
Exploring Antimicrobial Resistance Profiles of E. coli Isolates in Dairy Cattle: A Baseline Study across Dairy Farms with Varied Husbandry Practices in Puerto Rico

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Clearance
2.2. Samples and Processing
2.3. E. coli Identification
2.4. Antibiotic Susceptibility Testing
2.5. DNA Extraction
2.6. Whole Genome Sequencing and Bioinformatic Analyses
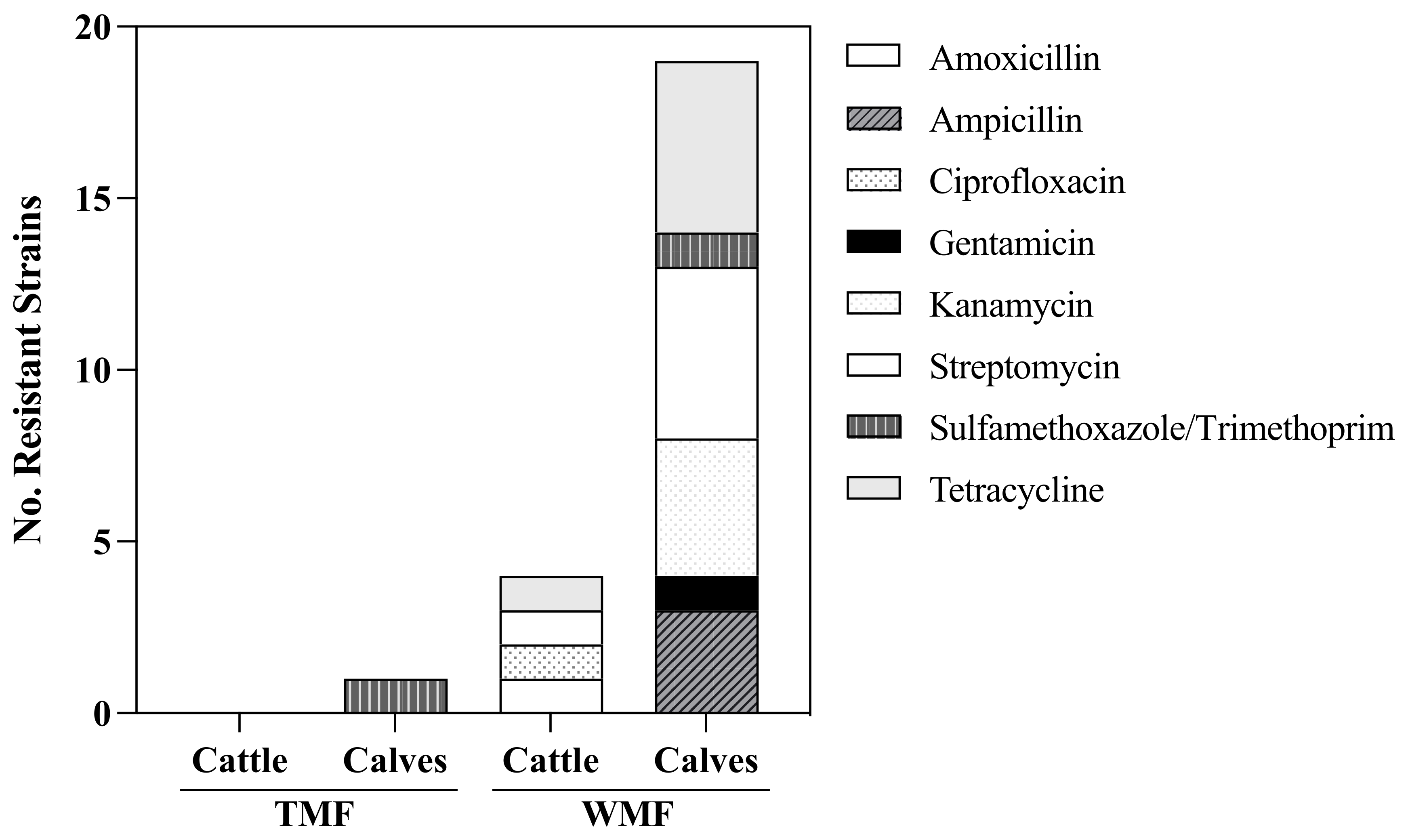
3. Results
Phenotypic and In Silico Antibiotic Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sneeringer, S.; MacDonald, J.M.; Key, N.; McBride, W.D.; Mathews, K. Economics of Antibiotic Use in U.S. Livestock Production; US Department of Agriculture: Washington, DC, USA, 2017. Available online: https://www.ers.usda.gov/webdocs/publications/45485/err-200.pdf (accessed on 6 November 2023).
- Abdus Sobur, M.; Al Momen Sabuj, A.; Sarker, R.; Taufiqur Rahman, A.M.M.; Lutful Kabir, S.M.; Tanvir Rahman, M. Antibiotic-Resistant Escherichia coli and Salmonella spp. Associated with Dairy Cattle and Farm Environment Having Public Health Significance. Vet. World 2019, 12, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Guo, X.; Xu, J.; Kong, X.; Gao, S.; Shan, Z. Pollution Characteristics and Environmental Risk Assessment of Typical Veterinary Antibiotics in Livestock Farms in Southeastern China. J. Environ. Sci. Health Part B 2014, 49, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Critically Important Antimicrobials for Human Medicine. 6th Revision. 2018 Ranking of Medically Important Antimicrobials for Risk Management of Antimicrobial Resistance Due to Non-Human Use. Available online: https://www.who.int/publications/i/item/9789241515528 (accessed on 6 November 2023).
- FDA. Summary Report on Antimicrobials Sold or Distributed for Use in Food-Producing Animals. 2017. Available online: https://www.fda.gov/media/119332/download (accessed on 6 November 2023).
- Provenza, F.D.; Kronberg, S.L.; Gregorini, P. Is Grassfed Meat and Dairy Better for Human and Environmental Health? Front. Nutr. 2019, 6, 431052. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Taft, D.H.; Maldonado-Gomez, M.X.; Johnson, D.; Treiber, M.L.; Lemay, D.G.; DePeters, E.J.; Mills, D.A. The Fecal Resistome of Dairy Cattle Is Associated with Diet during Nursing. Nat. Commun. 2019, 10, 4406. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Choi, S.H.; Nogoy, K.M.; Liang, S. Review: The Development of the Gastrointestinal Tract Microbiota and Intervention in Neonatal Ruminants. Animal 2021, 15, 100316. [Google Scholar] [CrossRef] [PubMed]
- Maynou, G.; Bach, A.; Terré, M. Feeding of Waste Milk to Holstein Calves Affects Antimicrobial Resistance of Escherichia coli and Pasteurella multocida Isolated from Fecal and Nasal Swabs. J. Dairy Sci. 2017, 100, 2682–2694. [Google Scholar] [CrossRef] [PubMed]
- Duse, A.; Waller, K.P.; Emanuelson, U.; Unnerstad, H.E.; Persson, Y.; Bengtsson, B. Risk Factors for Antimicrobial Resistance in Fecal Escherichia coli from Preweaned Dairy Calves. J. Dairy Sci. 2015, 98, 500–516. [Google Scholar] [CrossRef]
- Revista Del SEA 2017—Vol 3—Servicio de Extensión Agrícola. Available online: https://www.uprm.edu/sea/mdocs-posts/sea-del-oeste-vol-1-ene-mar/ (accessed on 6 November 2023).
- Urie, N.J.; Lombard, J.E.; Shivley, C.B.; Kopral, C.A.; Adams, A.E.; Earleywine, T.J.; Olson, J.D.; Garry, F.B. Preweaned Heifer Management on US Dairy Operations: Part I. Descriptive Characteristics of Preweaned Heifer Raising Practices. J. Dairy Sci. 2018, 101, 9168–9184. [Google Scholar] [CrossRef]
- Belmar, M.; Aly, S.; Karle, B.M.; Pereira, R.V. Evaluation of Four Commercial Tests for Detecting Ceftiofur in Waste Milk Bulk Tank Samples. PLoS ONE 2019, 14, e0224884. [Google Scholar] [CrossRef]
- Tempini, P.N.; Aly, S.S.; Karle, B.M.; Pereira, R.V. Multidrug Residues and Antimicrobial Resistance Patterns in Waste Milk from Dairy Farms in Central California. J. Dairy Sci. 2018, 101, 8110–8122. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fern Andez Esc Amez, P.S.; Girones, R.; Koutsoumanis, K.; Lindqvist, R.; et al. Risk for the Development of Antimicrobial Resistance (AMR) Due to Feeding of Calves with Milk Containing Residues of Antibiotics. EFSA J. 2017, 15, e04665. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Kenner, B.; Russell, D.; Valdes, C.; Sowell, A.; Pham, X.; Terán, A.; Kaufman, J. Economic Research Service Administrative Publication Number 114 Puerto Rico’s Agricultural Economy in the Aftermath of Hurricanes Irma and Maria: A Brief Overview. 2023. Available online: https://www.ers.usda.gov/publications/pub-details/?pubid=106260 (accessed on 6 November 2023).
- USDA National Agricultural Statistics Service (USDA NASS): 2017 Census of Agriculture. Available online: https://www.nass.usda.gov/AgCensus/ (accessed on 27 April 2022).
- Anjum, M.F.; Schmitt, H.; Börjesson, S.; Berendonk, T.U.; Donner, E.; Stehling, E.G.; Boerlin, P.; Topp, E.; Jardine, C.; Li, X.; et al. The Potential of Using E. coli as an Indicator for the Surveillance of Antimicrobial Resistance (AMR) in the Environment. Curr. Opin. Microbiol. 2021, 64, 152–158. [Google Scholar] [CrossRef]
- Jang, J.; Hur, H.G.; Sadowsky, M.J.; Byappanahalli, M.N.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and Public Health Implications—A Review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [PubMed]
- Omisakin, F.; MacRae, M.; Ogden, I.D.; Strachan, N.J.C. Concentration and Prevalence of Escherichia coli O157 in Cattle Feces at Slaughter. Appl. Environ. Microbiol. 2003, 69, 2444–2447. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Teixeira, A.G.V.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C. Fecal Microbial Diversity in Pre-Weaned Dairy Calves as Described by Pyrosequencing of Metagenomic 16S RDNA. Associations of Faecalibacterium Species with Health and Growth. PLoS ONE 2013, 8, e63157. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Weinstein, M.P. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Gangiredla, J.; Rand, H.; Benisatto, D.; Payne, J.; Strittmatter, C.; Sanders, J.; Wolfgang, W.J.; Libuit, K.; Herrick, J.B.; Prarat, M.; et al. GalaxyTrakr: A Distributed Analysis Tool for Public Health Whole Genome Sequence Data Accessible to Non-Bioinformaticians. BMC Genom. 2021, 22, 114. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- GitHub—Tseemann/Mlst: Scan Contig Files against PubMLST Typing Schemes. Available online: https://github.com/tseemann/mlst (accessed on 27 April 2022).
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Bessonov, K.; Laing, C.; Robertson, J.; Yong, I.; Ziebell, K.; Gannon, V.P.J.; Nichani, A.; Arya, G.; Nash, J.H.E.; Christianson, S. ECTyper: In Silico Escherichia coli Serotype and Species Prediction from Raw and Assembled Whole-Genome Sequence Data. Microb. Genom. 2021, 7, 000728. [Google Scholar] [CrossRef]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef]
- Brunton, L.A.; Reeves, H.E.; Snow, L.C.; Jones, J.R. A Longitudinal Field Trial Assesing the Impact of Feeding Waste Milk Containing Antibiotic Residues on the Prevalence of ESBL-Producing Escherichia coli in Calves. Prev. Vet. Med. 2014, 117, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.V.V.; Carroll, L.M.; Lima, S.; Foditsch, C.; Siler, J.D.; Bicalho, R.C.; Warnick, L.D. Impacts of Feeding Preweaned Calves Milk Containing Drug Residues on the Functional Profile of the Fecal Microbiota. Sci. Rep. 2018, 8, 554. [Google Scholar] [CrossRef] [PubMed]
- Aust, V.; Knappstein, K.; Kunz, H.J.; Kaspar, H.; Wallmann, J.; Kaske, M. Feeding Untreated and Pasteurized Waste Milk and Bulk Milk to Calves: Effects on Calf Performance, Health Status and Antibiotic Resistance of Faecal Bacteria. J. Anim. Physiol. Anim. Nutr. 2013, 97, 1091–1103. [Google Scholar] [CrossRef]
- Langford, F.M.; Weary, D.M.; Fisher, L. Antibiotic Resistance in Gut Bacteria from Dairy Calves: A Dose Response to the Level of Antibiotics Fed in Milk. J. Dairy Sci. 2003, 86, 3963–3966. [Google Scholar] [CrossRef] [PubMed]
- Gelalcha, B.D.; Agga, G.E.; Dego, O.K. Antimicrobial Usage for the Management of Mastitis in the USA: Impacts on Antimicrobial Resistance and Potential Alternative Approaches. In Mastitis in Dairy Cattle, Sheep and Goats; Dego, O.K., Ed.; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar]
- Gaire, T.N.; Scott, H.M.; Sellers, L.; Nagaraja, T.G.; Volkova, V.V. Age Dependence of Antimicrobial Resistance among Fecal Bacteria in Animals: A Scoping Review. Front. Vet. Sci. 2021, 7, 622495. [Google Scholar] [CrossRef]
- Goulart, D.B.; Mellata, M. Escherichia Coli Mastitis in Dairy Cattle: Etiology, Diagnosis, and Treatment Challenges. Front. Microbiol. 2022, 13, 928346. [Google Scholar] [CrossRef]
- Edgar, R.; Bibi, E. MdfA, an Escherichia coli Multidrug Resistance Protein with an Extraordinarily Broad Spectrum of Drug Recognition. J. Bacteriol. 1997, 179, 2274–2280. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Meng, J.; McDermott, P.F.; Wang, F.; Yang, Q.; Cao, G.; Hoffmann, M.; Zhao, S. Presence of Disinfectant Resistance Genes in Escherichia coli Isolated from Retail Meats in the USA. J. Antimicrob. Chemother. 2014, 69, 2644–2649. [Google Scholar] [CrossRef] [PubMed]
- Eibl, C.; Bexiga, R.; Viora, L.; Guyot, H.; Félix, J.; Wilms, J.; Tichy, A.; Hund, A. The Antibiotic Treatment of Calf Diarrhea in Four European Countries: A Survey. Antibiotics 2021, 10, 910. [Google Scholar] [CrossRef]
- Blickwede, M.; Schwarz, S. Molecular Analysis of Florfenicol-Resistant Escherichia coli Isolates from Pigs. J. Antimicrob. Chemother. 2004, 53, 58–64. [Google Scholar] [CrossRef]
- Meunier, D.; Jouy, E.; Lazizzera, C.; Doublet, B.; Kobisch, M.; Cloeckaert, A.; Madec, J.Y. Plasmid-Borne Florfenicol and Ceftiofur Resistance Encoded by the FloR and BlaCMY-2 Genes in Escherichia coli Isolates from Diseased Cattle in France. J. Med. Microbiol. 2010, 59, 467–471. [Google Scholar] [CrossRef] [PubMed]
- White, D.G.; Hudson, C.; Maurer, J.J.; Ayers, S.; Zhao, S.; Lee, M.D.; Bolton, L.; Foley, T.; Sherwood, J. Characterization of Chloramphenicol and Florfenicol Resistance in Escherichia coli Associated with Bovine Diarrhea. J. Clin. Microbiol. 2000, 38, 4593–4598. [Google Scholar] [CrossRef] [PubMed]
- Hartzell, A.; Chen, C.; Lewis, C.; Liu, K.; Reynolds, S.; Dudley, E.G. Escherichia coli O157:H7 of Genotype Lineage-Specific Polymorphism Assay 211111 and Clade 8 Are Common Clinical Isolates Within Pennsylvania. Foodborne Pathog. Dis. 2011, 8, 763–768. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Mohamed, K.; Fan, Y.; Achtman, M.; The Agama Study Group; Brown, D.; Chattaway, M.; Dallman, T.; Delahay, R.; et al. The EnteroBase User’s Guide, with Case Studies on Salmonella Transmissions, Yersinia Pestis Phylogeny, and Escherichia Core Genomic Diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef]
- Habets, A.; Touzain, F.; Lucas, P.; Huong, N.T.T.; Iguchi, A.; Crombé, F.; Korsak, N.; Piérard, D.; Saulmont, M.; Cox, E.; et al. Identification of Five Serotypes of Enteropathogenic Escherichia coli from Diarrheic Calves and Healthy Cattle in Belgium and Comparative Genomics with Shigatoxigenic E. coli. Vet. Sci. 2022, 9, 492. [Google Scholar] [CrossRef]
- Stoesser, N.; Sheppard, A.E.; Moore, C.E.; Golubchik, T.; Parry, C.M.; Nget, P.; Saroeun, M.; Day, N.P.J.; Giess, A.; Johnson, J.R.; et al. Extensive Within-Host Diversity in Fecally Carried Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolates: Implications for Transmission Analyses. J. Clin. Microbiol. 2015, 53, 2122–2131. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, V.; de Lurdes Enes Dapkevicius, M.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals 2020, 10, 2239. [Google Scholar] [CrossRef] [PubMed]
- Baharoglu, Z.; Garriss, G.; Mazel, D. Multiple Pathways of Genome Plasticity Leading to Development of Antibiotic Resistance. Antibiotics 2013, 2, 288–315. [Google Scholar] [CrossRef] [PubMed]
- Muloi, D.M.; Wee, B.A.; McClean, D.M.H.; Ward, M.J.; Pankhurst, L.; Phan, H.; Ivens, A.C.; Kivali, V.; Kiyong’a, A.; Ndinda, C.; et al. Population Genomics of Escherichia coli in Livestock-Keeping Households across a Rapidly Developing Urban Landscape. Nat. Microbiol. 2022, 7, 581–589. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, S.J.; Lissemore, K.D.; Kelton, D.F.; Duffield, T.F.; Leslie, K.E. Major Advances in Disease Prevention in Dairy Cattle. J. Dairy Sci. 2006, 89, 1267–1279. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antibiotic Family | Gene | TMF | WMF | Total Number (%) | ||
|---|---|---|---|---|---|---|
| Cattle (n = 8) | Calves (n = 8) | Cattle (n = 8) | Calves (n = 8) | |||
| No. Positive Strains | ||||||
| Aminoglycoside | aac(3)-Ild | -* | -* | -* | 1 | 16 (20) |
| aadA1 | -* | -* | -* | 2 | ||
| aadA2 | -* | -* | -* | 1 | ||
| aadA5 | -* | 1 | -* | -* | ||
| aph(3′)-Ia | -* | -* | -* | 3 | ||
| aph(3″)-Ib | -* | -* | 1 | 3 | ||
| aph(6)-ld | -* | -* | 1 | 3 | ||
| Beta-lactam | blaTEM-1B | -* | -* | -* | 1 | 5 (6) |
| blaTEM-57 | -* | -* | -* | 3 | ||
| blaCTX-M1 | -* | -* | -* | 1 | ||
| Folate pathway antagonist | dfrA12 | -* | -* | -* | 1 | 2 (3) |
| dfrA17 | -* | 1 | -* | -* | ||
| Phenicol | floR | -* | -* | 1 | 5 | 6 (8) |
| Macrolide | mdfA | 8 | 8 | 8 | 8 | 32 (41) |
| Quinolone | qnrB19 | -* | -* | 1 | 3 | 4 (5) |
| Sulfonamide | sul1 | -* | -* | -* | 1 | 6 (8) |
| sul2 | -* | 1 | -* | 3 | ||
| sul3 | -* | -* | -* | 1 | ||
| Tetracycline | tet(A) | -* | -* | 1 | 5 | 9 (11) |
| tet(B) | -* | -* | -* | 3 | ||
| All AMR genes number (%) | 8 (10) | 11 (14) | 13 (16) | 48 (60) | ||
| Animal | Sample ID | SRR Accession No. | AMR Gene Profile | Predicted Resistance | AMR Phenotype | Phylogroup | Serotype | ST |
|---|---|---|---|---|---|---|---|---|
| TMF Cattle | UPR-0066 | SRR10987123 | mdf(A) | MCL | -* | B1 | O150:H8 | 906 |
| UPR-0069 | SRR10987153 | mdf(A) | MCL | -* | B1 | O117:H9 | 10783 | |
| UPR-0072 | SRR10990262 | mdf(A) | MCL | -* | B1 | OXY60:H21 | 56 | |
| UPR-0073 | SRR10987140 | mdf(A) | MCL | -* | B1 | SB13:H3 | 4623 | |
| UPR-0074 | SRR10987149 | mdf(A) | MCL | -* | D | O1:H18 | 38 | |
| UPR-0075 | SRR10987248 | mdf(A) | MCL | -* | A | O66:H25 | 2325 | |
| UPR-0077 | SRR10987249 | mdf(A) | MCL | -* | E | O158:H9 | 10761 | |
| UPR-0078 | SRR10987152 | mdf(A) | MCL | -* | B1 | O88:H25 | 5765 | |
| TMF Calves | UPR-0079 | SRR10987160 | mdf(A) | MCL | -* | B1 | O149:H8 | 4392 |
| UPR-0080 | SRR10990269 | mdf(A) | MCL | -* | B2 | O40:H30 | 8217 | |
| UPR-0081 | SRR10987150 | aadA5, dfrA17, mdf(A), sul2 | MCL, SMZ, STM, TMP | STM, SXT | B1 | OXY13:H40 | 155 | |
| UPR-0082 | SRR10990363 | mdf(A) | MCL | -* | B1 | O8:H30 | 8217 | |
| UPR-0083 | SRR10987132 | mdf(A) | MCL | -* | B1 | O40:H30 | 8217 | |
| UPR-0084 | SRR10987130 | mdf(A) | MCL | -* | B1 | O8:H30 | 8217 | |
| UPR-0085 | SRR10987119 | mdf(A) | MCL | -* | A | O177:H11 | 765 | |
| UPR-0087 | SRR10987158 | mdf(A) | MCL | -* | A | O1:H18 | 38 | |
| WMF Cattle | UPR-0037 | SRR10990315 | mdf(A) | MCL | -* | B1 | O90:H7 | 3580 |
| UPR-0038 | SRR10987121 | mdf(A) | MCL | -* | B1 | O21:H21 | 101 | |
| UPR-0039 | SRR10990265 | mdf(A) | MCL | -* | B1 | O90:H7 | 3580 | |
| UPR-0042 | SRR10987156 | mdf(A) | MCL | -* | B1 | OXY38:H7 | 10759 | |
| UPR-0043 | SRR10990242 | aph(3″)-Ib, aph(6)-Id, mdf(A), floR, qnrB19, tet(A) | CHL, CIP, FLO, MCL, STM, TET | AMC, STM, TET | D | O141:H32 | 1721 | |
| UPR-0045 | SRR10987145 | mdf(A) | MCL | -* | A | O174:H16 | 2280 | |
| UPR-0050 | SRR10990246 | mdf(A) | MCL | -* | E | OXY20:H28 | 942 | |
| UPR-0051 | SRR10987139 | mdf(A) | MCL | -* | B1 | O53:H45 | 10760 | |
| WMF Calves | UPR-0053 | SRR10987157 | mdf(A) | MCL | -* | B1 | O177:H11 | 765 |
| UPR-0054 | SRR10987162 | mdf(A) | MCL | -* | A | O53:H51 | 5768 | |
| UPR-0055 | SRR10987159 | aac(3)-IId, aph(3′)-Ia, aph(3″)-Ib, aph(6)-Id, blaCTX-M-1, blaTEM-57, floR, mdf(A), sul2, tet(A), tet(B) | AMC, CHL, ERT, GEN, KAN, MCL, STM, TET | AMP, GEN, KAN, STM, TET | B1 | O154:H25 | 58 | |
| UPR-0058 | SRR10987154 | ant(3″)-Ia, aph(3′)-Ia, floR, qnrB19, mdf(A), sul3, tet(A), tet(B) | CHL, CIP, KAN, MCL, SMZ, STM, TET | KAN, STM, TET | A | O108:H34 | 10598 | |
| UPR-0059 | SRR10987133 | ant(3″)-Ia, aph(3′)-Ia, aph(3″)-Ib, aph(6)-Id, blaCTX-M-1, blaTEM-57, floR, qnrB19, mdf(A), sul2, sul3, tet(A), tet(B) | AMP, CIP, ERT, KAN, MCL, SMZ, STM, TET | AMP, KAN, STM, TET | A | O108:H35 | 10598 | |
| UPR-0060 | SRR10990264 | mdf(A) | MCL | -* | A | OXY72:H23 | 224 | |
| UPR-0061 | SRR10987144 | aph(3′)-Ia, aph(3″)-Ib, aph(6)-Id, blaCTX-M-1, blaTEM-1B, blaTEM-57, floR, mdf(A), sul2, tet(A), tet(B) | AMP, CHL, CIP, ERT, KAN, MCL, SMZ, STM, TET | AMP, KAN, STM, TET | B1 | O38:H26 | 10758 | |
| UPR-0063 | SRR10987120 | aadA2, dfrA12, floR, qnrB19, mdf(A), sul1, tet(A) | CHL, CIP, MCL, SMZ, STM, TET, TMP | STM, TET, STX | D | O27:H29 | 398 |
| Antibiotic Class | Phenotype Resistant | Genotype Resistant | Phenotype Sensitive | Genotype Sensitive | Correlation (%) |
|---|---|---|---|---|---|
| Aminoglycoside | 7 | 7 | 25 | 25 | 100 |
| Beta-lactam | 4 | 5 | 28 | 27 | 96 |
| Macrolide | -* | 32 | -* | 0 | -* |
| Phenicol | -* | 32 | -* | 0 | -* |
| Quinolone | 3 | 0 | 29 | 32 | 91 |
| Sulfonamide | 6 | 2 | 26 | 30 | 87 |
| Tetracycline | 6 | 6 | 26 | 26 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malavez, Y.; Nieves-Miranda, S.M.; Loperena Gonzalez, P.N.; Padin-Lopez, A.F.; Xiaoli, L.; Dudley, E.G. Exploring Antimicrobial Resistance Profiles of E. coli Isolates in Dairy Cattle: A Baseline Study across Dairy Farms with Varied Husbandry Practices in Puerto Rico. Microorganisms 2023, 11, 2879. https://doi.org/10.3390/microorganisms11122879
Malavez Y, Nieves-Miranda SM, Loperena Gonzalez PN, Padin-Lopez AF, Xiaoli L, Dudley EG. Exploring Antimicrobial Resistance Profiles of E. coli Isolates in Dairy Cattle: A Baseline Study across Dairy Farms with Varied Husbandry Practices in Puerto Rico. Microorganisms. 2023; 11(12):2879. https://doi.org/10.3390/microorganisms11122879
Chicago/Turabian StyleMalavez, Yadira, Sharon M. Nieves-Miranda, Paola N. Loperena Gonzalez, Adrian F. Padin-Lopez, Lingzi Xiaoli, and Edward G. Dudley. 2023. "Exploring Antimicrobial Resistance Profiles of E. coli Isolates in Dairy Cattle: A Baseline Study across Dairy Farms with Varied Husbandry Practices in Puerto Rico" Microorganisms 11, no. 12: 2879. https://doi.org/10.3390/microorganisms11122879