, I.; Kang, S.C. Chemical Composition, Mechanism of Antibacterial Action and Antioxidant Activity of Leaf Essential Oil of Forsythia Koreana Deciduous Shrub. Asian Pac. J. Trop. Med. 2015, 8, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Kachur, K.; Suntres, Z. The Antibacterial Properties of Phenolic Isomers, Carvacrol and Thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Bennik, M.H.J.; Moezelaar, R. The Phenolic Hydroxyl Group of Carvacrol Is Essential for Action against the Food-Borne Pathogen Bacillus Cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial Properties of Plant Essential Oils against Human Pathogens and Their Mode of Action: An Updated Review. Available online: https://www.hindawi.com/journals/ecam/2016/3012462/ (accessed on 22 August 2018).
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed]
- Mempin, R.; Tran, H.; Chen, C.; Gong, H.; Kim Ho, K.; Lu, S. Release of Extracellular ATP by Bacteria during Growth. BMC Microbiol. 2013, 13, 301. [Google Scholar] [CrossRef]
- Stevens, M.J. Coarse-Grained Simulations of Lipid Bilayers. J. Chem. Phys. 2004, 121, 11942–11948. [Google Scholar] [CrossRef]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Martín-Belloso, O. Antimicrobial Activity of Essential Oils on Salmonella Enteritidis, Escherichia Coli, and Listeria Innocua in Fruit Juices. J. Food Prot. 2006, 69, 1579–1586. [Google Scholar] [CrossRef]
- Moosavy, M.-H.; Basti, A.A.; Misaghi, A.; Salehi, T.Z.; Abbasifar, R.; Mousavi, H.A.E.; Alipour, M.; Razavi, N.E.; Gandomi, H.; Noori, N. Effect of Zataria Multiflora Boiss. Essential Oil and Nisin on Salmonella Typhimurium and Staphylococcus Aureus in a Food Model System and on the Bacterial Cell Membranes. Food Res. Int. 2008, 41, 1050–1057. [Google Scholar] [CrossRef]
 ) at MIC and untreated cultures (
) at MIC and untreated cultures ( ). Mean values of triplicate independent experiments and standard deviations are shown. *** Significant differences between treatments compared to the control (p < 0.0001).
) at MIC and untreated cultures (). Mean values of triplicate independent experiments and standard deviations are shown. *** Significant differences between treatments compared to the control (p < 0.0001).
). Mean values of triplicate independent experiments and standard deviations are shown. *** Significant differences between treatments compared to the control (p < 0.0001).
) at MIC and untreated cultures (). Mean values of triplicate independent experiments and standard deviations are shown. *** Significant differences between treatments compared to the control (p < 0.0001).
 ) and after treatment with L. graveolens essential oil (
) and after treatment with L. graveolens essential oil ( ) at MIC. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (p < 0.05) in treatments at specific times and different lowercase letters show significant differences (p < 0.05) in the time of a specific treatment.
) and after treatment with L. graveolens essential oil () at MIC. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (p < 0.05) in treatments at specific times and different lowercase letters show significant differences (p < 0.05) in the time of a specific treatment.
) at MIC. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (p < 0.05) in treatments at specific times and different lowercase letters show significant differences (p < 0.05) in the time of a specific treatment.
) and after treatment with L. graveolens essential oil () at MIC. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (p < 0.05) in treatments at specific times and different lowercase letters show significant differences (p < 0.05) in the time of a specific treatment.
 ), with 30 mM PMB (
), with 30 mM PMB ( ) and 4 mM CTAB (
) and 4 mM CTAB ( ) monitored by fluorescence intensity change. Untreated cells were used as negative controls (
) monitored by fluorescence intensity change. Untreated cells were used as negative controls ( ). Mean values of triplicate independent experiments and standard deviations are shown. * Significant differences between treatments compared to the control (p < 0.05).
), with 30 mM PMB () and 4 mM CTAB () monitored by fluorescence intensity change. Untreated cells were used as negative controls (). Mean values of triplicate independent experiments and standard deviations are shown. * Significant differences between treatments compared to the control (p < 0.05).
). Mean values of triplicate independent experiments and standard deviations are shown. * Significant differences between treatments compared to the control (p < 0.05).
), with 30 mM PMB () and 4 mM CTAB () monitored by fluorescence intensity change. Untreated cells were used as negative controls (). Mean values of triplicate independent experiments and standard deviations are shown. * Significant differences between treatments compared to the control (p < 0.05).
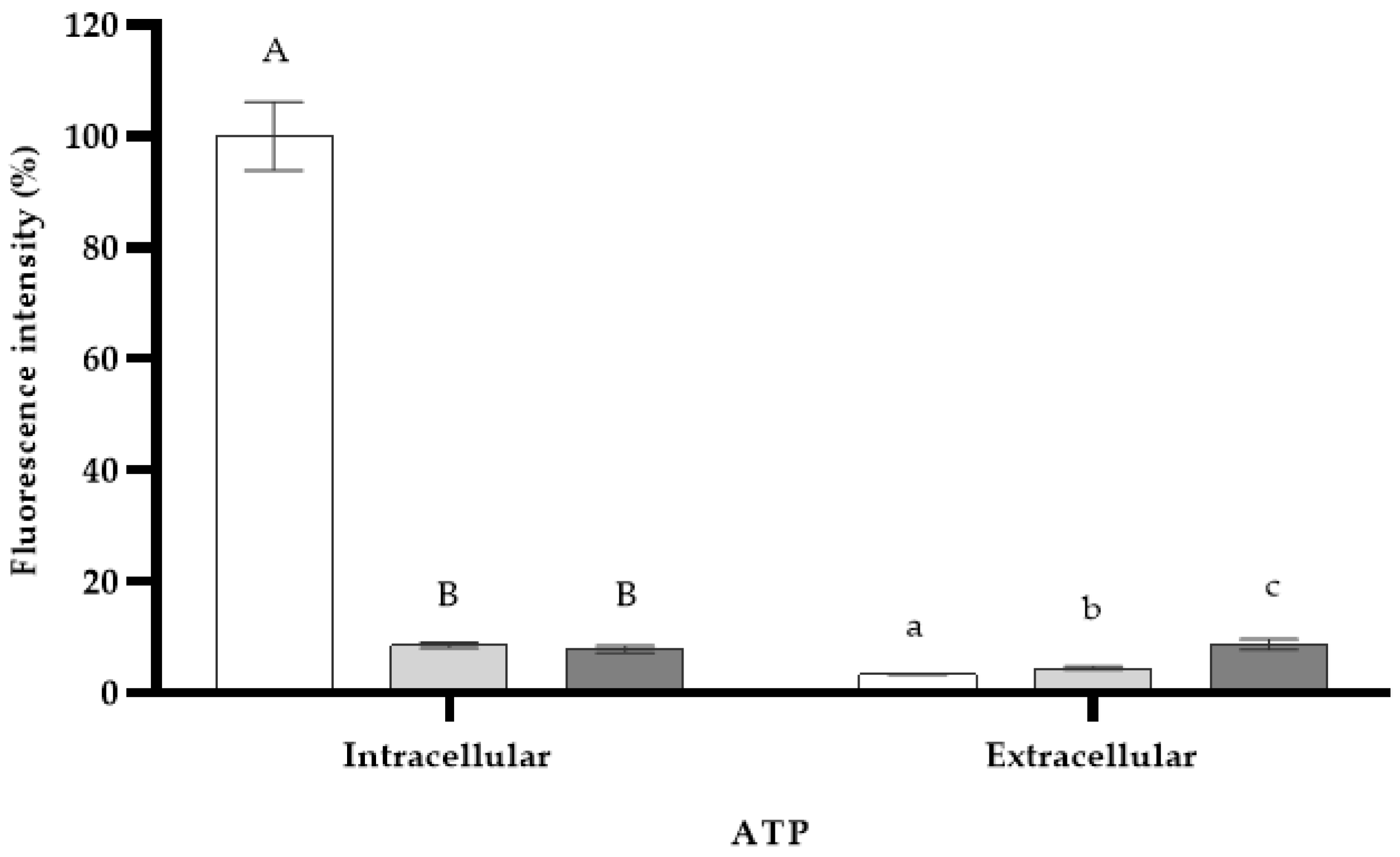
 ). DMSO (
). DMSO ( ) and PMB (
) and PMB ( ) were used as controls. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (DF = 2; F = 221.1; p < 0.0001) in intracellular ATP and different lowercase letters show significant differences (DF = 2; F = 23.0; p = 0.0015) in extracellular ATP.
). DMSO () and PMB () were used as controls. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (DF = 2; F = 221.1; p < 0.0001) in intracellular ATP and different lowercase letters show significant differences (DF = 2; F = 23.0; p = 0.0015) in extracellular ATP.
) were used as controls. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (DF = 2; F = 221.1; p < 0.0001) in intracellular ATP and different lowercase letters show significant differences (DF = 2; F = 23.0; p = 0.0015) in extracellular ATP.
). DMSO () and PMB () were used as controls. Mean values of triplicate independent experiments and standard deviations are shown. Different capital letters indicate significant differences (DF = 2; F = 221.1; p < 0.0001) in intracellular ATP and different lowercase letters show significant differences (DF = 2; F = 23.0; p = 0.0015) in extracellular ATP.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Retention Time (min) | Class of Compounds | Relative Abundance (%) | Match Quality |
|---|---|---|---|---|
| myrcene | 2.82 | mh | 1.3 | 96 |
| α-terpipene | 3.22 | mh | 1.3 | 98 |
| p-cymene | 3.38 | mh | 6.5 | 93 |
| 1,8-cineole | 3.49 | om | 4.0 | 99 |
| γ terpinene | 3.92 | mh | 7.3 | 96 |
| terpinen-4-ol | 5.80 | om | 1.1 | 97 |
| α-terpineol | 6.57 | om | 1.4 | 91 |
| thymol | 8.16 | om | 42.7 | 97 |
| carvacrol | 8.37 | om | 22.2 | 92 |
| (E)-caryophyllene (=β-caryophyllene) | 9.93 | sh | 3.5 | 99 |
| Wavelength | Time (min) | Released Material (%) | |
|---|---|---|---|
| Control | Lippia graveolens (0.4 mg/mL) | ||
| 260 nm | 0 30 60 90 120 | 0.00 ± 0.00 a 0.00 ± 0.00 a 0.01 ± 0.04 a 0.01 ± 0.04 a 0.00 ± 0.00 a | 0.00 ± 0.00 a 3.40 ± 1.17 b 7.37 ± 2.80 b 9.24 ± 3.56 b 11.10 ± 4.32 b |
| 280 nm | 0 30 60 90 120 | 0.00 ± 0.00 a 0.11 ± 0.13 a 0.40 ± 0.20 a 0.30 ± 0.19 a 0.11 ± 0.13 a | 0.00 ± 0.00 a 1.80 ± 0.49 ab 4.50 ± 1.62 b 7.50 ± 2.85 b 9.20 ± 3.55 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio Ortega, A.; Guinoiseau, E.; Poli, J.-P.; Quilichini, Y.; de Rocca Serra, D.; del Carmen Travieso Novelles, M.; Espinosa Castaño, I.; Pino Pérez, O.; Berti, L.; Lorenzi, V. The Primary Mode of Action of Lippia graveolens Essential Oil on Salmonella enterica subsp. Enterica Serovar Typhimurium. Microorganisms 2023, 11, 2943. https://doi.org/10.3390/microorganisms11122943
Rubio Ortega A, Guinoiseau E, Poli J-P, Quilichini Y, de Rocca Serra D, del Carmen Travieso Novelles M, Espinosa Castaño I, Pino Pérez O, Berti L, Lorenzi V. The Primary Mode of Action of Lippia graveolens Essential Oil on Salmonella enterica subsp. Enterica Serovar Typhimurium. Microorganisms. 2023; 11(12):2943. https://doi.org/10.3390/microorganisms11122943
Chicago/Turabian StyleRubio Ortega, Annie, Elodie Guinoiseau, Jean-Pierre Poli, Yann Quilichini, Dominique de Rocca Serra, Maria del Carmen Travieso Novelles, Ivette Espinosa Castaño, Oriela Pino Pérez, Liliane Berti, and Vannina Lorenzi. 2023. "The Primary Mode of Action of Lippia graveolens Essential Oil on Salmonella enterica subsp. Enterica Serovar Typhimurium" Microorganisms 11, no. 12: 2943. https://doi.org/10.3390/microorganisms11122943