
Molecular Detection and Genotyping of Theileria spp. in Deer (Cervidae) in Korea
 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
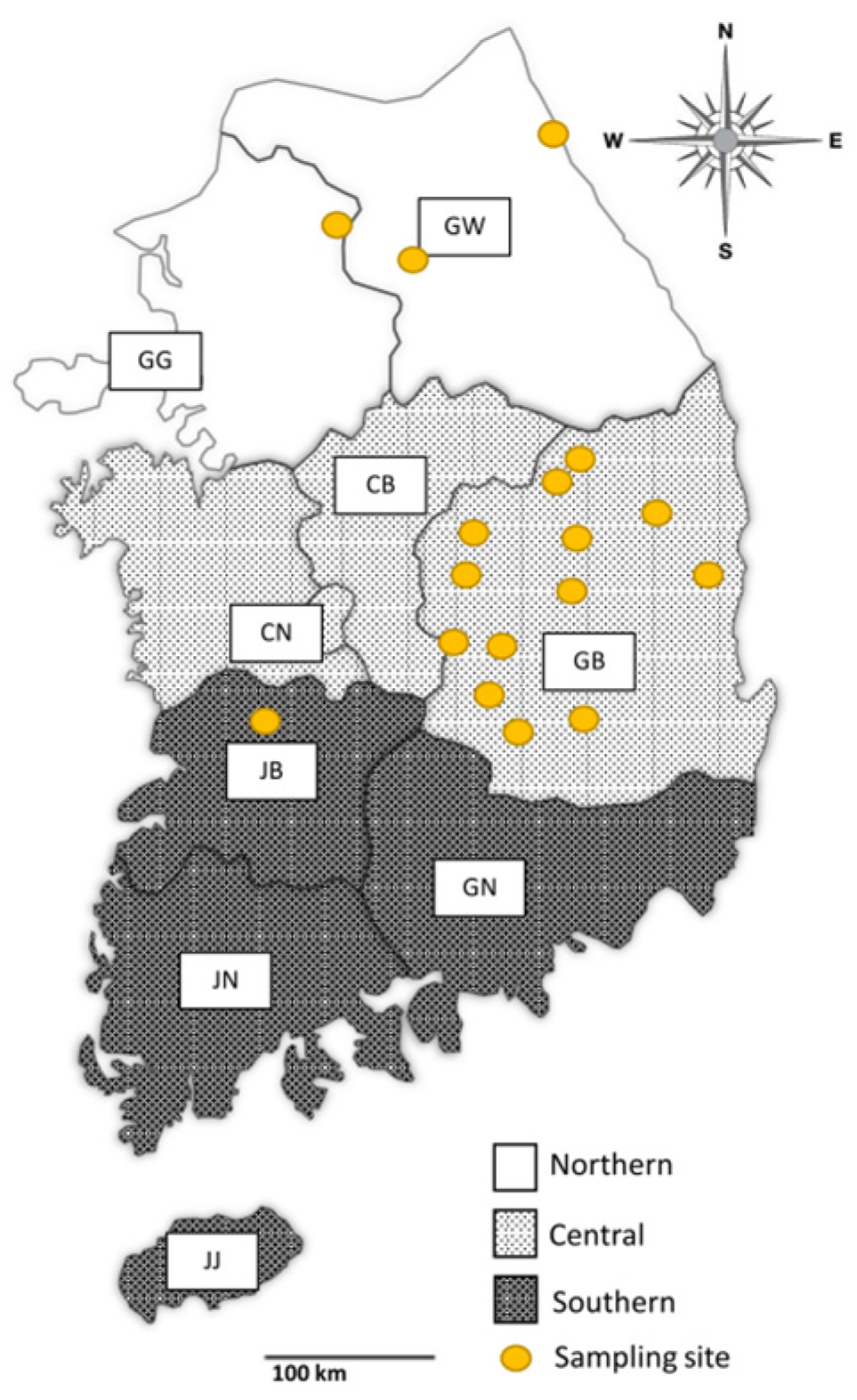
2.1. Blood Sample Collection
2.2. DNA Extraction and PCR Assay
2.3. Statistical and Phylogenetic Analysis
3. Results
3.1. Region-Wise Infection Rates of Theileria
3.2. Seasonal Variations in Theileria Infection Rates
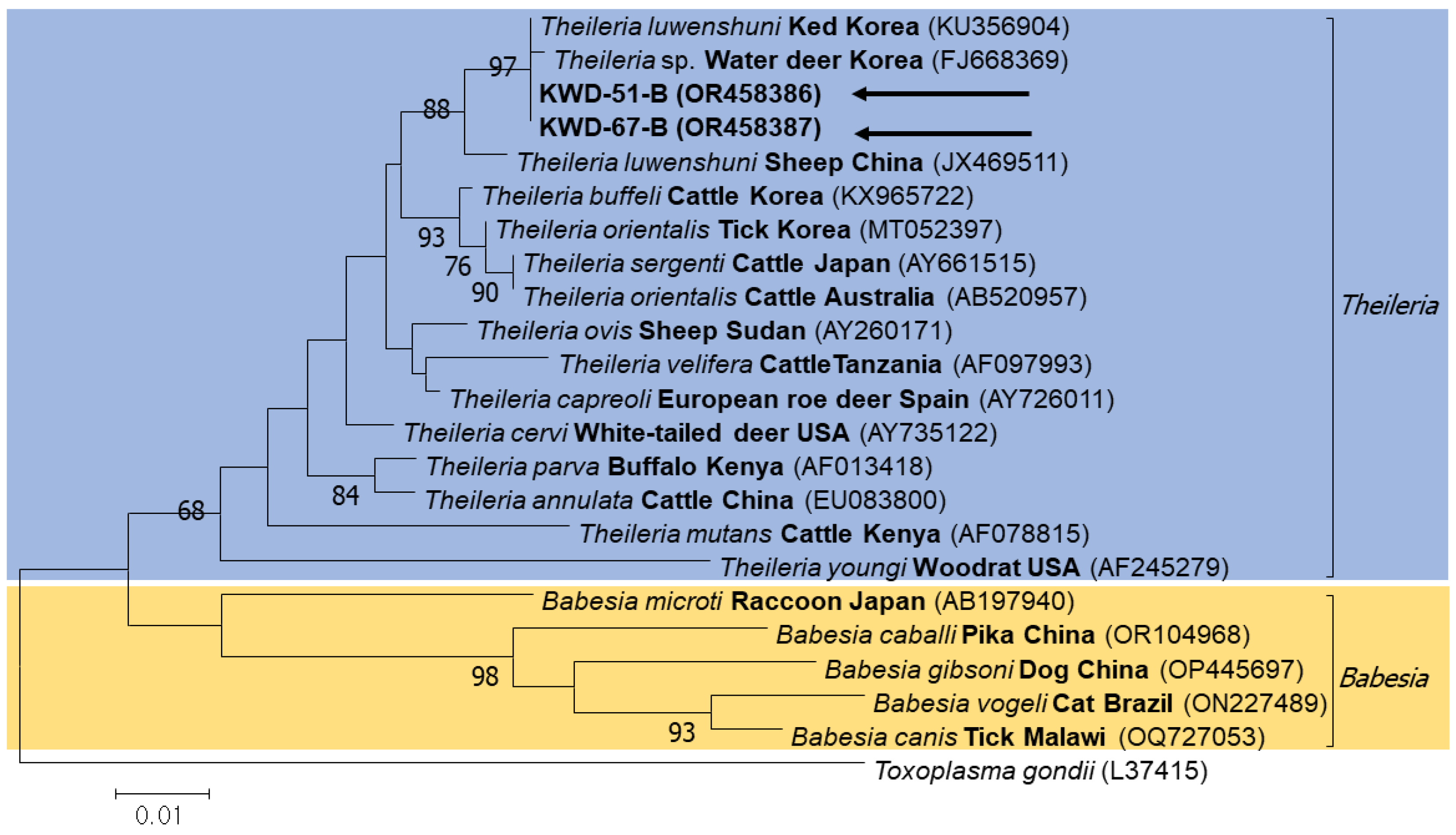
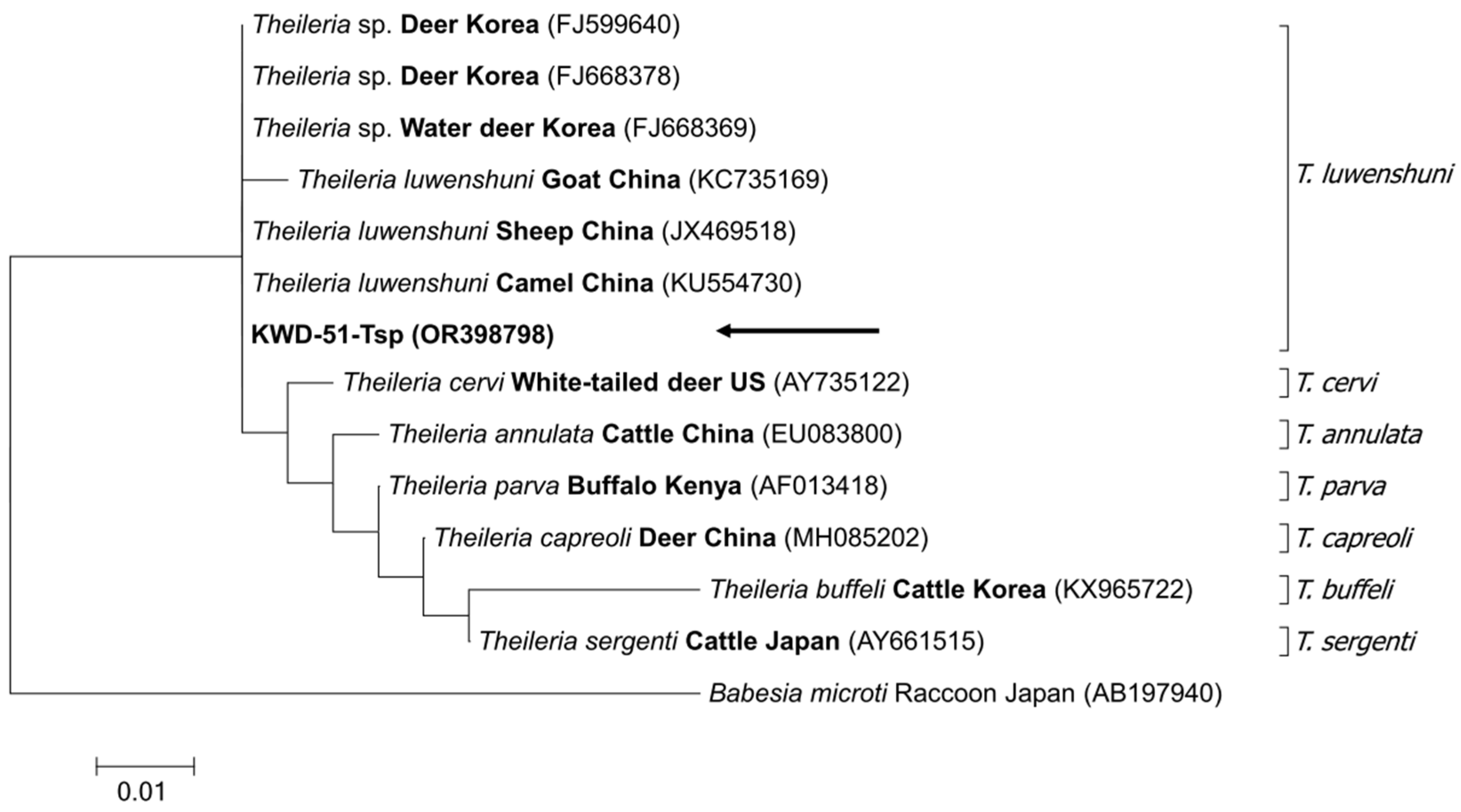
3.3. Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mans, B.J.; Pienaar, R.; Latif, A.A. A review of Theileria diagnostics and epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.O.; Niu, Q.; Yang, J.; Hassan, M.A.; Yu, P.; Guan, G.; Chen, Z.; Liu, G.; Luo, J.; Yin, H. Identification of 12 piroplasms infecting ten tick species in China using reverse line blot hybridization. J. Parasitol. 2017, 103, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Iori, A.; Gabrielli, S.; Calderini, P.; Moretti, A.; Pietrobelli, M.; Tampieri, M.P.; Galuppi, R.; Cancrini, G. Tick reservoirs for piroplasms in central and northern Italy. Vet. Parasitol. 2010, 170, 291–296. [Google Scholar] [CrossRef]
- Estrada-Peña, A. Ticks as vectors: Taxonomy, biology and ecology. Rev. Sci. Tech. 2015, 34, 53–65. [Google Scholar] [CrossRef]
- Anderson, J.F.; Magnarelli, L.A. Biology of ticks. Infect. Dis. Clin. N. Am. 2008, 22, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, A. A historical review of Babesia spp. associated with deer in Europe: Babesia divergens/Babesia divergens-like, Babesia capreoli, Babesia venatorum, Babesia cf. odocoilei. Vet. Parasitol. 2021, 294, 109433. [Google Scholar] [CrossRef]
- Uilenberg, G. Babesia–a historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Ahmed, J.S.; Luo, J.; Schnittger, L.; Seitzer, U.; Jongejan, F.; Yin, H. Phylogenetic position of small-ruminant infecting piroplasms. Ann. N. Y. Acad. Sci. 2006, 1081, 498–504. [Google Scholar] [CrossRef]
- Allsopp, M.T.; Allsopp, B.A. Molecular sequence evidence for the reclassification of some Babesia species. Ann. N. Y. Acad. Sci. 2006, 1081, 509–517. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Alkathiri, B.; Ahn, K.S.; Lee, H.S.; Cho, Y.S.; Youn, S.Y.; Seo, M.G.; Kwak, D.; Shin, S.S.; Lee, S.H. Molecular epidemiology of species in ticks and its potential threat to livestock in the Republic of Korea. Acta Trop. 2023, 238, 106780. [Google Scholar] [CrossRef] [PubMed]
- Bishop, R.; Musoke, A.; Morzaria, S.; Gardner, M.; Nene, V. Theileria: Intracellular protozoan parasites of wild and domestic ruminants transmitted by ixodid ticks. Parasitology 2004, 129 (Suppl. S1), S271–S283. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.T.; Pallas, E.; Gestal, J.J.; Guitián, F.J.; Olmeda, A.S.; Goethert, H.K.; Telford, S.R. Infection of dogs in north-west Spain with a Babesia microti-like agent. Vet. Rec. 2001, 149, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Kjemtrup, A.M.; Robinson, T.; Conrad, P.A. Description and epidemiology of Theileria youngi n. sp. from a northern Californian dusky-footed woodrat (Neotoma fuscipes) population. J. Parasitol. 2001, 87, 373–378. [Google Scholar] [CrossRef]
- Morrison, W.I. The aetiology, pathogenesis and control of theileriosis in domestic animals. Rev. Sci. Tech. 2015, 34, 599–611. [Google Scholar] [CrossRef]
- Seo, M.G.; Yun, S.H.; Choi, S.K.; Cho, G.J.; Park, Y.S.; Cho, K.H.; Kwon, O.D.; Kwak, D. Molecular and phylogenetic analysis of equine piroplasms in the Republic of Korea. Res. Vet. Sci. 2013, 94, 579–583. [Google Scholar] [CrossRef]
- Dobbelaere, D.A.; Küenzi, P. The strategies of the parasite: A new twist in host-pathogen interactions. Curr. Opin. Immunol. 2004, 16, 524–530. [Google Scholar] [CrossRef]
- Sivakumar, T.; Hayashida, K.; Sugimoto, C.; Yokoyama, N. Evolution and genetic diversity of Theileria. Infect. Genet. Evol. 2014, 27, 250–263. [Google Scholar] [CrossRef]
- Baneth, G.; Florin-Christensen, M.; Cardoso, L.; Schnittger, L. Reclassification of Theileria annae as Babesia vulpes sp. nov. Parasit. Vectors 2015, 8, 207. [Google Scholar] [CrossRef]
- Jacobson, L.S.; Clark, I.A. The pathophysiology of canine babesiosis: New approaches to an old puzzle. J. S. Afr. Vet. Assoc. 1994, 65, 134–145. [Google Scholar]
- Matjila, P.T.; Leisewitz, A.L.; Oosthuizen, M.C.; Jongejan, F.; Penzhorn, B.L. Detection of a Theileria species in dogs in South Africa. Vet. Parasitol. 2008, 157, 34–40. [Google Scholar] [CrossRef] [PubMed]
- D’Oliveira, C.; van der Weide, M.; Habela, M.A.; Jacquiet, P.; Jongejan, F. Detection of Theileria annulata in blood samples of carrier cattle by PCR. J. Clin. Microbiol. 1995, 33, 2665–2669. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, M.J.; Hong, Y.; van der Weide, M.; Qi, B.; Nijman, I.J.; Guangyuan, L.; Jongejan, F. Molecular characterisation of the Theileria buffeli/orientalis group. Int. J. Parasitol. 2000, 30, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Mhadhbi, M.; Naouach, A.; Boumiza, A.; Chaabani, M.F.; BenAbderazzak, S.; Darghouth, M.A. In vivo evidence for the resistance of Theileria annulata to buparvaquone. Vet. Parasitol. 2010, 169, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Gebrekidan, H.; Hailu, A.; Kassahun, A.; Rohoušová, I.; Maia, C.; Talmi-Frank, D.; Warburg, A.; Baneth, G. Theileria infection in domestic ruminants in northern Ethiopia. Vet. Parasitol. 2014, 200, 31–38. [Google Scholar] [CrossRef]
- Zanet, S.; Trisciuoglio, A.; Bottero, E.; de Mera, I.G.; Gortazar, C.; Carpignano, M.G.; Ferroglio, E. Piroplasmosis in wildlife: Babesia and Theileria affecting free-ranging ungulates and carnivores in the Italian Alps. Parasit. Vectors 2014, 7, 70. [Google Scholar] [CrossRef]
- Phipps, L.P.; Hernández-Triana, L.M.; Goharriz, H.; Welchman, D.; Johnson, N. Detection of Theileria luwenshuni in sheep from Great Britain. Parasit. Vectors 2016, 9, 203. [Google Scholar] [CrossRef]
- Almazán, C.; Scimeca, R.C.; Reichard, M.V.; Mosqueda, J. Babesiosis and Theileriosis in North America. Pathogens 2022, 11, 168. [Google Scholar] [CrossRef]
- Romero-Salas, D.; Solis-Cortés, M.; Zazueta-Islas, H.M.; Flores-Vásquez, F.; Cruz-Romero, A.; Aguilar-Domínguez, M.; Salguero-Romero, J.L.; de León, A.P.; Fernández-Figueroa, E.A.; Lammoglia-Villagómez, M.Á.; et al. Molecular detection of Theileria equi in horses from Veracruz, Mexico. Ticks Tick Borne Dis. 2021, 12, 101671. [Google Scholar] [CrossRef]
- Remesar, S.; Díaz, P.; Prieto, A.; Markina, F.; Díaz Cao, J.M.; López-Lorenzo, G.; Fernández, G.; López, C.M.; Panadero, R.; Díez-Baños, P.; et al. Prevalence and distribution of Babesia and Theileria species in roe deer from Spain. Int. J. Parasitol. Parasites Wildl. 2019, 9, 195–201. [Google Scholar] [CrossRef]
- Da Silveira, J.A.; Rabelo, E.M.; Ribeiro, M.F. Detection of Theileria and Babesia in brown brocket deer (Mazama gouazoubira) and marsh deer (Blastocerus dichotomus) in the state of Minas Gerais, Brazil. Vet. Parasitol. 2011, 177, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.; Gedye, K.; McFadden, A.; Pulford, D.; Heath, A.; Pomroy, W. Review of the New Zealand Theileria orientalis Ikeda type epidemic and epidemiological research since 2012. Pathogens 2021, 10, 1346. [Google Scholar] [CrossRef] [PubMed]
- McFadden, A.M.; Vink, D.; Pulford, D.J.; Lawrence, K.; Gias, E.; Heath, A.C.; McFadden, C.B.; Bingham, P. Monitoring an epidemic of Theileria-associated bovine anaemia (Ikeda) in cattle herds in New Zealand. Prev. Vet. Med. 2016, 125, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Storey-Lewis, B.; Mitrovic, A.; McParland, B. Molecular detection and characterisation of Babesia and Theileria in Australian hard ticks. Ticks Tick Borne Dis. 2018, 9, 471–478. [Google Scholar] [CrossRef]
- Kohli, S.; Atheya, U.K.; Thapliyal, A. Prevalence of theileriosis in cross-bred cattle: Its detection through blood smear examination and polymerase chain reaction in Dehradun district, Uttarakhand, India. Vet. World 2014, 7, 168–171. [Google Scholar] [CrossRef]
- Naz, S.; Maqbool, A.; Ahmed, S.; Ashraf, K.; Ahmed, N.; Saeed, K.; Latif, M.; Iqbal, J.; Ali, Z.; Shafi, K.; et al. Prevalence of theileriosis in small ruminants in Lahore-Pakistan. J. Vet. Anim. Sci. 2011, 2, 16–20. [Google Scholar]
- Bawm, S.; Kakisaka, K.; Thu, M.J.; Chel, H.M.; Oo, Y.M.N.; Soe, N.C.; Win, S.Y.; Htun, L.L.; Win, M.M.; Suzuki, H.; et al. First molecular detection of Theileria luwenshuni from goats in Myanmar. Parasitol. Res. 2018, 117, 3361–3364. [Google Scholar] [CrossRef]
- Lee, S.H.; Moumouni, P.F.A.; Galon, E.M.; Vudriko, P.; Liu, M.; Benedicto, B.; Tumwebaze, M.A.; Boldbaatar, D.; Umemiya-Shirafuji, R.; Fukumoto, S.; et al. Differential diagnosis and molecular characterization of Theileria spp. in sika deer (Cervus nippon) in Hokkaido, Japan. Parasitol. Int. 2019, 70, 23–26. [Google Scholar] [CrossRef]
- Liu, J.; Yang, J.; Guan, G.; Liu, A.; Wang, B.; Luo, J.; Yin, H. Molecular detection and identification of piroplasms in sika deer (Cervus nippon) from Jilin Province, China. Parasit. Vectors 2016, 9, 156. [Google Scholar] [CrossRef]
- Ge, Y.; Pan, W.; Yin, H. Prevalence of Theileria infections in goats and sheep in southeastern China. Vet. Parasitol. 2012, 186, 466–469. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.Y.; Liu, G.; Lyu, C.; Hu, Y.; An, Q.; Qiu, H.Y.; Zhao, Q.; Wang, C.R. Prevalence of Theileria in cattle in China: A systematic review and meta-analysis. Microb. Pathog. 2022, 162, 105369. [Google Scholar] [CrossRef] [PubMed]
- Jang, D.H. Epizootiological study of theileriasis in Korea. Korean J. Parasitol. 1974, 12, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Seong, G.; Han, Y.J.; Oh, S.S.; Chae, J.S.; Yu, D.H.; Park, J.; Park, B.K.; Yoo, J.G.; Choi, K.S. Detection of tick-borne pathogens in the Korean Water Deer (Hydropotes inermis argyropus) from Jeonbuk Province, Korea. Korean J. Parasitol. 2015, 53, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.U.; Park, Y.J.; Ryu, J.H.; Jang, D.H.; Hwang, S.; Cho, H.C.; Park, J.; Han, J.I.; Choi, K.S. Identification of zoonotic tick-borne pathogens from Korean water deer (Hydropotes inermis argyropus). Vector Borne Zoonotic Dis. 2020, 20, 745–754. [Google Scholar] [CrossRef]
- Han, J.I.; Jang, H.J.; Lee, S.J.; Na, K.J. High prevalence of Theileria sp. in wild Chinese water deer (Hydropotes inermis argyropus) in South Korea. Vet. Parasitol. 2009, 164, 311–314. [Google Scholar] [CrossRef]
- Choi, K.S.; Yu, D.H.; Chae, J.S.; Park, B.K.; Yoo, J.G.; Park, J. Seasonal changes in hemograms and Theileria orientalis infection rates among Holstein cattle pastured in the mountains in the Republic of Korea. Prev. Vet. Med. 2016, 127, 77–83. [Google Scholar] [CrossRef]
- Ahmed, J.S.; Glass, E.J.; Salih, D.A.; Seitzer, U. Innate immunity to tropical theileriosis. Innate Immun. 2008, 14, 5–12. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne disease: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef]
- Kwak, D.; Seo, M.G. Genetic diversity of bovine hemoprotozoa in South Korea. Pathogens 2020, 9, 768. [Google Scholar] [CrossRef]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Molecular detection and phylogenetic analysis of canine tick-borne pathogens from Korea. Ticks Tick Borne Dis. 2020, 11, 101357. [Google Scholar] [CrossRef]
- Han, Y.J.; Park, J.; Lee, Y.S.; Chae, J.S.; Yu, D.H.; Park, B.K.; Kim, H.C.; Choi, K.S. Molecular identification of selected tick-borne pathogens in wild deer and raccoon dogs from the Republic of Korea. Vet. Parasitol. Reg. Stud. Rep. 2017, 7, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, K.T.; Kwon, O.D.; Ock, Y.; Kim, T.; Choi, D.; Kwak, D. Novel Detection of Coxiella spp., Theileria luwenshuni, and T. ovis Endosymbionts in Deer Keds (Lipoptena fortisetosa). PLoS ONE 2016, 11, e0156727. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Diversity and genotypic analysis of tick-borne pathogens carried by ticks infesting horses in Korea. Med. Vet. Entomol. 2021, 35, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Lee, S.; Choi, C.; Kim, S.; Kang, C.; Lee, K.; Yun, Y. Investigation of Theileria sp. from ticks and roe deer (Capreolus pygargus) in Jeju Island. J. Vet. Clin. 2014, 31, 6–10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region | No. Tested | No. Positive (%) | p Value |
|---|---|---|---|
| Northern | 5 | 5 (100) | <0.001 |
| Central | 140 | 4 (2.9) | |
| Southern | 1 | 1 (100) | |
| Unknown | 14 | 3 (21.4) | |
| Total | 160 | 13 (8.1) |
| Season | No. Tested | No. Positive (%) | p Value |
|---|---|---|---|
| Spring | 49 | 9 (18.4) | 0.014 |
| Summer | 45 | 4 (8.9) | |
| Autumn | 26 | 0 | |
| Winter | 33 | 0 | |
| Unknown | 7 | 0 | |
| Total | 160 | 13 (8.1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, C.U.; Lee, H.; Seo, M.-G.; Lee, S.-H.; Kim, K.-T.; Nazim, K.; Song, J.-S.; Bae, D.H.; Rhee, M.H.; Kwon, O.-D.; et al. Molecular Detection and Genotyping of Theileria spp. in Deer (Cervidae) in Korea. Microorganisms 2023, 11, 2740. https://doi.org/10.3390/microorganisms11112740
Chung CU, Lee H, Seo M-G, Lee S-H, Kim K-T, Nazim K, Song J-S, Bae DH, Rhee MH, Kwon O-D, et al. Molecular Detection and Genotyping of Theileria spp. in Deer (Cervidae) in Korea. Microorganisms. 2023; 11(11):2740. https://doi.org/10.3390/microorganisms11112740
Chicago/Turabian StyleChung, Chang Uk, Haeseung Lee, Min-Goo Seo, Seung-Hun Lee, Kyoo-Tae Kim, Kaifa Nazim, Jung-Sun Song, Dong Hwa Bae, Man Hee Rhee, Oh-Deog Kwon, and et al. 2023. "Molecular Detection and Genotyping of Theileria spp. in Deer (Cervidae) in Korea" Microorganisms 11, no. 11: 2740. https://doi.org/10.3390/microorganisms11112740