
Reconstruction of the Steroid 1(2)-Dehydrogenation System from Nocardioides simplex VKM Ac-2033D in Mycolicibacterium Hosts
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganisms and Cultivation

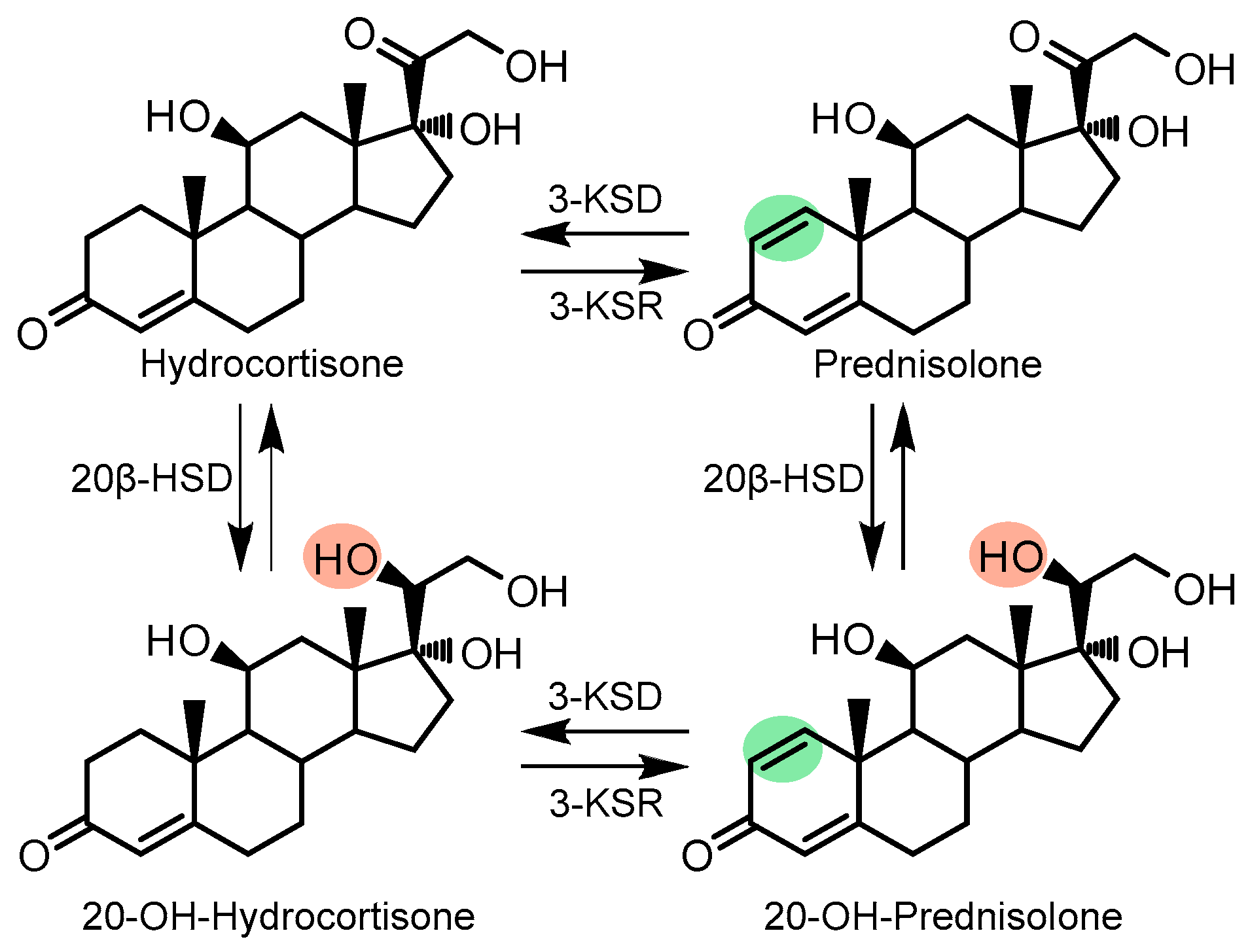{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Description | Source |
|---|---|---|
| E. coli DH5α | Strain for maintenance and amplification of plasmids | Thermo Fisher Scientific (USA) |
| Nocardioides simplex VKM Ac-2033D | Strain producing 1,2-dehydrosteroids, source of kstD2NS (KR76_27125) | All-Russian Collection of Microorganisms (VKM) |
| Mycolicibacterium neoaurum NRRL B-3805∆kstD | Sterol-transforming strain producing AD, kstD knockouted | Dr. M. Smith, University of York, York, UK [38]. |
| Mycolicibacterium smegmatis BD | M. smegmatis mc2 155, kshB and kstD knockouted | Karpov et al., 2022 [39] |
| Mycolicibacterium neoaurum NRRL B-3805∆kstD/pMVT61 | M. neoaurum NRRL B-3805∆kstD with pMVT61 plasmid | This study |
| Mycolicibacterium neoaurum NRRL B-3805∆kstD/pMami_k | M. neoaurum NRRL B-3805∆kstD with pMami_k plasmid | This study |
| Mycolicibacterium smegmatis BD/pMVT61 | M. smegmatis BD with pMVT61 plasmid | This study |
| Mycolicibacterium smegmatis BD/pMami_k | M. smegmatis BD with pMami_k plasmid | This study |
| Mycolicibacterium smegmatis BD/pMV261-N | M. smegmatis BD with pMV261-N plasmid | This study |
| Mycolicibacterium smegmatis BD/pMhsp_k | M. smegmatis BD with pMhsp_k plasmid | This study |
| pSMT3-M | E. coli—Mycobacterium high-copy shuttle vector, HygR, Phsp60, 5.7 kb | Carroll et al., 2010 [40] |
| pSMT3-MN | pSMT3-M with NdeI site introduced to the polylinker | This study |
| pMVT61 | E. coli—Mycobacterium low-copy shuttle vector, KmR, inducible acetamidase promoter (Pami), C-terminal His-tag, 8.0 kb | Karpov et al., 2022 [39] |
| pMV261 | E. coli—Mycobacterium low-copy shuttle vector, KmR, Phsp60, 4.4 kb | Stover et al., 1991 [41] |
| pMV261-N | pMV261 with NdeI site introduced to the polylinker | This study |
| pSM_k | pSMT3-MN with kstD2NS gene | This study |
| pMami_k | pMVT61 with kstD2NS gene | This study |
| pMhsp_k | pMV261-N with kstD2NS gene | This study |
2.3. Construction of Recombinant Plasmids
2.4. Total DNA Isolation
2.5. Isolation of Plasmid DNA
2.6. Electrotransformation of Mycolicibacterium Cells
2.7. SDS-PAGE Analysis of Proteins
2.8. Biotransformation of Steroids
2.9. Steroid Assays
2.10. Calculations
2.11. Statistics
3. Results
3.1. Construction of Recombinant Mycolicibacterium Strains
3.2. Biotransformation of Steroids by Recombinant Strains of M. neoaurum B-3805∆kstD
3.3. Biotransformation of Hydrocortisone by Recombinant Strains of M. smegmatis
3.4. Steroid 20-Carbonyl Group Reduction
3.5. Estimation of Minor Modifications and Destruction of Steroids
3.6. Reverse Activity of 1(2)-Hydrogenation of Prednisolone
3.7. The Effect of Temperature on Hydrocortisone Bioconversion by Mycolicibacteria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Donova, M.V.; Egorova, O.V. Microbial Steroid Transformations: Current State and Prospects. J. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef] [PubMed]
- Rohman, A.; Dijkstra, B.W. Application of Microbial 3-Ketosteroid Δ1-Dehydrogenases in Biotechnology. Biotechnol. Adv. 2021, 49, 107751. [Google Scholar] [CrossRef]
- Tong, W.-Y.; Dong, X. Microbial Biotransformation: Recent Developments on Steroid Drugs. Recent Pat. Biotechnol. 2009, 3, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Fokina, V.V.; Sukhodolskaya, G.V.; Shutov, A.A.; Nikolayeva, V.M.; Donova, M.V.; Savinova, T.S.; Lukashev, N.V.; Surovtsev, V.V. Microbiological Method of Producing 21-Acetoxypregna-1,4,9(11),16-Tetraene-3,20-Dione from 21-Acetoxypregna-4,9(11)16-Triene-3,20-Dione. Patent RU2480475, 27 July 2011. [Google Scholar]
- Spelling, T. Process for the Overexpression of Dehydrogenases. U.S. Patent US7416866B2, 26 August 2008. [Google Scholar]
- Marcos-Escribano, A.; Bermejo, F.A.; Bonde-Larsen, A.L.; Retuerto, J.I.; Sierra, I.H. 1,2-Dehydrogenation of Steroidal 6-Methylen Derivatives. Synthesis of Exemestane. Tetrahedron 2009, 65, 7587–7590. [Google Scholar] [CrossRef]
- Samuel, S.; Nguyen, T.; Choi, H.A. Pharmacologic Characteristics of Corticosteroids. J. Neurocrit. Care 2017, 10, 53–59. [Google Scholar] [CrossRef]
- Donova, M.V. Biokonversiya Steroidnyh Soedinenij Aktinobakteriyami [Bioconversion of Steroid Compounds by Actinobacteria]; ONTI of the Pushchinsky Scientific Center RAN: Pushchino, Russia, 2009; p. 196. [Google Scholar]
- Fokina, V.V.; Sukhodol’skaya, G.V.; Gulevskaya, S.A.; Gavrish, E.Y.; Evtushenko, L.I.; Donova, M.V. The 1(2)-Dehydrogenation of Steroid Substrates by Nocardioides simplex VKM Ac-2033D. Microbiology 2003, 72, 24–29. [Google Scholar] [CrossRef]
- Rohman, A.; Dijkstra, B.W. The Role and Mechanism of Microbial 3-Ketosteroid Δ1-Dehydrogenases in Steroid Breakdown. J. Steroid Biochem. Mol. Biol. 2019, 191, 105366. [Google Scholar] [CrossRef]
- Medentsev, A.G.; Arinbasarova, A.Y.; Koshcheyenko, K.A.; Akimenko, V.K. Svyaz’ 3-Ketosteroid-Δ1-degidrogenazy s Dykhatel’noy Tsep’yu Bakteriy Arthrobacter globiformis. [Relationship between 3-Ketosteroid-Δ1-Dehydrogenase and the Respiratory Chain of the Bacteria Arthrobacter globiformis]. Biochemistry 1983, 48, 1726–1732. [Google Scholar]
- Medentsev, A.G.; Arinbasarova, A.Y.; Koshcheyenko, K.A.; Akimenko, V.K.; Skryabin, G.K. Regulation of 3-Ketosteroid-1-En-Dehydrogenase Activity of Arthrobacter globiformis Cells by a Respiratory Chain. J. Steroid Biochem. Mol. Biol. 1985, 23, 365–368. [Google Scholar] [CrossRef]
- Wagner, B.; Atrat, P.G.; Clark-Curtiss, J.E.; Wagner, M. Localization of the Steroid 1-Dehydrogenase in Rhodococcus erythropolis IMET 7030 by Immunoelectron Microscopy. J. Basic Microbiol. 1992, 32, 65–71. [Google Scholar] [CrossRef]
- Wójcik, P.; Glanowski, M.; Wojtkiewicz, A.M.; Rohman, A.; Szaleniec, M. Universal Capability of 3-Ketosteroid Δ1-Dehydrogenases to Catalyze Δ1-Dehydrogenation of C17-Substituted Steroids. Microb. Cell Fact. 2021, 20, 119. [Google Scholar] [CrossRef] [PubMed]
- Shtratnikova, V.Y.; Schelkunov, M.I.; Fokina, V.V.; Pekov, Y.A.; Ivashina, T.; Donova, M.V. Genome-Wide Bioinformatics Analysis of Steroid Metabolism-Associated Genes in Nocardioides simplex VKM Ac-2033D. Curr. Genet. 2016, 62, 643–656. [Google Scholar] [CrossRef]
- Shtratnikova, V.Y.; Schelkunov, M.I.; Fokina, V.V.; Bragin, E.Y.; Shutov, A.A.; Donova, M.V. Different Genome-Wide Transcriptome Responses of Nocardioides simplex VKM Ac-2033D to Phytosterol and Cortisone 21-Acetate. BMC Biotechnol. 2021, 21, 7. [Google Scholar] [CrossRef] [PubMed]
- Yamané, T.; Nakatani, H.; Sada, E.; Omata, T.; Tanaka, A.; Fukui, S. Steroid Bioconversion in Water-Insoluble Organic Solvents: Δ1-Dehydrogenation by Free Microbial Cells and by Cells Entrapped in Hydrophilic or Lipophilic Gels: Steroid Conversion in Organic Solvents. Biotechnol. Bioeng. 1979, 21, 2133–2145. [Google Scholar] [CrossRef]
- Larsson, P.O.; Ohlson, S.; Mosbach, K. New Approach to Steroid Conversion Using Activated Immobilised Microorganisms. Nature 1976, 263, 796–797. [Google Scholar] [CrossRef]
- Arinbasarova, A.Y.; Medentsev, A.G.; Akimenko, V.K.; Koshcheyenko, K.A.; Skryabin, G.K. Redox Reactions in Hydrocortisone Transformation by Arthrobacter globiformis Cells. J. Steroid Biochem. 1985, 23, 307–312. [Google Scholar] [CrossRef]
- Ghosh, D.; Wawrzak, Z.; Weeks, C.M.; Duax, W.L.; Erman, M. The Refined Three-Dimensional Structure of 3α,20β-Hydroxysteroid Dehydrogenase and Possible Roles of the Residues Conserved in Short-Chain Dehydrogenases. Structure 1994, 2, 629–640. [Google Scholar] [CrossRef]
- Gerber, A.; Milhim, M.; Hartz, P.; Zapp, J.; Bernhardt, R. Genetic Engineering of Bacillus megaterium for High-Yield Production of the Major Teleost Progestogens 17α,20β-Di- and 17α,20β,21α-Trihydroxy-4-Pregnen-3-One. Metab. Eng. 2016, 36, 19–27. [Google Scholar] [CrossRef]
- Flegler, A.; Lipski, A. Engineered CRISPR/Cas9 System for Transcriptional Gene Silencing in Arthrobacter Species Indicates Bacterioruberin Is Indispensable for Growth at Low Temperatures. Curr. Microbiol. 2022, 79, 199. [Google Scholar] [CrossRef]
- Luo, J.-M.; Cui, H.-L.; Jia, H.-C.; Li, F.; Cheng, H.-J.; Shen, Y.-B.; Wang, M. Identification, Biological Characteristics, and Active Site Residues of 3-Ketosteroid Δ1-Dehydrogenase Homologues from Arthrobacter simplex. J. Agric. Food Chem. 2020, 68, 9496–9512. [Google Scholar] [CrossRef]
- Wojtkiewicz, A.M.; Wójcik, P.; Procner, M.; Flejszar, M.; Oszajca, M.; Hochołowski, M.; Tataruch, M.; Mrugała, B.; Janeczko, T.; Szaleniec, M. The Efficient Δ1-Dehydrogenation of a Wide Spectrum of 3-Ketosteroids in a Broad pH Range by 3-Ketosteroid Dehydrogenase from Sterolibacterium denitrificans. J. Steroid Biochem. Mol. Biol. 2020, 202, 105731. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, X.; Wang, Y.; Han, Y.; Sun, J.; Shi, J.; Zhang, B. Identification, Function, and Application of 3-Ketosteroid Δ1-Dehydrogenase Isozymes in Mycobacterium neoaurum DSM 1381 for the Production of Steroidic Synthons. Microb. Cell Fact. 2018, 17, 77. [Google Scholar] [CrossRef]
- Zhang, R.; Xu, X.; Cao, H.; Yuan, C.; Yuminaga, Y.; Zhao, S.; Shi, J.; Zhang, B. Purification, Characterization, and Application of a High Activity 3-Ketosteroid-Δ1-Dehydrogenase from Mycobacterium neoaurum DSM 1381. Appl. Microbiol. Biotechnol. 2019, 103, 6605–6616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ren, Y.; He, J.; Cheng, S.; Yuan, J.; Ge, F.; Li, W.; Zhang, Y.; Xie, G. Multiplicity of 3-Ketosteroid Δ1-Dehydrogenase Enzymes in Gordonia neofelifaecis NRRL B-59395 with Preferences for Different Steroids. Ann. Microbiol. 2015, 65, 1961–1971. [Google Scholar] [CrossRef]
- Wang, X.; Feng, J.; Zhang, D.; Wu, Q.; Zhu, D.; Ma, Y. Characterization of New Recombinant 3-Ketosteroid-Δ1-Dehydrogenases for the Biotransformation of Steroids. Microbiol. Biotechnol. 2017, 101, 6049–6060. [Google Scholar] [CrossRef]
- Mao, S.; Wang, J.-W.; Liu, F.; Zhu, Z.; Gao, D.; Guo, Q.; Xu, P.; Ma, Z.; Hou, Y.; Cheng, X.; et al. Engineering of 3-Ketosteroid-∆1-Dehydrogenase Based Site-Directed Saturation Mutagenesis for Efficient Biotransformation of Steroidal Substrates. Microb. Cell Fact. 2018, 17, 141. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, R.; Feng, J.; Wu, Q.; Zhu, D.; Ma, Y. A New 3-Ketosteroid-Δ1–Dehydrogenase with High Activity and Broad Substrate Scope for Efficient Transformation of Hydrocortisone at High Substrate Concentration. Microorganisms 2022, 10, 508. [Google Scholar] [CrossRef]
- Shao, M.; Sha, Z.; Zhang, X.; Rao, Z.; Xu, M.; Yang, T.; Xu, Z.; Yang, S. Efficient Androst-1,4-Diene-3,17-Dione Production by Co-Expressing 3-Ketosteroid-Δ1-Dehydrogenase and Catalase in Bacillus Subtilis. J. Appl. Microbiol. 2017, 122, 119–128. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, D.; Yang, T.; Xu, M.; Rao, Z. Over-Expression of Mycobacterium neoaurum 3-Ketosteroid-Δ1-Dehydrogenase in Corynebacterium crenatum for Efficient Bioconversion of 4-Androstene-3,17-Dione to Androst-1,4-Diene-3,17-Dione. Electron. J. Biotechnol. 2016, 24, 84–90. [Google Scholar] [CrossRef]
- Chen, M.-M.; Wang, F.-Q.; Lin, L.-C.; Yao, K.; Wei, D.-Z. Characterization and Application of Fusidane Antibiotic Biosynethsis Enzyme 3-Ketosteroid-∆1-Dehydrogenase in Steroid Transformation. Appl. Microbiol. Biotechnol. 2012, 96, 133–142. [Google Scholar] [CrossRef]
- Wei, W.; Fan, S.-Y.; Wang, F.-Q.; Wei, D.-Z. Accumulation of Androstadiene-Dione by Overexpression of Heterologous 3-Ketosteroid Δ1-Dehydrogenase in Mycobacterium neoaurum NwIB-01. World J. Microbiol. Biotechnol. 2014, 30, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tian, Y.; Wang, J.; Li, Y.; Wang, H.; Mao, S.; Liu, X.; Wang, C.; Bie, S.; Lu, F. Construction of Engineered Arthrobacter simplex with Improved Performance for Cortisone Acetate Biotransformation. Appl. Microbiol. Biotechnol. 2013, 97, 9503–9514. [Google Scholar] [CrossRef] [PubMed]
- Dovbnya, D.V.; Khomutov, S.M.; Kollerov, V.V.; Donova, M.V. Obtaining of 11α-Hydroxyandrost-4-Ene-3,17-Dione from Natural Sterols. In Microbial Steroids; Barredo, J.-L., Herráiz, I., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1645, pp. 259–269. ISBN 978-1-4939-7182-4. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Loraine, J.K.; Smith, M.C.M. Genetic Techniques for Manipulation of the Phytosterol Biotransformation Strain Mycobacterium neoaurum NRRL B-3805. In Microbial Steroids; Barredo, J.-L., Herráiz, I., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1645, pp. 93–108. ISBN 978-1-4939-7182-4. [Google Scholar] [CrossRef]
- Karpov, M.V.; Nikolaeva, V.M.; Fokina, V.V.; Shutov, A.A.; Kazantsev, A.V.; Strizhov, N.I.; Donova, M.V. Creation and Functional Analysis of Mycolicibacterium smegmatis Recombinant Strains Carrying the Bacillary Cytochromes CYP106A1 and CYP106A2 Genes. Appl. Biochem. Microbiol. 2022, 58, 947–957. [Google Scholar] [CrossRef]
- Carroll, P.; Schreuder, L.J.; Muwanguzi-Karugaba, J.; Wiles, S.; Robertson, B.D.; Ripoll, J.; Ward, T.H.; Bancroft, G.J.; Schaible, U.E.; Parish, T. Sensitive Detection of Gene Expression in Mycobacteria under Replicating and Non-Replicating Conditions Using Optimized Far-Red Reporters. PLoS ONE 2010, 5, e9823. [Google Scholar] [CrossRef]
- Stover, C.K.; de la Cruz, V.F.; Fuerst, T.R.; Burlein, J.E.; Benson, L.A.; Bennett, L.T.; Bansal, G.P.; Young, J.F.; Lee, M.H.; Hatfull, G.F.; et al. New Use of BCG for Recombinant Vaccines. Nature 1991, 351, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Vlahov, R.; Pramatarova, V.; Spassov, G.; Suchodolskaya, G.V.; Koshcheenko, K.A. Transformation of Microcrystalline Hydrocortisone by Free and Immobilized Cells of Arthrobacter simplex. Appl. Microbiol. Biotechnol. 1990, 33, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Smolders, A.J.J.; Pinheiro, H.M.; Noronha, P.; Cabral, J.M.S. Steroid Bioconversion in a Microemulsion System. Biotechnol. Bioeng. 1991, 38, 1210–1217. [Google Scholar] [CrossRef]
- Kollerov, V.V.; Lobastova, T.G.; Monti, D.; Deshcherevskaya, N.O.; Ferrandi, E.E.; Fronza, G.; Riva, S.; Donova, M.V. Deoxycholic Acid Transformations Catalyzed by Selected Filamentous Fungi. Steroids 2016, 107, 20–29. [Google Scholar] [CrossRef]
- Dovbnya, D.V.; Ivashina, T.V.; Khomutov, S.M.; Shutov, A.A.; Deshcherevskaya, N.O.; Donova, M.V. Obtaining of 24-Norchol-4-Ene-3,22-Dione from Phytosterol with Mutants of Mycolicibacterium neoaurum. In Microbial Steroids; Barreiro, C., Barredo, J.-L., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2023; Volume 2704, pp. 291–312. ISBN 978-1-07-163384-7. [Google Scholar] [CrossRef]
- Convention on the Elaboration of a European Pharmacopoeia; Council of Europe; European Department for the Quality of Medicines; Europarat; European Directorate for the Quality of Medicines & Healthcare; European Pharmacopoeia Commission. European Pharmacopoeia, 7th ed.; Publ. in Accordance with the Convention on the Elaboration of a European Pharmacopoeia; Council of Europe: Strasburg, France, 2010; Volume 2, pp. 2786–2787. ISBN 978-92-871-7222-8. [Google Scholar]
- Anilkumar, G.; Chauhan, M.M.; Ajitkumar, P. Cloning and Expression of the Gene Coding for FtsH Protease from Mycobacterium tuberculosis H37Rv. Gene 1998, 214, 7–11. [Google Scholar] [CrossRef]
- Al-Zarouni, M.; Dale, J.W. Expression of Foreign Genes in Mycobacterium bovis BCG Strains Using Different Promoters Reveals Instability of the hsp60 Promoter for Expression of Foreign Genes in Mycobacterium bovis BCG Strains. Tuberculosis 2002, 82, 283–291. [Google Scholar] [CrossRef]
- Sun, H.; Yang, J.; Song, H. Engineering Mycobacteria Artificial Promoters and Ribosomal Binding Sites for Enhanced Sterol Production. Biochem. Eng. J. 2020, 162, 107739. [Google Scholar] [CrossRef]
- Dovbnya, D.; Khomutov, S.; Nikolayeva, V.; Donova, M. CD-Medium Control of Microbial Sterol Sidechain Cleavage. In Proceedings of the Ninth International Symposium on Cyclodextrins, Santiago de Compostela, Spain, 31 May–3 June 1998; Springer: Dordrecht, The Netherlands, 1999; pp. 395–398. [Google Scholar]
- Shao, M.; Zhang, X.; Rao, Z.; Xu, M.; Yang, T.; Li, H.; Xu, Z. Enhanced Production of Androst-1,4-Diene-3,17-Dione by Mycobacterium neoaurum JC-12 Using Three-Stage Fermentation Strategy. PLoS ONE 2015, 10, e0137658. [Google Scholar] [CrossRef] [PubMed]
- Arinbasarova, A.Y.; Karpov, A.V.; Fokina, V.V.; Medentsev, A.G.; Koshcheyenko, K.A. Kinetic Characteristics of 1-En-Dehydrogenation of 6α-Methylhydrocortisone by Cells of Arthrobacter Globiformis 193. Enzym. Microb. Technol. 1996, 19, 501–506. [Google Scholar] [CrossRef]
- Fokina, V.V.; Karpov, A.V.; Sidorov, I.A.; Andrjushina, V.A.; Arinbasarova, A.Y. The Influence of β-Cyclodextrin on the Kinetics of 1-En-Dehydrogenation of 6α-Methylhydrocortisone by Arthrobacter globiformis Cells. Appl. Microbiol. Biotechnol. 1997, 47, 645–649. [Google Scholar] [CrossRef]
- Sukhodolskaya, G.V.; Donova, M.V.; Nikolaeva, V.M.; Koshcheyenko, K.A.; Dovbnya, D.V.; Khomutov, S.M.; Gulevskaya, S.A. Method of the Producing 1(2)-Dehydroderivatives of 4-Delta-3-Ketosteroids. Patent RU2156302, 20 September 2000. [Google Scholar]
- Goren, T.; Harni, M.; Rimonk, S.; Aharonowitz, Y. 1-Ene-Steroid Reductase of Mycobacterium SP. NRRL B-3805. J. Steroid Biochem. 1983, 19, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Tekucheva, D.N.; Nikolayeva, V.M.; Karpov, M.V.; Timakova, T.A.; Shutov, A.V.; Donova, M.V. Bioproduction of Testosterone from Phytosterol by Mycolicibacterium neoaurum Strains: “One-Pot”, Two Modes. Bioresour. Bioprocess. 2022, 9, 116. [Google Scholar] [CrossRef]
- Tekucheva, D.N.; Fokina, V.V.; Nikolaeva, V.M.; Shutov, A.A.; Karpov, M.V.; Donova, M.V. Cascade Biotransformation of Phytosterol to Testosterone by Mycolicibacterium neoaurum VKM Ac-1815D and Nocardioides simplex VKM Ac-2033D Strains. Microbiology 2022, 91, 303–312. [Google Scholar] [CrossRef]
- Etienne, G.; Laval, F.; Villeneuve, C.; Dinadayala, P.; Abouwarda, A.; Zerbib, D.; Galamba, A.; Daffé, M. The Cell Envelope Structure and Properties of Mycobacterium smegmatis mc2 155: Is There a Clue for the Unique Transformability of the Strain? Microbiology 2005, 151, 2075–2086. [Google Scholar] [CrossRef]
- Liu, J.; Nikaido, H. A Mutant of Mycobacterium smegmatis Defective in the Biosynthesis of Mycolic Acids Accumulates Meromycolates. Proc. Natl. Acad. Sci. USA 1999, 96, 4011–4016. [Google Scholar] [CrossRef]
- Xiong, L.-B.; Liu, H.-H.; Zhao, M.; Liu, Y.-J.; Song, L.; Xie, Z.-Y.; Xu, Y.-X.; Wang, F.-Q.; Wei, D.-Z. Enhancing the Bioconversion of Phytosterols to Steroidal Intermediates by the Deficiency of kasB in the Cell Wall Synthesis of Mycobacterium neoaurum. Microb. Cell Fact. 2020, 19, 80. [Google Scholar] [CrossRef]
- Daugelat, S.; Kowall, J.; Mattow, J.; Bumann, D.; Winter, R.; Hurwitz, R.; Kaufmann, S.H.E. The RD1 Proteins of Mycobacterium tuberculosis: Expression in Mycobacterium smegmatis and Biochemical Characterization. Microbes Infect. 2003, 5, 1082–1095. [Google Scholar] [CrossRef] [PubMed]
- Marsheck, W.J.; Kraychy, S.; Muir, R.D. Microbial Degradation of Sterols. Appl. Microbiol. 1972, 23, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Snapper, S.B.; Melton, R.E.; Mustafa, S.; Kieser, T.; Jacobs, W.R., Jr. Isolation and Characterization of Efficient Plasmid Transformation Mutants of Mycobacterium smegmatis. Mol. Microbiol. 1990, 4, 1911–1919. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ge, F.; Tan, Y.; Zhang, G.; Li, W. Genome-Wide Transcriptome Profiling of Mycobacterium smegmatis mc2 155 Cultivated in Minimal Media Supplemented with Cholesterol, Androstenedione or Glycerol. Int. J. Mol. Sci. 2016, 17, 689. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Zappaterra, F.; Summa, D.; Semeraro, B.; Fantin, G. Δ1-Dehydrogenation and C20-Reduction of Cortisone and Hydrocortisone Catalyzed by Rhodococcus Strains. Molecules 2020, 25, 2192. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Cabezón, L.; Galán, B.; García, J.L. Unravelling a New Catabolic Pathway of C-19 Steroids in Mycobacterium smegmatis: C-19 Steroids in M. smegmatis. Environ. Microbiol. 2018, 20, 1815–1827. [Google Scholar] [CrossRef]
- Wang, X.; Hua, C.; Xu, X.; Wei, D. Two-Step Bioprocess for Reducing Nucleus Degradation in Phytosterol Bioconversion by Mycobacterium neoaurum NwIB-R10hsd4A. Appl. Biochem. Biotechnol. 2019, 188, 138–146. [Google Scholar] [CrossRef] [PubMed]








| Strain | Cultivation and Bioconversion Conditions | Maximal Specific Steroid 1(2)-Dehydrogenase Activity, μmol/(h × g) (DCW) | Molar Yield, % (mol/mol) | Estimation of Steroid Destruction, % (mol/mol) | ||||
|---|---|---|---|---|---|---|---|---|
| 1(2)-Dehydro-steroids | Prednisolone | 20β-Hydroxy-steroids | ||||||
| Growth Duration before Addition of Hydrocortisone, h | t, °C | Bioconversion Duration, h | ||||||
| M. neoaurum B-3805∆kstD/pMami_k | 24 ** | 30 | 120 | 7.11 ± 1.01 | 19.81 ± 1.48 | 18.26 ± 1.12 | 5.48 ± 0.29 | 5.02 ± 0.40 |
| 24 ** | 35 | 120 | 0.31 ± 0.01 | 1.12 ± 0.032 | 1.12 ± 0.032 | 6.73 ± 0.44 | 8.41 ± 0.34 | |
| 36 ** | 30 | 120 | 9.75 ± 2.83 | 35.24 ± 4.1 | 32.74 ± 3.26 | 6.24 ± 0.76 | 5.57 ± 0.86 | |
| M. neoaurum B-3805∆kstD/pMVT61 | 36 ** | 30 | 120 | 0 | 0 | 0 | 6.11 ± 0.19 | 4.65 ± 0.75 |
| 24 ** | 35 | 120 | 0 | 0 | 0 | 7.19 ± 1.04 | 9.51 ± 0.82 | |
| M. smegmatis BD/pMami_k | 24 ** | 30 | 48 | 71.19 ± 2.37 | 82.38 ± 3.54 | 81.20 ± 2.71 | 1.34 ± 0.24 | 4.63 ± 0.92 |
| 24 ** | 35 | 48 | 108.01 ± 5.28 | 80.74 ± 2.66 | 78.77 ± 2.73 | 2.73 ± 0.36 | 6.23 ± 2.06 | |
| 36 ** | 30 | 48 | 102.82 ± 2.14 | 95.46 ± 1.73 | 94.14 ± 1.79 | 1.32 ± 0.21 | 2.74 ± 0.82 | |
| M. smegmatis BD/pMVT61 | 24 ** | 35 | 48 | 0.759 ± 0.023 | 0.64 ± 0.012 | 0.64 ± 0.012 | 4.02 ± 0.18 | 5.32 ± 0.89 |
| M. smegmatis BD/pMhsp_k | 24 | 30 | 48 | 93.26 ± 2.88 | 94.63 ± 1.15 | 93.42 ± 0.88 | 0.81 ± 0.09 | 2.16 ± 0.18 |
| 24 | 35 | 48 | 92.82 ± 1.91 | 88.56 ± 0.28 | 86.97 ± 0.28 | 1.67 ± 0.34 | 4.51 ± 0.27 | |
| 36 | 30 | 48 | 123.23 ± 3.67 | 96.73 ± 1.12 | 95.43 ± 1.72 | 0.71 ± 0.12 | 0.43 ± 0.03 | |
| M. smegmatis BD/pMV261-N | 24 | 35 | 48 | 0.719 ± 0.03 | 0.69 ± 0.026 | 0.69 ± 0.026 | 2.48 ± 0.05 | 6.69 ± 1.44 |
| 36 | 30 | 48 | 0.329 ± 0.01 | 0.58 ± 0.042 | 0.58 ± 0.042 | 2.55 ± 0.68 | 5.83 ± 1.61 | |
| N. simplex VKM Ac-2033D | 24 ** | 30 | 3 | 1922.1 ± 41.3 | 97.63 ± 1.39 | 77.27 ± 2.18 | 20.7 ± 1.03 | 0.46 ± 0.039 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fufaeva, S.R.; Dovbnya, D.V.; Ivashina, T.V.; Shutov, A.A.; Donova, M.V. Reconstruction of the Steroid 1(2)-Dehydrogenation System from Nocardioides simplex VKM Ac-2033D in Mycolicibacterium Hosts. Microorganisms 2023, 11, 2720. https://doi.org/10.3390/microorganisms11112720
Fufaeva SR, Dovbnya DV, Ivashina TV, Shutov AA, Donova MV. Reconstruction of the Steroid 1(2)-Dehydrogenation System from Nocardioides simplex VKM Ac-2033D in Mycolicibacterium Hosts. Microorganisms. 2023; 11(11):2720. https://doi.org/10.3390/microorganisms11112720
Chicago/Turabian StyleFufaeva, Svetlana R., Dmitry V. Dovbnya, Tanya V. Ivashina, Andrei A. Shutov, and Marina V. Donova. 2023. "Reconstruction of the Steroid 1(2)-Dehydrogenation System from Nocardioides simplex VKM Ac-2033D in Mycolicibacterium Hosts" Microorganisms 11, no. 11: 2720. https://doi.org/10.3390/microorganisms11112720