1. Introduction
Eggs are a cheap and nutritious food, with more than 1.2 billion eggs consumed worldwide annually. China is the country with the highest levels of consumption, and it has been found to account for 35% of global egg production [
1], which highlights the importance of food safety in hen eggs. Moreover, the international export of eggs from China is low because of concerns about food safety and quality [
2]. In order to enhance the food safety of eggs and address this issue, it is essential to conduct an initial investigation into the current condition of eggs.
Egg safety concerns are associated with drug residues, illegal or excessive additives, and microbial contamination. The first two are fixed and can be easily avoided by strict laws and regulations as well as farming practices, but the later may dynamically change due to vertical and horizontal transmission during farming and transport processes [
3]. Eggs and egg products are the known food source of 44% of the human cases of salmonellosis in the European Union [
4]. To maintain egg safety from a microbial aspect, the United States Food and Drug Administration (FDA), the United States Department of Agriculture (USDA), and the European Union have established a series of rules (e.g., the Egg Safety Rule, Egg Safety Regulations, Egg Grading, and the European Union’s Regulation No 2160/2003). These rules include criteria for monitoring and testing for
Salmonella Enteritidis in hens, eggs, and their environments, which ensures egg quality standards and safety. National health standards and rules [
2] (GB 2749-2003, SN/T 0422-2010, and NY/T 754-2011) have also been established in China for several microorganisms, including
Salmonella,
Shigella,
Staphylococcus aureus, etc. The bacterial contamination of eggshells at the time of collection affects the final products and may cause foodborne illnesses; thus, the high bacterial load of floor eggs means they cannot be eaten [
5]. In addition, the pathogenic bacteria and the microorganisms that are deposited on eggshells, in egg whites, and in egg yolks can affect egg quality and shelf life.
Recently, the V3-V4 variable region or full-length sequencing of 16S rRNA genes have been used to explore the microbiota in the guts of chickens [
6], in their feces [
7], on eggshells [
8], and in egg yolks [
9]. The results showed that the bacteria on eggshells changed as a function of the area [
8] and the breed of the chicken [
10], as well as due to farming practices, including the cages [
10], food, and water used [
3]. The microbiota in egg contents were tightly associated with the storage time [
11]. However, information about the impacts of the different seasons on the microbiota on eggshells remains scarce, and these impacts involve changes in temperature, humidity, rainfall, light, etc. Moreover, it is unclear whether the bacterial loads on eggshells from markets are similar to those in the interiors of hen eggs.
Here, hen eggs from local farms, as well as those from markets, were studied with full-length 16S rRNA sequencing to provide basic data on market quality and improve food safety in Shanghai, China.
4. Discussion
The bacteria on the eggshell surfaces demonstrated a high probability of trans-shell infection [
16], and their growth was tightly associated with environmental conditions, including temperature and humidity [
17]. The deposited bacteria on the fresh egg surfaces not only reflected the intestinal health statuses of the hens but also the pollution in the poultry farm environments. Therefore, the bacterial communities on the fresh chicken eggshells were investigated across the four seasons. The results showed that the autumn samples were more seriously contaminated than the spring and summer samples. In addition, during the winter season, the eggshell surfaces harbored greater varieties of microorganisms compared to the summer season (
Figure 1E), which appeared to contrast with the results of a previous study showing that bacteria had higher counts on eggshells exposed to high temperatures compared to those exposed to low temperature [
18]. This may be explained by the differences in the methods for laying and storage. In the present study, we obtained no data from the fresh egg contents because the extracted DNA was insufficient. To further explore contamination in the fresh egg contents, we introduced each sample from the contents onto TSA and blood agar plates. Nevertheless, no bacterial colonies were observed.
Here,
Firmicutes made up 50.87% of the total phyla on the fresh eggs and 51.36% on the market eggshell surfaces, which was consistent with a previous investigation on commercial eggshell surfaces [
4]. Based on the findings that the microbial community in the chicken cloaca as well as in the oviduct was also largely dominated by
Firmicutes [
19,
20], the vertical microbiota transmission was again shown to be critical for the contamination of eggshells and egg safety as described by Trudeau et al. [
7]. However,
Proteobacteria ranked second in the total samples, which was different from a similar study on eggshells from the Shanxi province that showed a majority made up of
Firmicutes and
Actinobacteriota [
8]. These differences may have been due to the different weather or different kinds of hens. The main bacteria producing malodorous trimethylamine in ceca were included in
Firmicutes and
Proteobacteria [
21], and thus, this suggested that the chicken eggs produced in Shanghai were more likely to spoil and should therefore be treated before sale or kept for a shorter time before cooking.
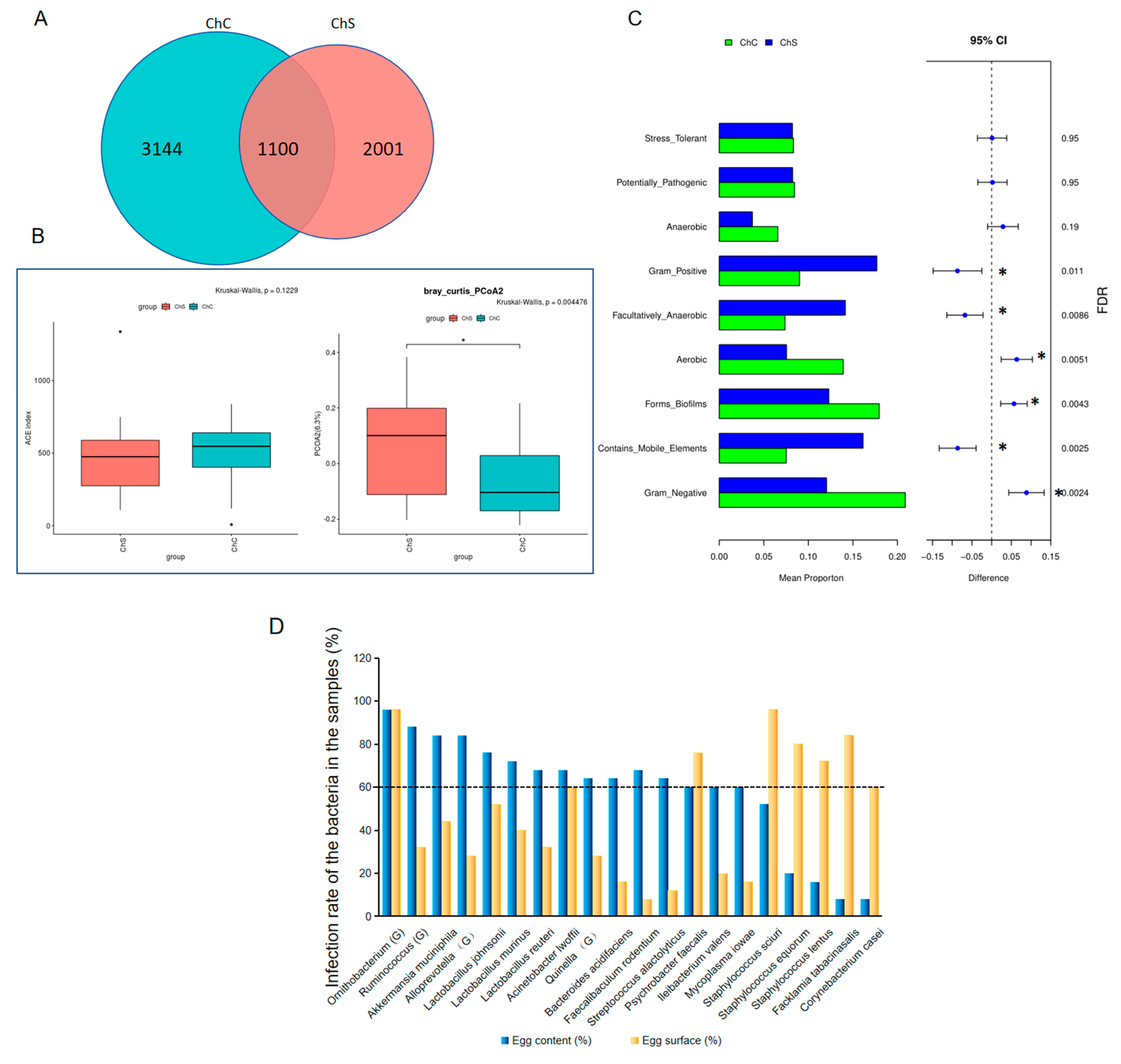
Intriguingly, when the microbiomes were compared to the egg surfaces, Gram-negative, aerobic, and biofilm-forming microorganisms in the contents were significantly abundant, while Gram-positive, facultative anaerobic bacteria and bacteria containing mobile elements were less abundant. We wondered why Gram-negative and biofilm-forming species were more abundant in the egg contents. Bacterial biofilms are communities of cells that are attached to surfaces and to each other, which can protect the imbedded bacteria from the host’s defense system to improve their survival potential [
22]. The biofilms formed by
S. mutans were shown to be highly resistant to harsh environments, host immunity, antimicrobial treatments, and lysozyme [
23]. Therefore, we concluded that the biofilm-forming bacteria including
Pseudomonas may have had more opportunity to overcome the lysozymes and invade the egg whites. As shown in this study,
Pseudomonas had the highest abundances in the market egg contents.
Enterobacteriaceae account for approximately 80% of GNB isolates. The genera/species that frequently affect humans are
Escherichia,
Proteus,
Enterobacter,
Klebsiella,
Citrobacter,
Yersinia, and
Salmonella. GNB have two membranes: external and internal [
24]. The former protects GNB from many antibiotics (including penicillin), detergents (peptidoglycan, which usually damages the inner membranes of cells), and lysozymes. Aminoglycosides, monolactams (aztreonam), and ciprofloxacin are drugs that specifically target Gram-negative organisms. Moreover, GNB can reach almost all systems in the human body and are apt to cause disease in humans [
24] We found a higher abundance of GNB in the egg contents, which requires further research to improve egg safety.
Klebsiella spp. is an opportunistic
Enterobacteriaceae that is highly pathogenic to poultry, livestock, and humans, and it is commonly found throughout China [
25,
26] as well as in other countries [
27,
28]. Here,
Klebsiella contamination on the eggshell surfaces from the poultry farms was detected, with a season-dependent occurrence rate. The highest infection rate was in autumn and the lowest rate was in summer. This could explain the high rates of respiratory diseases in autumn and winter in chickens that have been vaccinated against other respiratory disease-causing pathogens, including the Newcastle disease vaccine and infectious bursal disease vaccines.
Klebsiella spp. can survive for a long time in a henhouse, and the bacteria carried in poultry, its products, such as edible eggs and broiler chickens, and its environment may pose a serious risk to consumers. Moreover, the antibiotic-resistant isolates of
Klebsiella spp. may transfer resistance genes to other microorganisms—especially genes conferring resistance to carbapenems [
28]. Therefore, for the health of both chickens and consumers, surveillance and subsequent controls for this bacterium are supposed to be significant and necessary. In addition, this seasonal distribution trend of
Klebsiella was quite consistent with an investigation on
K. pneumoniae infection carried out in a hospital near Shanghai [
29]. This suggested that poultry, its products, and its environment may be the source of human infection with
K. pneumoniae, which deserves further investigation.
Moreover, we detected a potential pathogen,
R. anatipestifer, on the hen eggshells from a winter sample by 16S rRNA full-length sequencing.
R. anatipestifer is a widely distributed and highly infectious bacterial pathogen for ducklings, goslings, turkeys, and other poultry. It causes pericarditis, perihepatitis, airsaculitis, etc. in its host, which has led to severe economic damage to the duck breeding industry. Moreover, since 2019, chicken-derived
R. anatipestifer has been isolated from sick chicken organs (tarsal joints, sinuses, liver, cheese-containing fallopian tubes, brains, and dead embryo egg fluid) [
30]. In chickens, infection with
R. anatipestifer may not only cause high mortality but also facilitate secondary infections by other bacteria or viruses. In this study, we detected
R. anatipestifer on the normal eggshells, which may have indicated contamination of the environment by
R. anatipestifer due to the mixed rearing of ducks and chickens. Although chicken-derived
R.anatipestifer was not isolated in the samples and no clinical case has been reported in Shanghai, it is imperative to proactively trace a source for early screening and prevention. This is particularly crucial due to the novelty of this pathogen in chicken-derived infections.
Escherichia/shigella has been widely detected and described in gut microbiota and is a negative indicator for human diseases [
31,
32]. Recently, it has been reported as pathogenic or spoilage bacteria that appears on eggshell surfaces as well as in feces [
7]. We also found
Escherichia/shigella in the egg contents in low abundances, except for a few outliers. However, this should be studied further for egg food safety.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}