
Modulation of Zebrafish (Danio rerio) Intestinal Mucosal Barrier Function Fed Different Postbiotics and a Probiotic from Lactobacilli
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Products and Experimental Diets
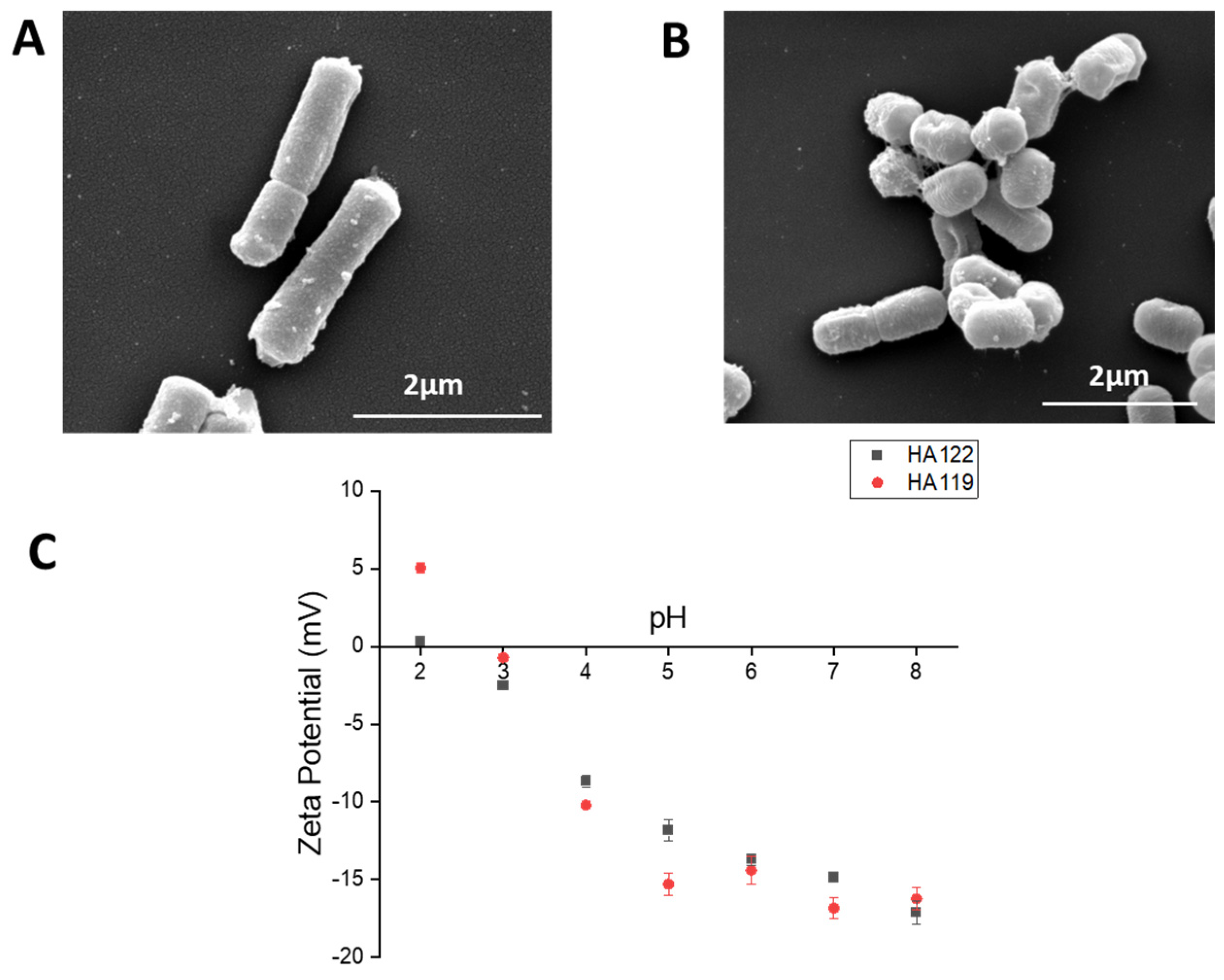
2.2. Characterization of Lactobacilli
2.3. Experimental Design and Feeding
2.4. Tissue Preparation and Light Microscopy
2.5. Intestinal Tissue Lysozyme and Cathepsin L
2.6. Gene Expression Analysis
2.7. Flow Cytometry of Intestinal Lymphoid Cells
2.8. Statistical Analysis
3. Results
3.1. Characterization of Postbiotics
3.2. Morphometric and Goblet Cell Chemotyping Show Strengthening of Mucosal Barrier
3.3. Mobilization CD4/CD8 Positive Cells
3.4. Modulation of Innate Intestinal Immune Responses by Postbiotic Groups
3.5. PCA Analysis of Immunity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; De Vos, W.M.; et al. Homeostasis of the gut barrier and potential biomarkers. Am. J. Physiol. Liver Physiol. 2017, 312, 93–171. [Google Scholar] [CrossRef]
- Schoultz, I.; Keita, Å.V. Cellular and molecular therapeutic targets in inflammatory bowel disease—Focusing on intestinal barrier function. Cells 2019, 8, 193. [Google Scholar] [CrossRef]
- Riedel, S.; Pheiffer, C.; Johnson, R.; Louw, J.; Muller, C.J. Intestinal barrier function and immune homeostasis are missing links in obesity and type 2 diabetes development. Front. Endocrinol. 2022, 12, 544–833. [Google Scholar] [CrossRef]
- Zhao, W.; Ho, H.E.; Bunyavanich, S. The gut microbiome in food allergy. Ann. Allergy Asthma Immunol. 2019, 122, 276–282. [Google Scholar] [CrossRef]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. BioEssays 2016, 38, 1167–1176. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Factories 2020, 19, 23. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van Der Sluis, M.; Bouma, J.; Boehm, G.; Van Goudoever, J.B.; Van Seuningen, I.; Renes, I.B. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: Implications for epithelial protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.; Graziani, C.; Guarino, A.; Lamborghini, A.; Masi, S.; Stanghellini, V. Gelatin tannate and tyndallized probiotics: A novel approach for treatment of diarrhea. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 873–883. [Google Scholar]
- Kataria, J.; Li, N.; Wynn, J.L.; Neu, J. Probiotic microbes: Do they need to be alive to be beneficial? Nutr. Rev. 2009, 67, 546–550. [Google Scholar] [CrossRef]
- Núria, P.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar]
- Taverniti, V.; Guglielmetti, S. Health-promoting properties of Lactobacillus helveticus. Front. Microbiol. 2012, 3, 392. [Google Scholar] [CrossRef]
- Mujagic, Z.; De Vos, P.; Boekschoten, M.V.; Govers, C.; Pieters, H.J.H.; De Wit, N.J.; Bron, P.A.; Masclee, A.A.; Troost, F.J. The effects of Lactobacillus plantarum on small intestinal barrier function and mucosal gene transcription; a randomized double-blind placebo controlled trial. Sci. Rep. 2017, 7, 401–428. [Google Scholar] [CrossRef]
- Paveljšek, D.; Ivičak-Kocjan, K.; Treven, P.; Benčina, M.; Jerala, R.; Rogelj, I. Distinctive probiotic features share common TLR2-dependent signalling in intestinal epithelial cells. Cell. Microbiol. 2021, 23, e13264. [Google Scholar] [CrossRef]
- Mohseni, A.H.; Casolaro, V.; Bermúdez-Humarán, L.G.; Keyvani, H.; Taghinezhad, S.S. Modulation of the PI3K/Akt/mTOR signalling pathway by probiotics as a fruitful target for orchestrating the immune response. Gut Microbes 2021, 13, 1886844. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, S.; Di, H.; Deng, Z.; Liu, J.; Wang, H. Gut health benefit and application of postbiotics in animal production. J. Anim. Sci. Biotechnol. 2022, 13, 38. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Cao, X.; Jin, X.; Jin, T. Pattern recognition receptors in zebrafish provide functional and evolutionary insight into innate immune signalling pathways. Cell. Mol. Immunol. 2017, 14, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Brugman, S. The zebrafish as a model to study intestinal inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutritional Requirements of Fish; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Association of Official Analytical Chemists—AOAC. Official Methods of Analysis, 18th ed.; AOAC INTERNATIONAL: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Caruana, J.C.; Dean, S.N.; Walper, S.A. Isolation and characterization of membrane vesicles from Lactobacillus species. Bio-Protocol 2021, 11, e4145. [Google Scholar] [CrossRef] [PubMed]
- Rawling, M.; Schiavone, M.; Apper, E.; Merrifield, D.L.; Castex, M.; Leclercq, E.; Foey, A. Yeast cell wall extracts from Saccharomyces cerevisiae varying in structure and composition differentially shape the innate immunity and mucosal tissue responses of the intestine of zebrafish (Danio rerio). Front. Immunol. 2023, 14, 1158390. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, E.; Pontefract, N.; Rawling, M.; Valdenegro, V.; Aasum, E.; Andujar, L.V.; Migaud, H.; Castex, M.; Merrifield, D. Dietary supplementation with a specific mannan-rich yeast parietal fraction enhances the gut and skin mucosal barriers of Atlantic salmon (Salmo salar) and reduces its susceptibility to sea lice (Lepeophtheirus salmonis). Aquaculture 2020, 529, 701–735. [Google Scholar] [CrossRef]
- Adams, A.; Thompson, K. Development of an enzyme-linked immunosorbent assay (ELISA) for the detection of Aeromonas salmonicida in fish tissue. J. Aquat. Anim. Health 1990, 2, 281–288. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Toda, H.; Shibasaki, Y.; Koike, T.; Ohtani, M.; Takizawa, F.; Ototake, M.; Moritomo, T.; Nakanishi, T. Alloantigen-specific killing is mediated by CD8-positive T cells in fish. Dev. Comp. Immunol. 2009, 33, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Toda, H.; Saito, Y.; Koike, T.; Takizawa, F.; Araki, K.; Yabu, T.; Somamoto, T.; Suetake, H.; Suzuki, Y.; Ototake, M.; et al. Conservation of characteristics and functions of CD4 positive lymphocytes in a teleost fish. Dev. Comp. Immunol. 2011, 35, 650–660. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.Rproject.org/ (accessed on 20 April 2023).
- Röhmel, J. Precision intervals for estimates of the difference in success rates for binary random variables based on the permutation principle. Biom. J. 1996, 38, 977–993. [Google Scholar] [CrossRef]
- Dohoo, I.R.; Ducrot, C.; Fourichon, C.; Donald, A.; Hurnik, D. An overview of techniques for dealing with large numbers of independent variables in epidemiologic studies. Prev. Vet. Med. 1997, 29, 221–239. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Waśko, A.; Polak-Berecka, M.; Kuzdraliński, A.; Skrzypek, T. Variability of S-layer proteins in Lactobacillus helveticus strains. Anaerobe 2014, 25, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Schär-Zammaretti, P.; Ubbink, J. The cell wall of lactic acid bacteria: Surface constituents and macromolecular conformations. Biophys. J. 2003, 85, 4076–4092. [Google Scholar] [CrossRef]
- Kurata, A.; Kiyohara, S.; Imai, T.; Yamasaki-Yashiki, S.; Zaima, N.; Moriyama, T.; Kishimoto, N.; Uegaki, K. Characterization of extracellular vesicles from Lactiplantibacillus plantarum. Sci. Rep. 2022, 12, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Dean, S.N.; Leary, D.H.; Sullivan, C.J.; Oh, E.; Walper, S.A. Isolation and characterization of Lactobacillus-derived membrane vesicles. Sci. Rep. 2019, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Mustapha, N.M.; Zulkifli, I.; Izuddin, W.I. Effects of feeding different postbiotics produced by Lactobacillus plantarum on growth performance, carcass yield, intestinal morphology, gut microbiota composition, immune status, and growth gene expression in broilers under heat stress. Animals 2019, 9, 644. [Google Scholar] [CrossRef]
- Matsuki, T.; Pédron, T.; Regnault, B.; Mulet, C.; Hara, T.; Sansonetti, P.J. Epithelial cell proliferation arrest induced by lactate and acetate from Lactobacillus casei and Bifidobacterium breve. PLoS ONE 2013, 8, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Khonyoung, D.; Yamauchi, K.E. Improved growth performance due to hypertrophied intestinal absorptive epithelial cells by heat-killed Lactobacillus sakei HS-1 in broiler chickens. J. Anim Sci. 2019, 97, 2066–2075. [Google Scholar] [CrossRef]
- Patnaude, L.; Mayo, M.; Mario, R.; Wu, X.; Knight, H.; Creamer, K.; Wilson, S.; Pivorunas, V.; Karman, J.; Phillips, L.; et al. Mechanisms and regulation of IL-22-mediated intestinal epithelial homeostasis and repair. Life Sci. 2021, 271, 119–195. [Google Scholar] [CrossRef]
- Ni, Y.; Zhang, Y.; Zheng, L.; Rong, N.; Yang, Y.; Gong, P.; Yang, Y.; Siwu, X.; Zhang, C.; Zhu, L.; et al. Bifidobacterium and Lactobacillus improve inflammatory bowel disease in zebrafish of different ages by regulating the intestinal mucosal barrier and microbiota. Life Sci. 2023, 324, 121–699. [Google Scholar] [CrossRef]
- Jariwala, R.; Mandal, H.; Bagchi, T. Indigenous lactobacilli strains of food and human sources reverse enteropathogenic E. coli O26: H11-induced damage in intestinal epithelial cell lines: Effect on redistribution of tight junction proteins. Microbiology 2017, 163, 1263–1272. [Google Scholar] [CrossRef]
- Deplancke, B.; Gaskins, H.R. Microbial modulation of innate defence: Goblet cells and the intestinal mucus layer. Am. J. Clin. Nutr. 2001, 73, 113–114. [Google Scholar] [CrossRef]
- Xie, S.; Zhao, S.; Jiang, L.; Lu, L.; Yang, Q.; Yu, Q. Lactobacillus reuteri stimulates intestinal epithelial proliferation and induces differentiation into goblet cells in young chickens. J. Agric. Food Chem. 2019, 67, 13758–13766. [Google Scholar] [CrossRef] [PubMed]
- Elahi, S.S.M.; Mirnejad, R.; Kazempoor, R.; Sotoodehnejadnematalahi, F. Study of the Histopathologic Effects of Probiotic Lactobacillus acidophilus in Exposure to E. coli O157: H7 in Zebrafish Intestine. Iran. Red Crescent Med. J. 2020, 22, 44–90. [Google Scholar]
- Lu, F.; Li, Y.; Wang, X.; Hu, X.; Liao, X.; Zhang, Y. Early-life polyphenol intake promotes Akkermansia growth and increase of host goblet cells in association with the potential synergistic effect of Lactobacillus. Food Res. Int. 2021, 149, 106–148. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, K.; Qi, W.; Zhou, Y.; Hong, T.; Xiong, T.; Xie, M.; Nie, S. Exopolysaccharides from Lactobacillus plantarum NCU116 enhances colonic mucosal homeostasis by controlling epithelial cell differentiation and c-Jun/Muc2 signalling. J. Agric. Food Chem. 2019, 67, 9831–9839. [Google Scholar] [CrossRef]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.Y. Antimicrobial peptides: New hope in the war against multidrug resistance. Zool. Res. 2019, 40, 488–505. [Google Scholar] [CrossRef]
- Schlee, M.; Harder, J.; Köten, B.; Stange, E.F.; Wehkamp, J.; Fellermann, K. Probiotic lactobacilli and VSL#3 induce enterocyte beta-defensin 2. Clin. Exp. Immunol. 2008, 151, 528–535. [Google Scholar]
- Möndel, M.; Schroeder, B.O.; Zimmermann, K.; Huber, H.; Nuding, S.; Beisner, J.; Fellermann, K.; Stange, E.F.; Wehkamp, J. Probiotic E. coli treatment mediates antimicrobial human β-defensin synthesis and fecal excretion in humans. Mucosal Immunol. 2009, 2, 166–172. [Google Scholar] [CrossRef]
- Becker, H.M.; Apladas, A.; Scharl, M.; Fried, M.; Rogler, G. Probiotic Escherichia coli Nissle 1917 and commensal E. coli K12 diferentially affect the infammasome in intestinal epithelial cells. Digestion 2014, 89, 110–118. [Google Scholar] [CrossRef]
- Rajakumari, D.; Viswanath, B.; Rani, A.U. Zebrafish: A novel model organism to assess probiotics influence on growth and development. In Recent Developments in Applied Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 2021; pp. 309–315. [Google Scholar]
- Qiu, Y.; Jiang, Z.; Hu, S.; Wang, L.; Ma, X.; Yang, X. Lactobacillus plantarum enhanced IL-22 production in natural killer (NK) cells that protect the integrity of intestinal epithelial cell barrier damaged by enterotoxigenic Escherichia coli. Int. J. Mol. Sci. 2017, 18, 2409. [Google Scholar] [CrossRef]
- Hou, Q.; Ye, L.; Liu, H.; Huang, L.; Yang, Q.; Turner, J.R.; Yu, Q. Lactobacillus accelerates ISCs regeneration to protect the integrity of intestinal mucosa through activation of STAT3 signalling pathway induced by LPLs secretion of IL-22. Cell Death Differ. 2018, 25, 1657–1670. [Google Scholar] [CrossRef]
- Hernández, P.P.; Strzelecka, P.M.; Athanasiadis, E.I.; Hall, D.; Robalo, A.F.; Collins, C.M.; Boudinot, P.; Levraud, J.P.; Cvejic, A. Single-cell transcriptional analysis reveals ILC-like cells in zebrafish. Sci. Immunol. 2018, 3, eaau5265. [Google Scholar] [CrossRef]
- Maeda, N.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Kase, T.; Yoshikai, Y. Oral administration of heat-killed Lactobacillus plantarum L-137 enhances protection against influenza virus infection by stimulation of type I interferon production in mice. Int. Immunopharmacol. 2009, 9, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Arimori, Y.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Shidara, O.; Ichikawa, H.; Yoshikai, Y. Daily intake of heat-killed Lactobacillus plantarum L-137 enhances type I interferon production in healthy humans and pigs. Immunopharmacol. Immunotoxicol. 2012, 34, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.C.; Lin, S.L.; Tsai, J.J.; Lin, M.Y. Heat-killed lactic acid bacteria enhance immunomodulatory potential by skewing the immune response toward Th1 polarization. J. Food Sci. 2011, 76, 260–267. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, 49–66. [Google Scholar] [CrossRef]
- Guo, S.; Gillingham, T.; Guo, Y.; Meng, D.; Zhu, W.; Walker, W.A.; Ganguli, K. Secretions of Bifidobacterium infantis and Lactobacillus acidophilus Protect Intestinal Epithelial Barrier Function. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.A. The probiotic paradox: Live and dead cells are biological response modifiers. Nutr. Res. Rev. 2010, 23, 37–46. [Google Scholar] [CrossRef]
- Hao, H.; Zhang, X.; Tong, L.; Liu, Q.; Liang, X.; Bu, Y.; Gong, P.; Liu, T.; Zhang, L.; Xia, Y.; et al. Effect of extracellular vesicles derived from Lactobacillus plantarum Q7 on gut microbiota and ulcerative colitis in mice. Front. Immunol. 2021, 12, 77147. [Google Scholar] [CrossRef]
- Khonyoung, D.; Yamauchi, K.E. Effects of heat-killed Lactobacillus plantarum L-137 on morphology of intestinal villi and epithelial cells in broiler chickens. J. Appl. Anim. Res. 2012, 40, 140–147. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Diet | |
|---|---|
| Wheat meal | 265.0 |
| Fabameal | 250.0 |
| Soybean meal | 150.0 |
| SPC60 | 239.0 |
| Rapeseed oil | 60.0 |
| Vitamin premix | 10.0 |
| Lysine HCL | 9.0 |
| DL methionine | 7.0 |
| Gelatin | 10.0 |
| Control | LPPro | LHPost | LPPost | |
|---|---|---|---|---|
| Dry matter (DM; %) | 90.1 ± 0.03 | 89.1 ± 0.09 | 89.9 ± 0.04 | 90.2 ± 0.12 |
| Crude Protein (% DM) | 39.6 ± 0.15 | 39.4 ± 0.23 | 39.9 ± 0.66 | 39.7 ± 0.73 |
| Crude Lipid (% DM) | 5.9 ± 0.09 | 5.3 ± 0.15 | 5.6 ± 0.05 | 5.9 ± 0.31 |
| Ash (% DM) | 4.0 ± 0.06 | 4.0 ± 0.05 | 4.1 ± 0.14 | 4.0 ± 0.01 |
| Gene | Functional Annotation | Accession Number | Primer Sequence (5′-3′) | Efficiency |
|---|---|---|---|---|
| Reference genes | ||||
| cops2 | COP9 signalosome subunit 2 | NM_001002055.1 | F-TCCAGATGTACACGCACAA RATCAGCCATCCCACATCCAC | 2.0 |
| metap1 | methionyl aminopeptidase 1 | NM_001025165.2 | F-GACGAGGGAGCCAAGGATT R-TCTGTGAAGCCTGGTATCCG | 2.0 |
| Mucosal barrier markers | ||||
| lyz | lysozyme | NM_139180.1 | F-ATGAAGGGCTGATGGATTGA R-GGTGGGTCTTAAACTGCTTTC | 2.1 |
| cath l | cathepsin L.1 | NM_001002368.1 | F-GGTGGACTGCTCTGGTTCTT R-CTCACCATCCTGGGCTTCAT | 2.0 |
| zo-2a | tight junction protein 2a (zona occludens 2) | NM_001201571.1 | F-CCTTGCTACCCAGTCCAGAAG R-GACGCAGACCAACGCTTTC | 2.0 |
| muc2.1 | mucin 2.1 | XM_021470771 | F-CTGTGCGGCTAAAGGATAATC R-CTCTGTGAGGCTGGGCAATC | 2.1 |
| Effector cytokines | ||||
| tnfα | tumor necrosis factor alpha | NM_212859.2 | F-CCATAAGACCCAGGCAATCA R-GGCAGCCTGGAAGTGAAATTG | 2.0 |
| il1β | interleukin 1, beta | NM_212844.2 | F-CGGAAGCAGCGACTTGAAAG R-AACACACAGGCTGAGCAGAA | 2.1 |
| ifnγ (ifng) | interferon gamma 1 | NM_212864.1 | F-CCCATCTTCCTGCGAATCCT R-GCTTCATCCACGCTGTCATTC | 2.1 |
| il22 | interleukin 22 | NM_001020792.1 | F-TGGAATCAGACGAGCACACA R-GGCTGGAGTAGTCGTGTTTACA | 2.1 |
| il17a | interleukin 17a/f1 | NM_001020787.1 | F-ACATAACGAGAGCCTGTATCCT R-CCTCAACGCCGTCTATCAGA | 2.0 |
| tgfβ | transforming growth factor, beta 2 | NM_194386.2 | F-AGGACAACACTGAGACTGAGTA R-GCAGTAGGGCAGGTCATTGT | 2.1 |
| Control | LHPro | LHPost | LPPost | p-Value | |
|---|---|---|---|---|---|
| Morphological parameter | |||||
| Muscularis thickness (µm) | 8.6 ± 1.9 | 9.9 ± 2.9 | 8.4 ± 1.7 | 10.2 ± 2.7 | 0.29 |
| Laminar propria width (µm) | 10.3 ± 1.6 a | 10.1 ± 0.8 ab | 8.5 ± 1.4 b | 9.0 ± 1.0 ab | 0.03 |
| Villi length (µm) | 114.7 ± 31.0 a | 158.6 ± 26.0 b | 137.2 ± 27.0 ab | 171.9 ± 47.0 b | 0.01 |
| IEL density (n/100 Enterocytes) | 9.0 ± 0.7 a | 11.7 ± 1.4 b | 15.9 ± 2.9 c | 11.7 ± 1.1 b | 0.01 |
| Goblet cell density (n/200 µm) | 12.0 ± 4.1 a | 11.3 ± 1.9 ac | 15.6 ± 4.4 bc | 13.7 ± 1.8 b | 0.02 |
| Goblet cell coverage (%) | 10.3 ± 2.2 a | 7.7 ± 2.4 b | 12.4 ± 7.3 abc | 17.1 ± 6.1 c | <0.05 |
| Goblet cell chemotyping | |||||
| Acido mucins (%) | 66.0 ± 4.1 a | 70.3 ± 2.5 ab | 76.1 ± 3.1 bc | 79.3 ± 1.6 c | <0.05 |
| Neutral mucins (%) | 17.9 ± 2.0 a | 15.0 ± 1.5 ab | 11.6 ± 2.0 b | 6.2 ± 1.0 c | <0.05 |
| Both acidic and neutral mucins (%) | 16.1 ± 2.3 | 14.7 ± 1.2 | 12.2 ± 1.9 | 14.4 ± 1.2 | 0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawling, M.; Schiavone, M.; Mugnier, A.; Leclercq, E.; Merrifield, D.; Foey, A.; Apper, E. Modulation of Zebrafish (Danio rerio) Intestinal Mucosal Barrier Function Fed Different Postbiotics and a Probiotic from Lactobacilli. Microorganisms 2023, 11, 2900. https://doi.org/10.3390/microorganisms11122900
Rawling M, Schiavone M, Mugnier A, Leclercq E, Merrifield D, Foey A, Apper E. Modulation of Zebrafish (Danio rerio) Intestinal Mucosal Barrier Function Fed Different Postbiotics and a Probiotic from Lactobacilli. Microorganisms. 2023; 11(12):2900. https://doi.org/10.3390/microorganisms11122900
Chicago/Turabian StyleRawling, Mark, Marion Schiavone, Amélie Mugnier, Eric Leclercq, Daniel Merrifield, Andrew Foey, and Emmanuelle Apper. 2023. "Modulation of Zebrafish (Danio rerio) Intestinal Mucosal Barrier Function Fed Different Postbiotics and a Probiotic from Lactobacilli" Microorganisms 11, no. 12: 2900. https://doi.org/10.3390/microorganisms11122900