
Aflatoxins Contamination in Feed Commodities: From Occurrence and Toxicity to Recent Advances in Analytical Methods and Detoxification

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Ecology of Aspergillus Section Flavi
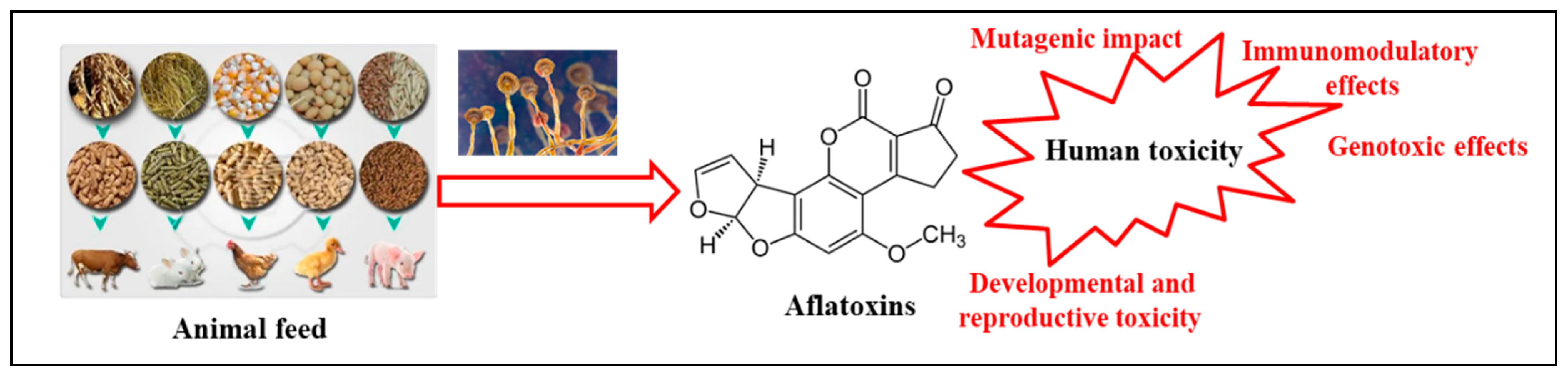
3. Toxicity of AFs
3.1. Genotoxic Effects
3.2. Mutagenic Impact
3.3. Developmental and Reproductive Toxicity and Immunomodulatory Effects
3.4. Gut Microbiota
3.5. Liver Damage
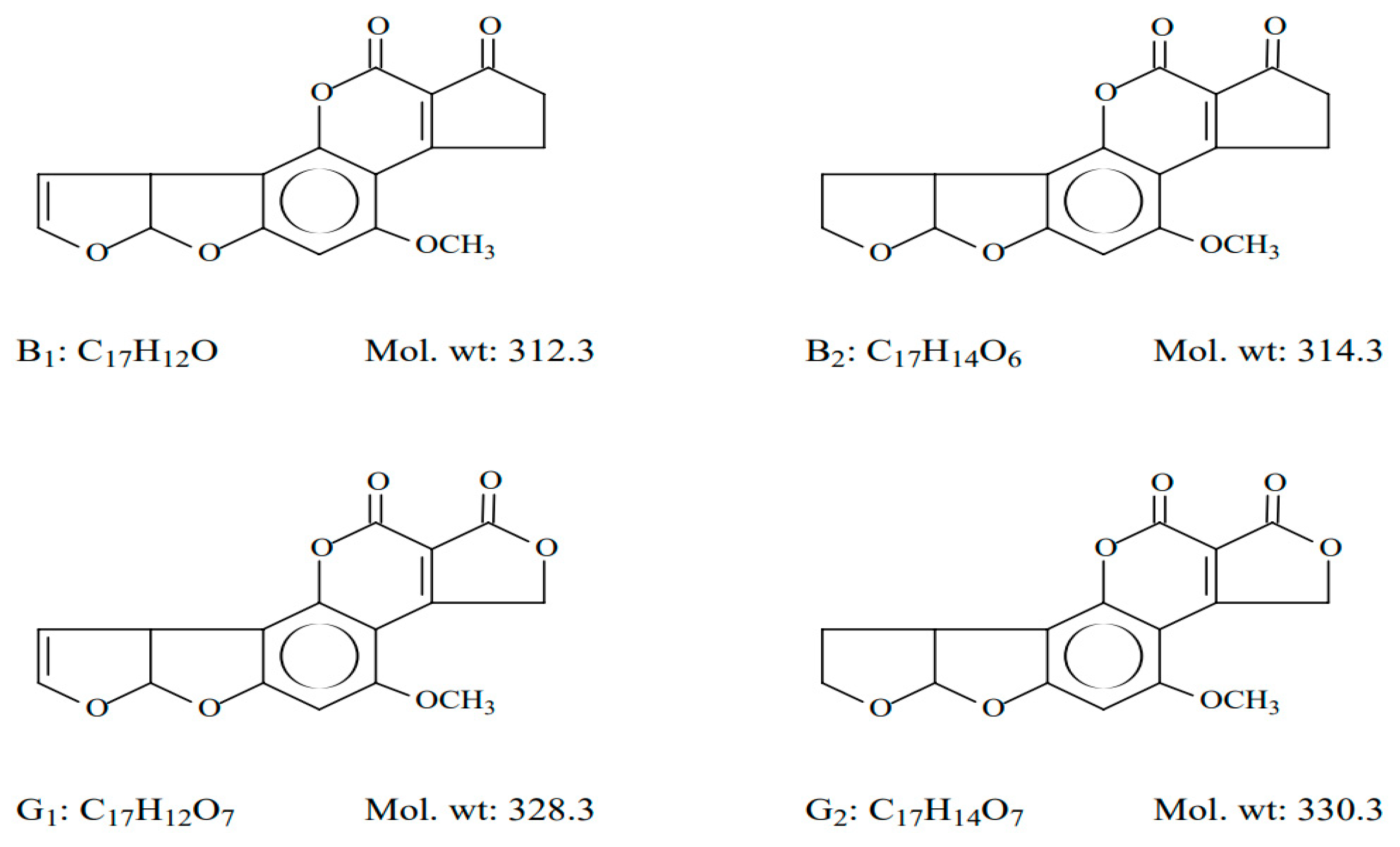
4. AFs Occurrence in Food/Feed Stuff
5. Chromatographic Technologies for AF Detection in Feeds
6. Prevention and Detoxification of AFs in Different Feed Matrices
6.1. Prevention of AFs in Feed Matrices
6.2. Post-Harvest Intervention Strategies
6.2.1. Physical Treatments
6.2.2. Chemical Agents
6.2.3. Biological Methods

{kind=link}
{kind=link}
| Rationale | Advantages | Treatment | Conditions (Measures/Reagents) | Pre-Harvest | Storage | Processing/Mixing | Feeding | References |
|---|---|---|---|---|---|---|---|---|
| Agricultural Prevention | ||||||||
| Interventions that prevent/deplete fungi infection or inhibit AFs production. | Prevention Low cost High durability | GAPs | tillage—farming planning crop rotation fertilizers biostimulants | × | [179] | |||
| Pesticides | azoles boscalid azoxystrobin cyprodinil fludioxonil | × | × | [185] [186] | ||||
| Transgenic crops | GMO RNA interferences CRISPR | × | [152,153,154,155] | |||||
| Atoxigenic A. spp strains | × | [155,156,157,158] | ||||||
| Bioremediation of fields | degradation transformation | × | [170] | |||||
| Physical Methods | ||||||||
| Interventions that permit to remove infected/contaminated commodities or kill/inactivate fungi and spores | Standardized protocol High efficacy, No harmful residues produced | Irradiation | gamma radiation UV lights X-rays Electrons | × | × | × | [188] [189] [183] | |
| Drying/Thermal treatment | microwave conventional heating | × | × | [181] | ||||
| Separation/sorting | manual sieves washing light-interaction-based devices | × | × | [179] [180] | ||||
| Chemical Methods | ||||||||
| Segregation, deactivation and degradation of AFs | Low cost | Ammonia | gaseous or liquid ammonia (usually 1.5–2%) alone for long time (until 15 days) or combined with other techniques | × | × | × | [36] | |
| Ozonisation | ozone 1.1–90 mg/l for a variable time (5 min–96 h) | × | × | × | [186] [187] | |||
| Alkalinization | potassium or sodium hydroxide ammonium carbonate | × | × | [189] | ||||
| Acidification | lactic acid propionic acid hydrochloric acid acetic acid citric acid phosphoric acid tartaric acid sorbic acid | × | × | [181] | ||||
| Nixtamalization | Cooking heat treatment+ alkaline solution (NaHCO3 and Ca(OH)2) + H2O2 | × | × | [189] [189] | ||||
| Active substances | turmeric powder curcumin resveratrol lycopene chitosan | × | × | × | × | [190] | ||
| Feed additives | acids antioxidants polymers sodium bisulfite | × | × | × | [116] | |||
| Adsorbents | clays activated charcoal bentonite montmorillonite, zeolite, hydrated sodium calcium aluminosilicate, kaolin, illite | × | × | × | × | [189] [190] | ||
| Biological Methods | ||||||||
| Inoculation of microbial strains in feed substrates. | Biodegradability Biosafety effectiveness regenerability | Bacteria | Bacillus spp. Lactobacillus spp. | × | × | × | [187] [187] | |
| Yeast and Fungi | Saccharomyces cerevisiae Rhizopus oligosporus Zygosaccharomyces rouxii | × | × | × | [189] [188] | |||
| Microorganism exudates/supernatants | Lactobacillus spp. | × | × | × | [190] | |||
| Enzymes | laccase manganese peroxidase oxidase | × | × | × | × | [188] | ||
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Garcia, S.N.; Osburn, B.I.; Jay-Russell, M.T. One Health for Food Safety, Food Security, and Sustainable Food Production. Front. Sustain. Food Syst. 2020, 4, 1. [Google Scholar] [CrossRef]
- FAO. Quality Assurance for Animal Feed Analysis Laboratories; FAO Animal Production and Health Manual No. 14.; FAO: Rome, Italy, 2011. [Google Scholar]
- OIE Animal Production Food Safety Working Group. Guide to good farming practices for animal production food safety. Rev. Sci. Tech. 2006, 25, 823–836. [Google Scholar]
- FAO; IFIF. Good Practices for the Feed Sector—Implementing the Codex Alimentarius Code of Practice on Good Animal Feeding; FAO Animal Production and Health Manual No. 24. Rome. 2020. Available online: https://www.fao.org/documents/card/en/c/cb1761en (accessed on 29 August 2023).
- European Food Safety Authority. EFSA CONTAM Panel, 2020. Scientific opinion—Risk assessment of aflatoxins in food. EFSA J. 2020, 18, 6040. [Google Scholar]
- International Agency for Research on Cancer (IARC). IARC Monographs on the Evaluation of the Carcinogenic Risks to Humans: Some Traditional Herbal Medicines, some Mycotoxins, Naphthalene and Styrene; IARC: Lyon, France, 2002; Volume 82, pp. 275–300. [Google Scholar]
- Smaoui, S.; Agriopoulou, S.; D’Amore, T.; Tavares, L.; Mousavi Khaneghah, A. The control of Fusarium growth and decontamination of produced mycotoxins by lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Sun, J.; Tan, Y.; Xiong, L.; Peng, B.; Peng, G.; Bai, X. Comparison of the health risks associated with exposure to toxic metals and metalloids following consumption of freshwater catches in China. Qual. Assur. Saf. Crops Foods 2022, 14, 1–12. [Google Scholar] [CrossRef]
- Aranega, J.P.; Oliveira, C.A. Occurrence of mycotoxins in pastures: A systematic review. Qual. Assur. Saf. Crops Foods 2022, 14, 135–144. [Google Scholar] [CrossRef]
- United Nations, Department of Economic and Social Affairs Sustainable Development. Transforming Our World: The 2030 Agenda for Sustainable Development; A/RES/70/1; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Thakaew, R.; Chaiklangmuang, S. Aflatoxin B1 elimination in low-grade maize by co-influence of heat and chemical treatment. Qual. Assur. Saf. Crops Foods 2023, 15, 55–67. [Google Scholar] [CrossRef]
- Basso, A.B.G.; Ali, S.; Corassin, C.H.; Rosim, R.E.; de Oliveria, C.A.F. Individual and combined decontamination effect of fermentation and ultrasound on aflatoxin B1 in wheat-based doughs: A preliminary study. Qual. Assur. Saf. Crops Foods 2022, 14, 96–103. [Google Scholar] [CrossRef]
- Norlia, M.; Jinap, S.; Nor-Khaizura, M.A.R.; Radu, S.; Samsudin, N.I.P.; Azri, F.A. Aspergillus section Flavi and aflatoxins: Occurrence, detection, and identification in raw peanuts and peanut-based products along the supply chain. Front. Microbiol. 2019, 10, 2602. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Novßkovß, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Horn, B.W.; Moore, G.G.; Carbone, I. Sexual reproduction in Aspergillus flavus. Mycologia 2009, 101, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W.; Ramirez-Prado, J.H.; Carbone, I. The sexual state of Aspergillus parasiticus. Mycologia 2009, 101, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W.; Sorensen, R.B.; Lamb, M.C.; Sobolev, V.S.; Olarte, R.A.; Worthington, C.J.; Carbone, I. Sexual reproduction in Aspergillus flavus sclerotia naturally produced in corn. Phytopathology 2014, 104, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W.; Gell, R.M.; Singh, R.; Sorensen, R.B.; Carbone, I. Sexual reproduction in Aspergillus flavus sclerotia: Acquisition of novel alleles from soil populations and uniparental mitochondrial inheritance. PLoS ONE 2016, 11, e0146169. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Heydt, M.; Rüfer, C.E.; Abdel-Hadi, A.; Magan, N.; Geisen, R. The production of aflatoxin B1 or G1 by Aspergillus parasiticus at various combinations of temperature and water activity is related to the ratio of aflS to afl R expression. Mycotoxin Res. 2010, 26, 241–246. [Google Scholar] [CrossRef]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. R. Soc. Interface 2012, 9, 757–767. [Google Scholar] [CrossRef]
- Bernaldez, V.; Cordoba, J.J.; Magan, N.; Peromingo, B.; Rodriguez, A. The influence of ecophysiological factors on growth, aflR gene expression and aflatoxin B1 production by a type strain of Aspergillus flavus. LWT-Food Sci. Technol 2017, 83, 283–291. [Google Scholar] [CrossRef]
- Lahouar, A.; Marin, S.; Crespo-Sempere, A.; Saïd, S.; Sanchis, V. Effects of temperature, water activity and incubation time on fungal growth and aflatoxin B1 production by toxinogenic Aspergillus flavus isolates on sorghum seeds. Rev. Argent. Microbiol. 2016, 48, 78–85. [Google Scholar] [CrossRef]
- Mousa, W.; Ghazali, F.M.; Jinap, S.; Ghazali, H.M.; Radu, S. Modelling the effect of water activity and temperature on growth rate and aflatoxin production by two isolates of Aspergillus flavus on paddy. J. Appl. Microbiol. 2011, 111, 1262–1274. [Google Scholar] [CrossRef]
- Pottenger, L.H.; Andrews, L.S.; Bachman, A.N.; Boogaard, P.J.; Cadet, J.; Embry, M.R.; Farmer, P.B.; Himmelstein, M.W.; Jarabek, A.M.; Martin, E.A.; et al. An organizational approach for the assessment of DNA adduct data in risk assessment: Case studies for aflatoxin B-1, tamoxifen and vinyl chloride. Crit. Rev. Toxicol. 2014, 44, 348–391. [Google Scholar] [CrossRef]
- Fasullo, M.; Freedland, J.; St, J.N.; Cera, C.; Hartog, M.; Ding, X.; Fasullo, M.; Egner, P. An in vitro system for measuring genotoxicity mediated by human CYP3A4 in Saccharomyces cerevisiae. Environ. Mol. Mutagen. 2017, 58, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Theumer, M.G.; Henneb, Y.; Khoury, L.; Snini, S.P.; Tadrist, S.; Canlet, C.; Puel, O.; Oswald, I.P.; Audebert, M. Genotoxicity of aflatoxins and their precursors in human cells. Toxicol. Lett. 2018, 287, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Paget, V.; Sichel, F.; Garon, D.; Lechevrel, M. Aflatoxin B1-induced TP53 mutational pattern in normal human cells using the FASAY (Functional Analysis of Separated Alleles in Yeast). Mutat. Res. 2008, 656, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Paget, V.; Lechevrel, M.; Andre, V.; Le Goff, J.; Pottier, D.; Billet, S.; Garcon, G.; Shirali, P.; Sichel, F. Benzo[a]pyrene, aflatoxin B1 and acetaldehyde mutational patterns in TP53 gene using a functional assay: Relevance to human cancer aetiology. PLoS ONE 2012, 7, e30921. [Google Scholar] [CrossRef]
- Corcuera, L.-A.; Vettorazzi, A.; Arbillaga, L.; Perez, N.; Gil, A.G.; Azqueta, A.; Gonzalez-Penas, E.; Garcia-Jalon, J.A.; Lopez de Cerain, A. Genotoxicity of Aflatoxin B1 and Ochratoxin A after simultaneous application of the in vivo micronucleus and comet assay. Food Chem. Toxicol. 2015, 76, 116–124. [Google Scholar] [CrossRef]
- Chawanthayatham, S.; Thiantanawat, A.; Egner, P.A.; Groopman, J.D.; Wogan, G.N.; Croy, R.G.; Essigmann, J.M. Prenatal exposure of mice to the human liver carcinogen aflatoxin B-1 reveals a critical window of susceptibility to genetic change. Int. J. Cancer 2015, 136, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Sriwattanapong, K.; Slocum, S.L.; Chawanthayatham, S.; Fedeles, B.I.; Egner, P.A.; Groopman, J.D.; Satayavivad, J.; Croy, R.G.; Essigmann, J.M. Editor’s highlight: Pregnancy alters aflatoxin B1 metabolism and increases DNA damage in mouse liver. Toxicol. Sci. 2017, 160, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Wattanawaraporn, R.; Woo, L.L.; Belanger, C.; Chang, S.-C.; Adams, J.E.; Trudel, L.J.; Bouhenguel, J.T.; Egner, P.A.; Groopman, J.D.; Croy, R.G.; et al. A single neonatal exposure to aflatoxin b1 induces prolonged genetic damage in two loci of mouse liver. Toxicol. Sci. 2012, 128, 326–333. [Google Scholar] [CrossRef]
- Shupe, T.; Sell, S. Low hepatic glutathione S-transferase and increased hepatic DNA adduction contribute to increased tumorigenicity of aflatoxin B1 in newborn and partially hepatectomized mice. Toxicol. Lett. 2004, 148, 1–9. [Google Scholar] [CrossRef]
- Gouas, D.A.; Shi, H.; Hautefeuille, A.H.; Ortiz-Cuaran, S.L.; Legros, P.C.; Szymanska, K.J.; Galy, O.; Egevad, L.A.; Abedi-Ardekani, B.; Wiman, K.G.; et al. Effects of the TP53 p.R249S mutant on proliferation and clonogenic properties in human hepatocellular carcinoma cell lines: Interaction with hepatitis B virus X protein. Carcinogenesis 2010, 31, 1475–1482. [Google Scholar] [CrossRef]
- Ortiz-Cuaran, S.; Hainaut, P. Molecular signatures of environmental mutagens in hepatocellular carcinoma. Genes Environ. 2011, 33, 141–148. [Google Scholar] [CrossRef]
- Huang, M.N.; Yu, W.; Teoh, W.W.; Ardin, M.; Jusakul, A.; Ng, A.W.T.; Boot, A.; Abedi-Ardekani, B.; Villar, S.; Myint, S.S.; et al. Genomescale mutational signatures of aflatoxin in cells, mice, and human tumors. Genome Res. 2017, 27, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; He, H.; Zang, M.; Wu, Q.; Zhao, H.; Lu, L.L.; Ma, P.; Zheng, H.; Wang, N.; Zhang, Y.; et al. Genetic features of aflatoxin-associated hepatocellular carcinoma. Gastroenterology 2017, 153, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Wogan, G.N.; Paglialunga, S.; Newberne, P.M. Carcinogenic effects of low dietary levels of aflatoxin B1 in rats. Food Cosmet. Toxicol. 1974, 12, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.H.; Greenblatt, M.; Lijinsky, W. Carcinogenesis in rats by aflatoxins B1, G1, and B2. Cancer Res. 1969, 29, 2206–2211. [Google Scholar]
- Cullen, J.M.; Ruebner, B.H.; Hsieh, L.S.; Hyde, D.M.; Hsieh, D. Carcinogenicity of dietary aflatoxin M in male Fischer rats compared to aflatoxin B1. Cancer Res. 1987, 47, 1913–1917. [Google Scholar]
- Halver, J.E. Aflatoxicosis and trout hepatoma. Bull.-Off. Int. Épizoot. 1968, 69, 1249–1278. [Google Scholar]
- Williams, D.E. The rainbow trout liver cancer model: Response to environmental chemicals and studies on promotion and chemoprevention. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 121–127. [Google Scholar] [CrossRef]
- Williams, D.E.; Orner, G.; Willard, K.D.; Tilton, S.; Hendricks, J.D.; Pereira, C.; Benninghoff, A.D.; Bailey, G.S. Rainbow trout (Oncorhynchus mykiss) and ultra-low dose cancer studies. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149C, 175–181. [Google Scholar] [CrossRef]
- Zuberi, Z.; Eeza, M.N.H.; Matysik, J.; Berry, J.P.; Alia, A. NMR-based metabolic profiles of intact zebrafish embryos exposed to aflatoxin B1 recapitulates hepatotoxicity and supports possible neurotoxicity. Toxins 2019, 11, 258. [Google Scholar] [CrossRef]
- Tanaka, T.; Mizukami, S.; Hasegawa-Baba, Y.; Onda, N.; Sugita-Konishi, Y.; Yoshida, T.; Shibutani, M. Developmental exposure of aflatoxin B-1 reversibly affects hippocampal neurogenesis targeting late-stage neural progenitor cells through suppression of cholinergic signaling in rats. Toxicology 2015, 336, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, W.; Wang, C.C.; Leung, L.K. Exposure to aflatoxin B1 in late gestation alters protein kinase C and apoptotic protein expression in murine placenta. Reprod. Toxicol. 2016, 61, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Hasanzadeh, S.; Amani, S. Aflatoxin B1 effects on ovarian follicular growth and atresia in the rat. Comp. Clin. Path. 2013, 22, 563–572. [Google Scholar] [CrossRef]
- Hasanzadeh, S.; Rezazadeh, L. Effects of aflatoxin B1 on the growth processes of spermatogenic cell series in adult male rats. Comp. Clin. Path. 2013, 22, 555–562. [Google Scholar] [CrossRef]
- Hasanzadeh, S.; Hosseini, E.; Rezazadeh, L. Effects of aflatoxin B-1 on profiles of gonadotropic (FSH and LH), steroid (testosterone and 17 beta-estradiol) and prolactin hormones in adult male rat. Iran. J. Vet. Res. 2011, 12, 332–336. [Google Scholar]
- Mohammadi, T.; Pousty, I.; Gilanpour, H. The effect of aflatoxin b1 on the sperm characteristics, sperm dna damage, fertility potential and early embryonic development in NMRI mice. Adv. Environ. Biol. 2014, 8, 1186–1191. [Google Scholar]
- Komsky-Elbaz, A.; Saktsier, M.; Roth, Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology 2018, 393, 42–50. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Q.-C.; Han, J.; Xiong, B.; Sun, S.-C. Aflatoxin B1 is toxic to porcine oocyte maturation. Mutagenesis 2015, 30, 527–535. [Google Scholar] [CrossRef]
- Sun, Y.; Su, J.; Liu, Z.; Liu, D.; Gan, F.; Chen, X.; Huang, K. Aflatoxin B1 promotes influenza replication and increases virus related lung damage via activation of TLR4 signaling. Front. Immunol. 2018, 9, 2297. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Liu, D.; Chen, J.; Gan, F.; Huang, K. Low-level aflatoxin B1 promotes influenza infection and modulates a switch in macrophage polarization from M1 to M2. Cell. Physiol. Biochem. 2018, 49, 1110–1126. [Google Scholar] [CrossRef]
- Meissonnier, G.; Marin, D.; Galtier, P.; Bertin, G.; Taranu, I.; Oswald, I.; Mengheri, E. Modulation of the immune response by a group of fungal food contaminant, the aflatoxins. In Nutrition and Immunity; Mengheri, E., Roselli, M., Britti, M.S., Finamore, A., Eds.; Research Signpost: Kerala, India, 2006; pp. 147–166. [Google Scholar]
- Mehrzad, J.; Bahari, A.; Bassami, M.R.; Mahmoudi, M.; Dehghani, H. Data on environmentally relevant level of aflatoxin B1-induced human dendritic cells’ functional alteration. Data Br. 2018, 18, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Mehrzad, J.; Hosseinkhani, S.; Malvandi, A.M. Human microglial cells undergo proapoptotic induction and inflammatory activation upon in vitro exposure to a naturally occurring level of aflatoxin B1. NeuroImmunomodulation 2018, 25, 176–183. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Opinion of the scientific panel on contaminants in the food chain (CONTAM) related to the potential increase of consumer health risk by a possible increase of the existing maximum levels for aflatoxins in almonds, hazelnuts and pistachios and derived products. EFSA J. 2007, 5, 446. [Google Scholar]
- Bondy, G.S. Immunological toxicity of mycotoxins. Stewart Postharvest Rev. 2008, 4, 1–6. [Google Scholar] [CrossRef]
- Wang, J.C.; Tang, L.L.; Glenn, T.C.; Wang, J.S. Aflatoxin B-1 induced compositional changes in gut microbial communities of male F344 rats. Toxicol. Sci. 2016, 150, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tang, L.; Wang, J.; Wang, J.S. Aflatoxin B1 disrupts gut-microbial metabolisms of short-chain fatty acids, long-chain fatty acids, and bile acids in male F344 rats. Toxicol. Sci. 2018, 164, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, L.; Chen, J.; Xiao, A. Response of intestinal bacterial flora to the long-term feeding of aflatoxin B1 (AFB1) in mice. Toxins 2017, 9, 317. [Google Scholar] [CrossRef]
- Salem, D.A. Natural Occurance of Aflatoxins in Feedstuffs and Milk of Dairy Farms in Assiut Porvince, Egypt. Wien. Tierarztl. Mon. 2002, 89, 86–91. [Google Scholar]
- Abrar, M.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Randhawa, M.A.; Saeed, F.; Waqas, K. Aflatoxins: Biosynthesis, occurrence, toxicity, and remedies. Crit. Rev. Food Sci. Nutr. 2013, 53, 862–874. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, M.Z.; Khan, A.; Javed, I.; Asi, M.R. Toxico-pathological effects in rats induced by concurrent exposure to aflatoxin and cypermethrin. Toxiconology 2009, 53, 33–41. [Google Scholar] [CrossRef]
- Trebak, F.; Alaoui, A.; Alexandre, D.; El Ouezzani, S.; Anouar, Y.; Chartrel, N.; Magoul, R. Impact of aflatoxin B1 on hypothalamic neuropeptides regulating feeding behavior. NeuroToxicology 2015, 49, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Wang, F.; Tang, L.; Massey, M.E.; Mitchell, N.J.; Su, J.; Williams, J.H.; Phillips, T.D.; Wang, J.-S. Integrative toxicopathological evaluation of aflatoxin B1 exposure in F344 rats. Toxicol. Pathol. 2013, 41, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Benkerroum, N. Chronic and acute toxicities of aflatoxins: Mechanisms of action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Yu, P.; Yang, K.; Cao, D. Aflatoxin B1: Metabolism, toxicology, and its involvement in oxidative stress and cancer development. Toxicol. Mech. Methods 2022, 32, 395–419. [Google Scholar] [CrossRef]
- Rotimi, O.A.; Rotimi, S.O.; Duru, C.U.; Ebebeinwe, O.J.; Abiodun, A.O.; Oyeniyi, B.O.; Faduyile, F.A. Acute aflatoxin B1—Induced hepatotoxicity alters gene expression and disrupts lipid and lipoprotein metabolism in rats. Toxicol. Rep. 2017, 4, 408–414. [Google Scholar] [CrossRef]
- Tadele, F.; Demissie, B.; Amsalu, A.; Demelash, H.; Mengist, Z.; Ambelu, A.; Yenew, C. Aflatoxin contamination of animal feeds and its predictors among dairy farms in Northwest Ethiopia: One Health approach implications. Front. Vet. Sci. 2023, 10, 1123573. [Google Scholar] [CrossRef]
- Rawat, M.; Varshney, A.; Rai, M.; Chikara, A.; Pohty, A.L.; Joshi, A.; Binjola, A.; Singh, C.P.; Rawat, K.; Rather, M.A.; et al. A comprehensive review on nutraceutical potential of underutilized cereals and cereal-based products. J. Agric. Food Res. 2023, 12, 100619. [Google Scholar] [CrossRef]
- Bilalis, D.; Roussis, I.; Kakabouki, I.; Folina, A. Quinoa (Chenopodium quinoa Willd.) crop under Mediterranean conditions: A review. Cienc. Investig. Agrar. 2019, 46, 51–68. [Google Scholar] [CrossRef]
- Ferrari, L.; Rizzi, N.; Grandi, E.; Clerici, E.; Tirloni, E.; Stella, S.; Bernardi, C.E.M.; Pinotti, L. Compliance between Food and Feed Safety: Eight-Year Survey (2013–2021) of Aflatoxin M1 in Raw Milk and Aflatoxin B1 in Feed in Northern Italy. Toxins 2023, 15, 168. [Google Scholar] [CrossRef]
- Smaoui, S.; D’Amore, T.; Agriopoulou, S.; Mousavi Khaneghah, A. Mycotoxins in Seafood: Occurrence, Recent Development of Analytical Techniques and Future Challenges. Separations 2023, 10, 217. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in Analysis and Detection of Major Mycotoxins in Foods. Foods 2020, 9, 518. [Google Scholar] [CrossRef] [PubMed]
- RASFF (Rapid Alert System for Food and Feed). Available online: https://food.ec.europa.eu/system/files/2023-08/acn_annual-report_2022.pdf (accessed on 19 August 2023).
- FOODAKAI. EU RASFF Alerts for Aflatoxins from 4 January 2016 until 2 March 2022. 2022. Available online: https://app.foodakai.com/users/sign_in (accessed on 29 August 2023).
- Souza, C.; Khaneghah, A.M.; Oliveira, C.A.F. The occurrence of aflatoxin M1 in industrial and traditional fermented milk: A systematic review study. Ital. J. Food Sci. 2021, 33, 12–23. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Kos, J.; Anić, M.; Radić, B.; Zadravec, M.; Janić Hajnal, E.; Pleadin, J. Climate Change—A Global Threat Resulting in Increasing Mycotoxin Occurrence. Foods 2023, 12, 2704. [Google Scholar] [CrossRef]
- Beyene, A.M.; Du, X.; Schrunk, D.; Ensley, S.; Rumbeiha, W.K. High-performance liquid chromatography and Enzyme-Linked Immunosorbent Assay techniques for detection and quantification of aflatoxin B1 in feed samples: A comparative study. BMC Res. Notes 2019, 12, 492. [Google Scholar] [CrossRef]
- Loi, M.; Logrieco, A.F.; Pusztahelyi, T.; Leiter, É.; Hornok, L.; Pócsi, I. Advanced mycotoxin control and decontamination techniques in view of an increased aflatoxin risk in Europe due to climate change. Front. Microbiol. 2023, 13, 1085891. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Vermeulen, L.C.; Gavai, A.K.; Liu, C. Climate change impacts on aflatoxin B1 in maize and aflatoxin M1 in milk: A case study of maize grown in Eastern Europe and imported to the Netherlands. PLoS ONE 2019, 14, e0218956. [Google Scholar] [CrossRef]
- Kos, J.; Lević, J.; Đuragić, O.; Kokić, B.; Miladinović, I. Occurrence and estimation of aflatoxin M1 exposure in milk in Serbia. Food Control 2014, 38, 41–46. [Google Scholar] [CrossRef]
- Milićević, D.; Pleadin, J. Mycotoxin incidence in southeastern European countries: Implications for the food supply chain. In Sustainable and Nutrition-Sensitive Food Systems for Healthy Diets and Prevention of Malnutrition in Europe and Central Asia; Fang, C., Gurinović, M., Eds.; FAO: Budapest, Hungary, 2023; pp. 117–149. [Google Scholar]
- Pleadin, J.; Kos, J.; Radić, B.; Vulić, A.; Kudumija, N.; Radović, R.; Janić Hajnal, E.; Mandić, A.; Anić, M. Aflatoxins in Maize from Serbia and Croatia: Implications of Climate Change. Foods 2023, 12, 548. [Google Scholar] [CrossRef]
- Álvarez-Días, F.; Torres-Parga, B.; Valdivia-Flores, A.G.; Quezada-Tristán, T.; Alejos-De La Fuente, J.I.; Sosa-Ramírez, J.; Rangel-Muñoz, E.J. Aspergillus flavus and Total Aflatoxins Occurrence in Dairy Feed and Aflatoxin M1 in Bovine Milk in Aguascalientes, Mexico. Toxins 2022, 14, 292. [Google Scholar] [CrossRef]
- Shen, M.H.; Singh, R.K. Determining aflatoxins in raw peanuts using immunoaffinity column as sample clean-up method followed by normal-phase HPLC-FLD analysis. Food Control 2022, 139, 109065. [Google Scholar] [CrossRef]
- Asghar, M.A.; Ahmed, A.; Iqbal, J.; Zahir, E.; Nauman, H. Fungal flora and aflatoxin contamination in Pakistani wheat kernels (Triticum aestivum L.) and their attribution in seed germination. J. Food Drug Anal. 2016, 24, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Khayoon, W.S.; Saad, B.; Yan, C.B.; Hashim, N.H.; Ali, A.S.M.; Salleh, M.I.; Salleh, B. Determination of aflatoxins in animal feeds by HPLC with multifunctional column clean-up. Food Chem. 2010, 118, 882–886. [Google Scholar] [CrossRef]
- Blesa, J.; Soriano, J.M.; Molto, J.C.; Marın, R.; Manes, J. Determination of aflatoxins in peanuts by matrix solid-phase dispersion and liquid chromatography. J. Chromatog. A 2003, 1011, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin contamination in the EU feed supply chain: A focus on cereal byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef]
- Kumar, A.; Dhanshetty, M.; Banerjee, K. Development and validation of a method for direct analysis of aflatoxins in animal feeds by ultra-high-performance liquid chromatography with fluorescence detection. J. AOAC Int. 2020, 103, 940–945. [Google Scholar] [CrossRef]
- Hu, Y.Y.; Zheng, P.; Zhang, Z.X.; He, Y.Z. Determination of aflatoxins in high-pigment content samples by matrix solid-phase dispersion and high-performance liquid chromatography. J. Agric. Food Chem. 2006, 54, 4126–4130. [Google Scholar] [CrossRef]
- Kosicki, R.; Błajet-Kosicka, A.; Grajewski, J.; Twarużek, M. Multiannual mycotoxin survey in feed materials and feedingstuffs. Anim. Feed Sci. Technol. 2016, 215, 165–180. [Google Scholar] [CrossRef]
- Šegvić Klarić, M.; Cvetnić, Z.; Pepeljnjak, S.; Kosalec, I. Co-occurrence of aflatoxins, ochratoxin A, fumonisins, and zearalenone in cereals and feed, determined by competitive direct enzyme-linked immunosorbent assay and thin-layer chromatography. Arh. Hig. Rada Toksikol. 2009, 60, 427–433. [Google Scholar] [CrossRef]
- Martins, H.M.; Marques, M.; Almeida, I.; Guerra, M.M.; Bernardo, F. Mycotoxins in feedstuffs in Portugal: An overview. Mycotoxin Res. 2008, 24, 19–23. [Google Scholar] [CrossRef]
- Sahin, H.Z.; Celik, M.; Kotay, S.; Kabak, B. Aflatoxins in dairy cow feed, raw milk and milk products from Turkey. Food Addit. Contam B Surveill. 2016, 9, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Agbetiameh, D.; Ortega-Beltran, A.; Awuah, R.T.; Atehnkeng, J.; Cotty, P.J.; Bandyopadhyay, R. Prevalence of aflatoxin contamination in maize and groundnut in Ghana: Population structure, distribution, and toxigenicity of the causal agents. Plant Dis. 2018, 102, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Ghana Standard Authority. Cereals and Pulses-Specification for Maize (Corn), 3rd ed.; GS 211; Ghana Standard Authority: Accra, Ghana, 2013. [Google Scholar]
- Abdallah, M.F.; Girgin, G.; Baydar, T. Mycotoxin Detection in Maize, Commercial Feed, and Raw Dairy Milk Samples from Assiut City, Egypt. Vet. Sci. 2019, 6, 6020057. [Google Scholar] [CrossRef] [PubMed]
- Madbouly, A.K.; Ibrahim, M.I.; Sehab, A.F.; Abdel-Wahhab, M.A. Co-occurrence of mycoflora, aflatoxins and fumonisins in maize and rice seeds from markets of different districts in Cairo, Egypt. Food Addit. Contam B Surveill. 2012, 5, 112–120. [Google Scholar] [CrossRef]
- Abdelhamid, A.M. Occurrence of some mycotoxins (aflatoxin, ochratoxin A, citrinin, zearalenone and vomitoxin) in various Egyptian feeds. Arc. Anim. Nutr. 1990, 40, 647–664. [Google Scholar] [CrossRef]
- Lutfullah, G.; Hussain, A. Studies on contamination level of aflatoxins in some cereals and beans of Pakistan. Food Control 2012, 23, 32–36. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No. 1881/2006 of 19 December 2006. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- Getachew, A.; Chala, A.; Hofgaard, I.S.; Brurberg, M.B.; Sulyok, M.; Tronsmo, A.M. Multimycotoxin and fungal analysis of maize grains from south and southwestern Ethiopia. Food Addit. Contam B Surveill. 2018, 11, 64–74. [Google Scholar] [CrossRef]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef]
- Giusiano, G.E.; Piontelli, E.; Fernández, M.S.; Mangiaterra, M.L.; Cattana, M.E.; Kocsubé, S.; Varga, J. Biodiversity of species of Aspergillus section Fumigati in semi-desert soils in Argentina. Rev. Argent. Microb. 2017, 49, 247–254. [Google Scholar] [CrossRef]
- Atukwase, A.; Kaaya, A.N.; Nuyanja, C. Factors associated with fumonisin contamination of maize in Uganda. J. Sci. Food Agric. 2009, 89, 2393–2398. [Google Scholar] [CrossRef]
- Fapohunda, S.O.; Adewunmi, A.A. Climate change and mycotoxins-The African experience. Croat. J. Food Sci. Technol. 2019, 11, 283–290. [Google Scholar] [CrossRef]
- Conteçotto, A.C.T.; Pante, G.C.; Castro, J.C.; Souza, A.A.; Lini, R.S.; Romoli, J.C.Z.; Abreu Filho, B.; Mikcha, J.M.G.; Mossini, S.A.G.; Machinski Junior, M. Occurrence, exposure evaluation and risk assessment in child population for aflatoxin M1 in dairy products in Brazil. Food Chem. Toxicol. 2021, 148, 111913. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B. Aflatoxins in foodstuffs: Occurrence and risk assessment in Turkey. J. Food Compos. Anal. 2021, 96, 103734. [Google Scholar] [CrossRef]
- Liu, J.; Sun, L.; Zhang, J.; Guo, J.; Chen, L.; Qi, D.; Zhang, N. Aflatoxin B1, zearalenone and deoxynivalenol in feed ingredients and complete feed from central China. Food Addit. Contam B Surveill. 2016, 9, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sun, L. Occurrence of Aflatoxin B1, deoxynivalenol and zearalenone in feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Pervaiz, W.; Zia, K.M.; Iqbal, S.Z. Assessment of aflatoxin B1 in animal feed and aflatoxin M1 in raw milk samples of different species of milking animals from Punjab, Pakistan. J. Food Saf. 2021, 41, e12893. [Google Scholar] [CrossRef]
- Romero, E.T.A.; Cumagun, C.J.R. Aflatoxin levels and Aspergillus species in maize in the Province of Isabela, Philippines. J. Phytopathol. 2023, 171, 489–503. [Google Scholar] [CrossRef]
- Aloui, A.; Salah-Abbès, J.B.; Zinedine, A.; Meile, J.C.; Riba, A.; Durand, N.; Montet, D.; Abbès, S.; Brabet, C. Occurrence of pre-and postharvest multi-mycotoxins in durum wheat grains collected in 2020 and 2021 in two climatic regions of Tunisia. Food Addit. Contam B Surveill. 2023, 16, 274–287. [Google Scholar] [CrossRef]
- Nsiah, B.; Ofori, H.; Oduro-Yeboah, C.; Kyereh, E.; Johnson, P.N.T. Incidences of Aflatoxin Contaminations in Ingredients, Feed and Products of Poultry from Two Regions in Ghana. Available online: http://dx.doi.org/10.2139/ssrn.4502881 (accessed on 20 September 2023).
- Magomya, A.M.; Mbatsav, T.O. Analysis and Health Risk Evaluation of Aflatoxin B1 Levels in Groundnut (Arachis hypogea) and Maize (Zea mays) Samples from Wukari, Nigeria. EJTAS 2023, 1, 886–893. [Google Scholar] [CrossRef]
- Rodríguez-Blanco, M.; Ramos, A.J.; Prim, M.; Sanchis, V.; Marín, S. Usefulness of the analytical control of aflatoxins in feedstuffs for dairy cows for the prevention of aflatoxin M1 in milk. Mycotoxin Res. 2020, 36, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Solano, B.; González-Peñas, E. Mycotoxin determination in animal feed: An LC-FLD method for simultaneous quantification of aflatoxins, ochratoxins and zearelanone in this matrix. Toxins 2020, 12, 374. [Google Scholar] [CrossRef] [PubMed]
- Smaoui, S.; Ben Braïek, O.; Ben Hlima, H. Mycotoxins analysis in cereals and related foodstuffs by liquid chromatography-tandem mass spectrometry techniques. J. Food Qual. 2020, 2020, 1–23. [Google Scholar] [CrossRef]
- Mushtaq, M.; Sultana, B.; Anwar, F.; Khan, M.Z.; Ashrafuzzaman, M. Occurrence of aflatoxins in selected processed foods from Pakistan. Int. J. Mol. Sci. 2012, 13, 8324–8337. [Google Scholar] [CrossRef]
- Oulkar, D.; Goon, A.; Dhanshetty, M.; Khan, Z.; Satav, S.; Banerjee, K. High-sensitivity direct analysis of aflatoxins in peanuts and cereal matrices by ultra-performance liquid chromatography with fluorescence detection involving a large volume flow cell. J. Environ. Sci. Health B 2018, 53, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, E.; Güvenç, D.; Aksoy, A. Determination of aflatoxin levels in maize grain by high performance liquid chromatography using an immunoaffinity column cleanup. Kafkas Univ. Vet. Fak. Derg. 2015, 21, 619–622. [Google Scholar]
- Imperato, R.; Campone, L.; Piccinelli, A.L.; Veneziano, A.; Rastrelli, L. Survey of aflatoxins and ochratoxin a contamination in food products imported in Italy. Food Control 2011, 22, 1905–1910. [Google Scholar] [CrossRef]
- Torović, L. Aflatoxins and ochratoxin A in flour: A survey of the Serbian retail market. Food Addit. Contam B Surveill. 2018, 11, 26–32. [Google Scholar] [CrossRef]
- Hell, K.; Fandohan, P.; Bandyopadhyay, R.; Kiewnick, S.; Sikora, R.; Cotty, P.J. Pre-and post-harvest management of aflatoxin in maize: An African perspective. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; Leslie, J.F., Bandyopadhyay, R., Visconti, A., Eds.; CAB International: Wallingford, UK, 2008; p. 219. ISBN 9781845930820. [Google Scholar]
- Kassaw, T.S.; Megerssa, Y.C.; Woldemariyam, F.T. Occurrence of Aflatoxins in Poultry Feed in Selected Chicken Rearing Villages of Bishoftu Ethiopia. Vet. Med. Res. Reports 2022, 13, 277–286. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, H. Development of a microwave-assisted-extraction-based method for the determination of aflatoxins B1, G1, B2, and G2 in grains and grain products. Anal. Bioanal. Chem. 2013, 405, 1623–1630. [Google Scholar] [CrossRef]
- Gizachew, D.; Szonyi, B.; Tegegne, A.; Hanson, J.; Grace, D. Aflatoxin contamination of milk and dairy feeds in the Greater Addis Ababa milk shed, Ethiopia. Food Control 2016, 59, 773–779. [Google Scholar] [CrossRef]
- Bahrami, R.; Shahbazi, Y.; Nikousefat, Z. Occurrence and seasonal variation of aflatoxin in dairy cow feed with estimation of aflatoxin M1 in milk from Iran. Food Agric. Immunol. 2016, 27, 388–400. [Google Scholar] [CrossRef]
- Kenei, F.; Mezene Woyessa, A.M.; Lema, K.; Gemmeda, M. Review on Occurrence and Public Health Significance of Aflatoxin in Dairy Products and Feed. J. Livest. Res. 2023, 13, 1–13. [Google Scholar]
- Oliveira, M.S.; Rocha, A.; Sulyok, M.; Krska, R.; Mallmann, C.A. Natural mycotoxin contamination of maize (Zea mays L.) in the South region of Brazil. Food Control 2017, 73, 127–132. [Google Scholar] [CrossRef]
- Dimitrieska-Stojković, E.; Stojanovska-Dimzoska, B.; Ilievska, G.; Uzunov, R.; Stojković, G.; Hajrulai-Musliu, Z.; Jankuloski, D. Assessment of aflatoxin contamination in raw milk and feed in Macedonia during 2013. Food Control 2016, 59, 201–206. [Google Scholar] [CrossRef]
- Kim, N.-H.; Hong, S.-Y.; Kang, J.W.; Cho, S.M.; Lee, K.R.; An, T.K.; Lee, C.; Chung, S.H. Simultaneous determination of multi-mycotoxins in cereal grains collected from South Korea by LC/MS/MS. Toxins 2017, 9, 106. [Google Scholar] [CrossRef]
- Keller, L.A.M.; Aronovich, M.; Keller, K.M.; Castagna, A.A.; Cavaglieri, L.R.; Rosa, C.A.D.R. Incidence of mycotoxins (AFB1 and AFM1) in feeds and dairy farms from Rio de Janeiro State, Brazil. Veter-Med. Open J. 2016, 1, 29–35. [Google Scholar] [CrossRef]
- Janić Hajnal, E.; Kos, J.; Krulj, J.; Krstović, S.; Jajić, I.; Pezo, L.; Šarić, B.; Nedeljković, N. Aflatoxins contamination of maize in Serbia: The impact of weather conditions in 2015. Food Addit. Contam. Part A 2017, 34, 1999–2010. [Google Scholar] [CrossRef]
- Hanvi, M.D.; Lawson-Evi, P.; Bouka, E.C.; Eklu-Gadegbeku, K. Aflatoxins in maize dough and dietary exposure in rural populations of Togo. Food Control 2021, 121, 107673. [Google Scholar] [CrossRef]
- Diaz, G.J.; Krska, R.; Sulyok, M. Mycotoxins and cyanogenic glycosides in staple foods of three indigenous people of the Colombian Amazon. Food Addit. Contam B Surveill. 2015, 8, 291–297. [Google Scholar] [CrossRef]
- Torres, A.M.; Barros, G.G.; Palacios, S.A.; Chulze, S.N.; Battilani, P. Review on pre-and post-harvest management of peanuts to minimize aflatoxin contamination. Food Res. Int. 2014, 62, 11–19. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Reverberi, M.; Geisen, R. Mycotoxins in harvested fruits and vegetables: Insights in producing fungi, biological role, conducive conditions, and tools to manage postharvest contamination. Postharvest Biol. Technol. 2016, 122, 95–105. [Google Scholar] [CrossRef]
- Loi, M.; Paciolla, C.; Logrieco, A.F.; Mulè, G. Plant Bioactive Compounds in Pre- and Postharvest Management for Aflatoxins Reduction. Front. Microbiol. 2020, 11, 243. [Google Scholar] [CrossRef] [PubMed]
- Wu, F. Mycotoxin reduction in Bt corn: Potential economic, health, and regulatory impacts. Transgenic Res. 2006, 15, 277–289. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Leszkowicz, A. Comparative data concerning aflatoxin contents in Bt maize and non-Bt isogenic maize in relation to human and animal health—A review. Acta Vet. Brno. 2015, 84, 47–53. [Google Scholar] [CrossRef]
- Wu, F. Mycotoxin risks are lower in biotech corn. Curr. Opin. Biotechnol. 2022, 78, 102792. [Google Scholar] [CrossRef]
- Rajasekaran, K.; Sayler, R.J.; Majumdar, R.; Sickler, C.M.; Cary, J.W. Inhibition of Aspergillus flavus Growth and Aflatoxin Production in Transgenic Maize Expressing the α-amylase Inhibitor from Lablab purpureus L. J. Vis. Exp. 2019, 144, e59169. [Google Scholar]
- Ferrara, M.; Gallo, A.; Cervini, C.; Gambacorta, L.; Solfrizzo, M.; Baker, S.E.; Perrone, G. Evidence of the involvement of a cyclase gene in the biosynthesis of ochratoxin A in Aspergillus carbonarius. Toxins 2021, 13, 892. [Google Scholar] [CrossRef]
- Moral, J.; Garcia-Lopez, M.T.; Camiletti, B.X.; Jaime, R.; Michailides, T.J.; Bandyopadhyay, R.; Ortega-Beltran, A. Present status and perspective on the future use of aflatoxin biocontrol products. Agronomy 2020, 10, 491. [Google Scholar] [CrossRef]
- Ouadhene, M.A.; Ortega-Beltran, A.; Sanna, M.; Cotty, P.J.; Battilani, P. Multiple Year Influences of the Aflatoxin Biocontrol Product AF-X1 on the A. flavus Communities Associated with Maize Production in Italy. Toxins 2023, 15, 184. [Google Scholar] [CrossRef]
- Regulation (EC). Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. Off. J. Eur. Union 2009, 309, 1–50. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32009R1107 (accessed on 31 August 2023).
- European Commission. Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. Consolidated Version 2022. Available online: http://data.europa.eu/eli/reg/2009/1107/2022-11-21 (accessed on 31 August 2023).
- European Food Safety Authority (EFSA). Alvarez, F.; Arena, M.; Auteri, D.; Binaglia, M.; Castoldi, A.F.; Chiusolo, A.; Colagiorgi, A.; Colas, M.; Crivellente, F.; De Lentdecker, C. Peer review of the pesticide risk assessment of the active substance Aspergillus flavus strain MUCL54911. EFSA J. 2022, 20, e07202. [Google Scholar] [CrossRef] [PubMed]
- Aikore, M.O.S.; Ortega-Beltran, A.; Eruvbetine, D.; Atehnkeng, J.; Falade, T.D.O.; Cotty, P.J.; Bandyopadhyay, R. Performance of Broilers Fed with Maize Colonized by Either Toxigenic or Atoxigenic Strains of Aspergillus flavus with and without an Aflatoxin-Sequestering Agent. Toxins 2019, 11, 565. [Google Scholar] [CrossRef] [PubMed]
- Mamo, F.T.; Shang, B.; Selvaraj, J.N.; Zheng, Y.; Liu, Y. Biocontrol efficacy of atoxigenic Aspergillus flavus strains against aflatoxin contamination in peanut field in Guangdong province, South China. Mycology 2022, 13, 143–152. [Google Scholar] [CrossRef]
- Xu, J.; Wang, P.; Zhou, Z.; Cotty, P.J.; Kong, Q. Selection of Atoxigenic Aspergillus flavus for Potential Use in Aflatoxin Prevention in Shandong Province, China. J. Fungi 2021, 7, 773. [Google Scholar] [CrossRef]
- Ortega-Beltran, A.; Moral, J.; Picot, A.; Puckett, R.D.; Cotty, P.J.; Michailides, T.J. Atoxigenic Aspergillus flavus isolates endemic to almond, fig, and pistachio orchards in California with potential to reduce aflatoxin contamination in these crops. Plant Dis. 2019, 103, 905–912. [Google Scholar] [CrossRef] [PubMed]
- European Commission. EU Pesticides Database—Active Substance: Aspergillus flavus strain MUCL 54911. Available online: https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/start/screen/active-substances/details/1401 (accessed on 19 August 2023).
- Bhatnagar-Mathur, P.; Sunkara, S.; Bhatnagar-Panwar, M.; Waliyar, F.; Sharma, K.K. Biotechnological advances for combating Aspergillus flavus and aflatoxin contamination in crops. Plant Sci. 2015, 234, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Masanga, J.O.; Matheka, J.M.; Omer, R.A.; Ommeh, S.C.; Monda, E.O.; Alakonya, A.E. Downregulation of transcription factor aflR in Aspergillus flavus confers reduction to aflatoxin accumulation in transgenic maize with alteration of host plant architecture. Plant Cell Rep. 2015, 34, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Thakare, D.; Zhang, J.; Wing, R.A.; Cotty, P.J.; Schmidt, M.A. Aflatoxin-free transgenic maize using host-induced gene silencing. Sci. Adv. 2017, 3, e1602382. [Google Scholar] [CrossRef]
- Lagogianni, C.S.; Tsitsigiannis, D.I. Effective chemical management for prevention of aflatoxins in maize. Phytopathol. Mediterr. 2018, 57, 186–197. [Google Scholar]
- Ferrigo, D.; Mondin, M.; Scopel, C.; Maso, E.D.; Stefenatti, M.; Raiola, A.; Causin, R. Effects of a prothioconazole- and tebuconazolebased fungicide on Aspergillus flavus development under laboratory and field conditions. Eur. J. Plant Pathol. 2019, 155, 151–161. [Google Scholar] [CrossRef]
- Masiello, M.; Somma, S.; Haidukowski, M.; Logrieco, A.F.; Moretti, A. Genetic polymorphisms associated to SDHI fungicides resistance in selected Aspergillus flavus strains and relation with aflatoxin production. Int. J. Food Microbiol. 2020, 334, 108799. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Stafstrom, W.; Priest, P.; Fuchs, J.; Windham, G.L.; Williams, W.P.; Nelson, R.J. Low-cost grain sorting technologies to reduce mycotoxin contamination in maize and groundnut. Food Control 2020, 118, 107363. [Google Scholar] [CrossRef] [PubMed]
- Sipos, P.; Peles, F.; Brassó, D.L.; Béri, B.; Pusztahelyi, T.; Pócsi, I.; Győri, Z. Physical and Chemical Methods for Reduction in Aflatoxin Content of Feed and Food. Toxins 2021, 13, 204. [Google Scholar] [CrossRef]
- Yao, H.; Zhu, F.; Kincaid, R.; Hruska, Z.; Rajasekaran, K. A Low-Cost, Portable Device for Detecting and Sorting Aflatoxin-Contaminated Maize Kernels. Toxins 2023, 15, 197. [Google Scholar] [CrossRef]
- Marshall, H.; Meneely, J.P.; Quinn, B.; Zhao, Y.; Bourke, P.; Gilmore, B.F.; Zhang, G.; Elliott, C.T. Novel decontamination approaches and their potential application for post-harvest aflatoxin control. Trends Food Sci. Technol. 2020, 106, 489–496. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Su, Q. The Toxification and Detoxification Mechanisms of Aflatoxin B1 in Human: An Update. In Aflatoxin B1 Occurrence, Detection and Toxicological Effects; InTech Open: Rijeka, Croatia, 2019; pp. 1–17. [Google Scholar]
- Di Stefano, V.; Pitonzo, R.; Cicero, N.; D’Oca, M.C. Mycotoxin contamination of animal feedingstuff: Detoxification by gamma-irradiation and reduction of aflatoxins and ochratoxin A concentrations. Food Addit. Contam. Part A 2014, 31, 2034–2039. [Google Scholar] [CrossRef]
- Faraji, H.; Yazdi, F.T.; Razmi, N. The influence of ultraviolet radiation on aflatoxin producing Aspergillus species’ isolated from Iranian rice. Toxicol. Rep. 2022, 9, 1528–1536. [Google Scholar] [CrossRef]
- El-Desouky, T.A.; Sharoba, A.M.A.; El-Desoukym, A.I.; El-Mansy, H.A.; Naguib, K. Effect of Ozone Gas on Degradation of Aflatoxin B1 and Aspergillus flavus Fungal. J. Environ. Anal. Toxicol. 2012, 2, 128. [Google Scholar]
- Agriopoulou, S.; Koliadima, A.; Karaiskakis, G.; Kapolos, J. Kinetic study of aflatoxins’ degradation in the presence of ozone. Food Control 2016, 61, 221–226. [Google Scholar] [CrossRef]
- Luo, X.; Wang, R.; Wang, L.; Li, Y.; Bian, Y.; Chen, Z. Effect of ozone treatment on aflatoxin B1 and safety evaluation of ozonized corn. Food Control 2014, 37, 171–176. [Google Scholar] [CrossRef]
- Kong, C.; Shin, S.Y.; Kim, B.G. Evaluation of mycotoxin sequestering agents for aflatoxin and deoxynivalenol: An in vitro approach. SpringerPlus 2014, 3, 346. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, L.; Gong, G.; Zhang, L.; Shi, L.; Dai, J.; Han, Y.; Wu, Y.; Khalil, M.M.; Sun, L. Invited Review: Remediation Strategies for Mycotoxin Control in Feed. J. Anim. Sci. Biotechnol. 2022, 13, 19. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on additives for use in animal nutrition. Off. J. Eur. Union 2003, 29, L268. [Google Scholar]
- Wang, X.; Wang, T.; Nepovimova, E.; Long, M.; Wu, W.; Kuca, K. Progress on the detoxification of aflatoxin B1 using natural anti-oxidants. Food Chem Toxicol. 2022, 169, 113417. [Google Scholar] [CrossRef] [PubMed]
- IARC. Mycotoxin Control in Low- and Middle-Income Countries; Wild, C.P., Miller, J.D., Groopman, J.D., Eds.; IARC: Lyon, France, 2015. [Google Scholar]
- Guan, Y.; Chen, J.; Nepovimova, E.; Long, M.; Wu, W.; Kuca, K. Aflatoxin Detoxification Using Microorganisms and Enzymes. Toxins 2021, 13, 46. [Google Scholar] [CrossRef]
- Zhang, F.; Geng, L.; Deng, J.; Huang, L.; Zhong, H.; Xin, S.; Fasoyin, O.E.; Wang, S. The MAP kinase AflSlt2 modulates aflatoxin biosynthesis and peanut infection in the fungus Aspergillus flavus. Int. J. Food Microbiol. 2020, 322, 108576. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, L.; Ma, Q.; Li, X.; Shi, H.; Zhou, T.; Zhang, J.; Ji, C. Effects of Bacillus subtilis ANSB060 on growth performance, meat quality and aflatoxin residues in broilers fed moldy peanut meal naturally contaminated with aflatoxins. Food Chem. Toxicol. 2013, 59, 748–753. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic acid bacteria as antibacterial agents to extend the shelf-life of fresh and minimally processed fruits and vegetables: Quality and safety aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Ma, Z.X.; Amaro, F.X.; Romero, J.J.; Pereira, O.G.; Jeong, K.C.; Adesogan, A.T. The capacity of silage inoculant bacteria to bind aflatoxin B1 in vitro and in artificially contaminated corn silage. J. Dairy Sci. 2017, 100, 7198–7210. [Google Scholar] [CrossRef] [PubMed]
- Oluwafemi, F.; Kumar, M.; Bandyopadhyay, R.; Ogunbanwo, T.; Ayanwande, B. Bio-detoxification of aflatoxin B1 in artificially contaminated maize grains using lactic acid bacteria. Toxin Rev. 2010, 29, 115–122. [Google Scholar] [CrossRef]
- Liu, J.; Song, W.J.; Zhang, N.Y.; Tan, J.; Krumm, C.S.; Sun, L.H.; Qi, D.S. Biodetoxification of aflatoxin B1 in cottonseed meal by fermentation of Cellulosimicrobium funkei in duckling diet. Poult. Sci. 2017, 96, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Kusumaningtyas, E.; Widiastuti, R.; Maryam, R. Reduction of aflatoxin B1 in chicken feed by using Saccharomyces cerevisiae, Rhizopus oligosporus and their combination. Mycopathologia 2006, 162, 307–311. [Google Scholar] [CrossRef]
- Zhou, G.; Chen, Y.; Kong, Q.; Ma, Y.; Liu, Y. Detoxification of Aflatoxin B1 by Zygosaccharomyces rouxii with Solid State Fermentation in Peanut Meal. Toxins 2017, 9, 42. [Google Scholar] [CrossRef] [PubMed]


| Continent | Country | Aflatoxins | No. of Samples | Positive Samples (%) | Range (µg/Kg) | Method | Reference |
|---|---|---|---|---|---|---|---|
| Asia | China | AFB1 | 2083 | 33.9 | 0.6–1818.0 | HPLC/FLD | [114] |
| 3500 | 81.9–100 | 1.2–27.4 | HPLC/FLD | [115] | |||
| Pakistan | AFB1 | 193 | 65.3 | LOD to 180.5 | HPLC/FLD | [116] | |
| Philippines | AFs | 107 | 50.5 | <3–1663 | ELISA | [117] | |
| Africa | Tunisia | AFB1 | 136 | 60.9 | 17–37.8 | UHPLCMS/MS | [118] |
| Ghana | AFs | 60 | 100 | 5.32–29.88 | HPLC/FLD | [119] | |
| Nigeria | AFB1 | 10 | 100 | 1.48–15.50 | TLC | [120] | |
| Ethiopia | AFs | 100 | 96 | LOD to 306.9 | ELISA | [71] | |
| North America | Mexico | AFs | 170 | 100 | 25.9–27.0 | HPLC/FLD | [88] |
| Europe | Spain | AFs | 193 | 34.7 | 0.05–6.45 | LC/FLD | [121] |
| 10 | n.d. 1 | n.d. 1 | HPLC/FLD | [122] |
| Country | Sample | Extraction and Clean up | Detection | Analytes | LOD Range | LOQ Range | Validation Parameters | References |
|---|---|---|---|---|---|---|---|---|
| India | Rice | Phosphate-buffered saline, loaded on IAC: (AFLAOCHRAPREP®) and eluted with MeOH | UHPLC-FLD | AFs (B1, B2, G1 and G2) | 0.02 ng/g | 0.25–1 ng/g | Linearity, sensitivity, accuracy and precision | [129] |
| India | Peanut-processed products and cereal (rice, corn, millets) | IAC and MeOH | UHPLC-FLD | AFs | - | -AFB1, AFG1: 0.025 mg/kg -AFB2, AFG2: 0.01 mg/kg | Selectivity, linearity, recovery and precision | [130] |
| India | Pigeon pea husk, poultry feed and wheat bran feed | IAC and MeOH | UHPLC-FLD | AFs | - | 0.5–2 ng/g | Linearity, sensitivity, accuracy and precision. | [117] |
| China | Chili powder, green bean, and black sesame | IAC | UHPLC-FLD | AFs | 0.1–0.25 ng/g | - | Linearity, accuracy and precision | [95] |
| China | Corn and wheat | Solid-phase extraction (SPE) | LC-FLD | AFs | 0.04–25 μg/kg | 0.11–0.61 μg/kg | Recovery and repeatability | [131] |
| Ethiopia | Maize, wheat bran, dairy feeds | MeCN | ELISA (Optical density) | AFB1 and AFM1 | - | - | Recovery and coefficient of variation (%CV) | [132] |
| Iran | Corn silage; corn; wheat; soybean nutrients dairy cow | MeOH | ELISA (450 nm) | AFB1 | - | - | - | [133] |
| Pakistan | Wheat | Easi-Extract® AF IAC | HPLC | AFs | 0.028–0.091 µg/kg | 0.066–0.273 µg/kg | Recovery | [124] |
| Iran | Corn silage | C18 SPE Column and MeOH | HPLC–FLD | AFs | 0.015–0.12 µg/kg | 0.05–0.4 µg/kg | Accuracy | [125] |
| China | Maize; wheat; animal feed | AokinImmunoClean CF AFLA | HPLC | AFB1 | 0.5 µg/kg | 1.5 µg/kg | Recovery | [134] |
| Brazil | Soybean kernels | IAC AflaStarTM Fit | HPLC-FLD | AFB1 | 0.13 µg/kg | 0.37 µg/kg | Selectivity, linearity, accuracy and precision | [135] |
| Macedonia | Maize; dairy cow feed | AflaPrep® IAC SPE | HPLC–FLD | AFB1 | 0.005 µg/kg | 0.014 µg/kg | Linearity, decision limits (CCα), CCβ, precision (repeatability), and recovery | [136] |
| Republic of Korea | Corn matrices | AflaTest® WB IAC | HPLC-FLD | AFs | 0.01–0.17 µg/kg | 0.01–0.51 µg/kg | Linearity, accuracy, and precision | [137] |
| Turkey | Dairy cow feeds | AflaTest® IAC | HPLC-FLD | AFs | 0.046–0.059 µg/kg | 0.153–0.197 µg/kg | Selectivity, recovery and precision | [99] |
| Brazil | Dairy cattle feed | AflaTest® IAC | HPLC | AFB1 | - | - | - | [138] |
| Serbia | Maize | MycoSep®224AflaZon SPE | HPLC-FLD | AFs | 0.2–0.6 µg/kg | 0.6–1.8 µg/kg | Linearity, recovery, repeatability and reproducibility | [139] |
| Ghana | Maize and groundnut | - | TLC (EtO2-MeOH-H2O2 (96:3:1) at 365 nm | AFs | 341 µg/kg | - | - | [100] |
| Togo | Maize | - | HPLC | AFB1 | 0.08 μg/kg | - | - | [140] |
| Colombia | Corn, rice, and cassava | LC–MS/MS | AFs | - | - | - | [141] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smaoui, S.; D’Amore, T.; Tarapoulouzi, M.; Agriopoulou, S.; Varzakas, T. Aflatoxins Contamination in Feed Commodities: From Occurrence and Toxicity to Recent Advances in Analytical Methods and Detoxification. Microorganisms 2023, 11, 2614. https://doi.org/10.3390/microorganisms11102614
Smaoui S, D’Amore T, Tarapoulouzi M, Agriopoulou S, Varzakas T. Aflatoxins Contamination in Feed Commodities: From Occurrence and Toxicity to Recent Advances in Analytical Methods and Detoxification. Microorganisms. 2023; 11(10):2614. https://doi.org/10.3390/microorganisms11102614
Chicago/Turabian StyleSmaoui, Slim, Teresa D’Amore, Maria Tarapoulouzi, Sofia Agriopoulou, and Theodoros Varzakas. 2023. "Aflatoxins Contamination in Feed Commodities: From Occurrence and Toxicity to Recent Advances in Analytical Methods and Detoxification" Microorganisms 11, no. 10: 2614. https://doi.org/10.3390/microorganisms11102614