
Plant-Root Exudate Analogues Influence Activity of the 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Gene in Pseudomonas hormoni G20-18T

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Media
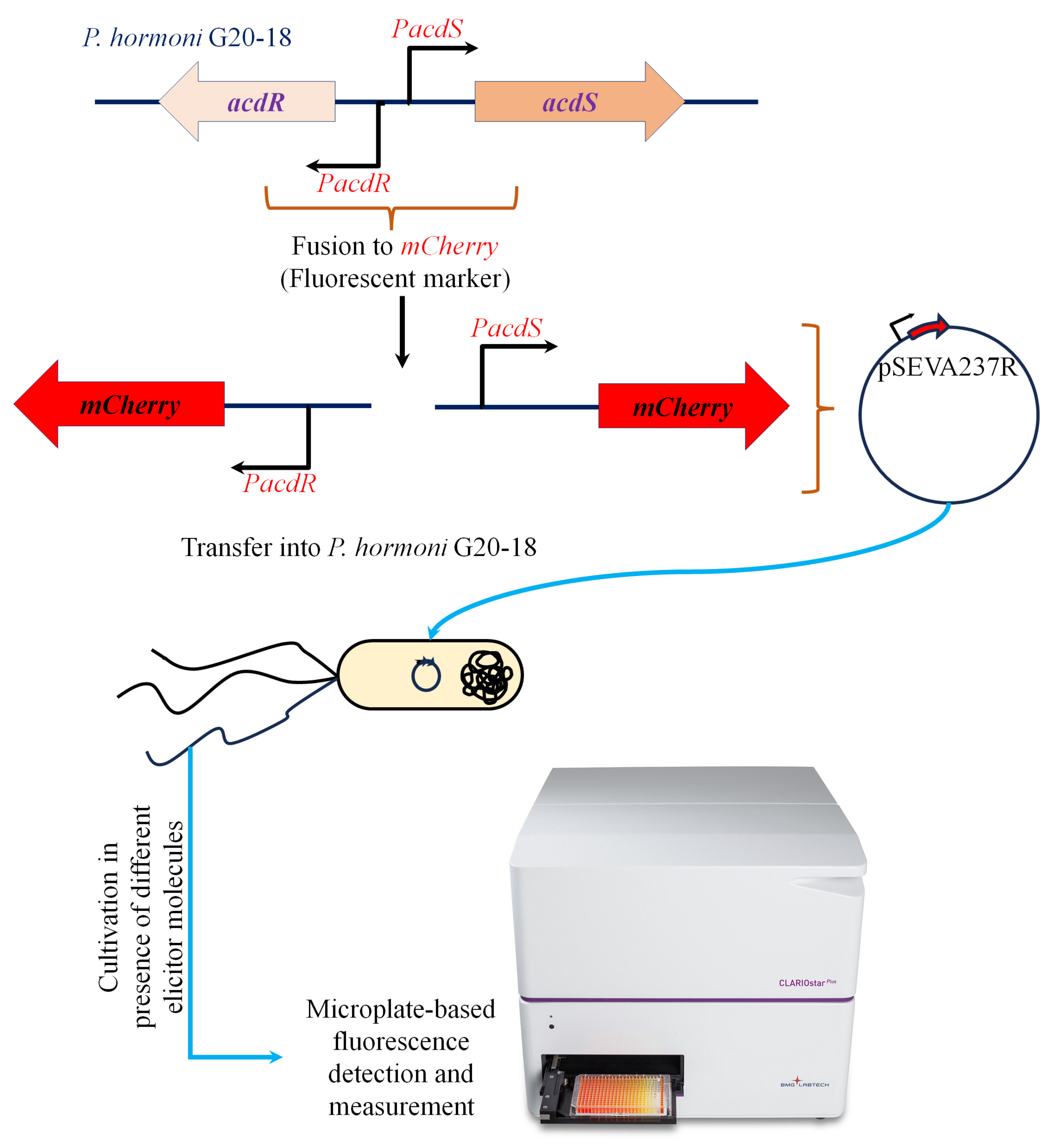
2.2. Construction of Promoter Fusions
2.3. Production and Characterization of Wheat Root Exudates
2.4. Activity of acdR and acdS Promoters under the Influence of Root Exudate Analogues
2.5. Wheat Seedling in Planta Studies
2.6. Statistical Analysis
3. Results and Discussion
3.1. Influence of Nitrogenous Compounds on acdS Promoter Activity
3.2. Influence of Amino Acids on acdS Promoter Activity
3.3. Influence of Sugars on acdS Promoter Activity
3.4. Influence of Indole-3-Acetic Acid (IAA) on acdS Promoter Activity
3.5. acdS Promoter Activity in Epiphytic vs. Endophytic Cells
3.6. Model for Regulation of acdS Promoter Activity in P. hormoni G20-18T
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esashi, Y. Ethylene and seed germination. In The Plant Hormone Ethylene; Matoo, A.K., Suttle, J.C., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 133–157. [Google Scholar]
- Ma, W.; Guinel, F.C.; Glick, B.R. Rhizobium leguminosarum biovar viciae 1-aminocyclopropane-1-carboxylate deaminase promotes nodulation of pea plants. Appl. Environ. Microbiol. 2003, 69, 4396–4402. [Google Scholar] [CrossRef]
- Jackson, M.B. Ethylene in root growth and development. In The Plant Hormone Ethylene; Matoo, A.K., Suttle, J.C., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 159–181. [Google Scholar]
- Grichko, V.P.; Glick, B.R. Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol. Biochem. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Checcucci, A.; Azzarello, E.; Bazzicalupo, M.; De Carlo, A.; Emiliani, G.; Mancuso, S.; Spini, G.; Viti, C.; Mengoni, A. Role and Regulation of ACC Deaminase Gene in Sinorhizobium meliloti: Is It a Symbiotic, Rhizospheric or Endophytic Gene? Front. Genet. 2017, 8, 6. [Google Scholar] [CrossRef]
- Glick, B.R.; Jacobson, C.B.; Schwarze, M.M.K.; Pasternak, J.J. 1-Aminocyclopropane-1-carboxylic acid deaminase mutants of the plant growth promoting rhizobacterium Pseudomonas putida GR12-2 do not stimulate canola root elongation. Can. J. Microbiol. 1994, 40, 911–915. [Google Scholar] [CrossRef]
- Cheng, Z.; Duncker, B.P.; McConkey, B.J.; Glick, B.R. Transcriptional regulation of ACC deaminase gene expression in Pseudomonas putida UW4. Can. J. Microbiol. 2008, 54, 128–136. [Google Scholar] [CrossRef]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [CrossRef]
- Uchiumi, T.; Ohwada, T.; Itakura, M.; Mitsui, H.; Nukui, N.; Dawadi, P.; Kaneko, T.; Tabata, S.; Yokoyama, T.; Tejima, K.; et al. Expression islands clustered on symbiosis island of Mesorhizobium loti genome. J. Bacteriol. 2004, 186, 2439–2448. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Soares, C.R.; McConkey, B.J.; Glick, B.R. New insights into 1-aminocyclopropane-1-carboxylate (ACC) deaminase phylogeny, evolution and ecological significance. PLoS ONE 2014, 9, e99168. [Google Scholar] [CrossRef] [PubMed]
- Prigent-Combaret, C.; Blaha, D.; Pothier, J.F.; Vial, L.; Poirier, M.A.; Wisniewski-Dyé, F.; Moënne-Loccoz, Y. Physical organization and phylogenetic analysis of acdR as leucine-responsive regulator of the 1-aminocyclopropane-1-carboxylate deaminase gene acdS in phytobeneficial Azospirillum lipoferum 4B and other Proteobacteria. FEMS Microbiol. Ecol. 2008, 65, 202–219. [Google Scholar] [CrossRef]
- Grichko, V.P.; Glick, B.R. Identification of DNA sequences that regulate the expression of the Enterobacter cloacae UW4 1-aminocyclopropane-1-carboxylic acid deaminase gene. Can. J. Microbiol. 2000, 46, 1159–1165. [Google Scholar] [CrossRef]
- Li, J.; Glick, B.R. Transcriptional regulation of the Enterobacter cloaceae UW41-aminocyclopropane-1-carboxylate (ACC) deaminase gene (AcdS). Can. J. Microbiol. 2001, 47, 259–267. [Google Scholar] [CrossRef]
- Ekimova, G.A.; Fedorov, D.N.; Doronina, N.V.; Khmelenina, V.N.; Mustakhimov, I.I. AcdR protein is an activator of transcription of 1-aminocyclopropane-1-carboxylate deaminase in Methylobacterium radiotolerans JCM 2831. Antonie Van Leeuwenhoek 2022, 115, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ovakim, D.H.; Charles, T.C.; Glick, B.R. An ACC deaminase minus mutant of Enterobacter cloacae UW4 no longer promotes root elongation. Curr. Microbiol. 2000, 41, 101–105. [Google Scholar] [CrossRef]
- Silva-Rocha, R.; Martínez-García, E.; Calles, B.; Chavarría, M.; Arce-Rodríguez, A.; de Las Heras, A.; Páez-Espino, A.D.; Durante-Rodríguez, G.; Kim, J. The Standard European Vector Architecture (SEVA): A coherent platform for the analysis and deployment of complex prokaryotic phenotypes. Nucleic Acids Res. 2013, 41, D666–D675. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, R.C.; Phippen, C.B.W.; Nielsen, K.F.; Olsson, S.; Stougaard, P. Biosynthesis of the antimicrobial cyclic lipopeptides nunamycin and nunapeptin by Pseudomonas fluorescens strain In5 is regulated by the LuxR-type transcriptional regulator NunF. Microbiol. Open 2017, 6, e00516. [Google Scholar] [CrossRef]
- Michelsen, C.F.; Watrous, J.; Glaring, M.A.; Kersten, R.; Koyama, N.; Dorrestein, P.C.; Stougaard, P. Nonribosomal peptides, key biocontrol components for Pseudomonas fluorescens In5, isolated from a Greenlandic suppressive soil. mBio 2015, 6, e00079. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y.; Razavi, B.S. Rhizosphere Size and Shape: Temporal Dynamics and Spatial Stationarity. Soil Biol. Biochem. 2019, 135, 343–360. [Google Scholar] [CrossRef]
- Jensen, M.; Poulsen, R.; Langebæk, R.; Jenssen, B.M.; Moe, J.; Ciesielski, T.M.; Dietz, R.; Sonne, C.; Madsen, J.; Hansen, M. The metabolome of pink-footed goose: Heavy metals and lipid metabolism. Environ. Res. 2023, 231 Pt 1, 116043. [Google Scholar] [CrossRef]
- Viant, M.R.; Ebbels, T.M.D.; Beger, R.D.; Ekman, D.R.; Epps, D.J.T.; Kamp, H.; Leonards, P.E.G.; Loizou, G.D.; MacRae, J.I.; van Ravenzwaay, B.; et al. Use cases, best practice and reporting standards for metabolomics in regulatory toxicology. Nat. Commun. 2019, 10, 3041. [Google Scholar] [CrossRef]
- Phillips, D.A.; Fox, T.C.; King, M.D.; Bhuvaneswari, T.V.; Teuber, L.R. Microbial products trigger amino acid exudation from plant roots. Plant Physiol. 2004, 136, 2887–2894. [Google Scholar] [CrossRef]
- Jalali, B.L.; Suryanarayana, D. Shift in the carbohydrate spectrum of root exudates of wheat in relation to its root-rot disease. Plant Soil. 1971, 34, 261–267. [Google Scholar] [CrossRef]
- Xiao, J.X.; Zheng, Y.; Tang, L. Effect of wheat and faba bean intercropping on root exudation of low molecular weight organic acids. J. Appl. Ecol. 2014, 25, 1739–1744. [Google Scholar]
- Zhang, A.-H.; Ma, W.-L.; Lei, F.-J.; An, N.-B.; Zhang, X.-X.; Liu, Z.-Q.; Zhang, L.-X. Research on chemotaxis response of Alternaria panax to amino acid of ginseng root exudates. China J. Chin. Mater. Medica 2017, 42, 2052–2057. [Google Scholar] [CrossRef]
- Li, J.; Lin, S.; Zhang, Q.; Zhang, Q.; Hu, W.; He, H. Fine-Root Traits of Allelopathic Rice at the Seedling Stage and Their Relationship with Allelopathic Potential. PeerJ 2019, 7, e7006. [Google Scholar] [CrossRef] [PubMed]
- Sebastiana, M.; Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Monteiro, F.; Figueiredo, A.; Maia, M.; Nascimento, R.; Silva, M.S.; Ferreira, A.N.; et al. Metabolomics and Transcriptomics to Decipher Molecular Mechanisms Underlying Ectomycorrhizal Root Colonization of an Oak Tree. Sci. Rep. 2021, 11, 8576. [Google Scholar] [CrossRef]
- Honma, M. Enzymatic determination of 1-aminocyclopropane-1-carboxylate deaminase. Agric. Biol. Chem. 1983, 47, 617–618. [Google Scholar]
- Viterbo, A.; Landau, U.; Kim, S.; Chernin, L.; Chet, I. Characterization of ACC deaminase from the biocontrol and plant growth-promoting agent Trichoderma asperellum T203. FEMS Microbiol. Lett. 2010, 305, 42–48. [Google Scholar] [CrossRef]
- Jacobson, C.B.; Pasternak, J.J.; Glick, B.R. Partial purification and characterization of 1-aminocyclopropane-1-carboxylate deaminase from the plant growth promoting rhizobacterium Pseudomonas putida GR12-2. Can. J. Microbiol. 2011, 40, 1019–1025. [Google Scholar] [CrossRef]
- Ziegler, C.A.; Freddolino, P.L. The leucine-responsive regulatory proteins/feast-famine regulatory proteins: An ancient and complex class of transcriptional regulators in bacteria and archaea. Critic. Rev. Biochem. Mol. Biol. 2021, 56, 373–400. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; He, H.; Dong, S.; Tang, L.; Yang, E.; Wang, W.; Zhang, B. The leucine-responsive regulatory protein SCAB_Lrp modulates thaxtomin biosynthesis, pathogenicity, and morphological development in Streptomyces scabies. Mol. Plant Pathol. 2023, 24, 167–178. [Google Scholar] [CrossRef]
- Ziegler, C.A.; Freddolino, P.L. Escherichia coli Leucine-Responsive Regulatory Protein Bridges DNA In Vivo and Tunably Dissociates in the Presence of Exogenous Leucine. mBio 2023, 14, e0269022. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.; Pascal, R.A.; Johnston, M.; Raines, R.; Dikshit, D.; Krantz, A.; Honma, M. Mechanistic studies on the pyridoxal phosphate enzyme 1-aminocyclopropane-1-carboxylate from Pseudomonas sp. Biochemistry 1981, 20, 7509–7519. [Google Scholar] [CrossRef] [PubMed]
- Honma, M. Chemically reactive sulfhydryl groups of 1-aminocyclopropane-1-carboxylate deaminase. Agric. Biol. Chem. 1985, 49, 567–571. [Google Scholar] [CrossRef]
- Bulusu, V.; Jayaraman, V.; Balaram, H. Metabolic fate of fumarate, a side product of the purine salvage pathway in the intraerythrocytic stages of Plasmodium falciparum. J. Biol. Chem. 2011, 286, 9236–9245. [Google Scholar] [CrossRef]
- Guest, J.R.; Green, J.; Irvine, A.S.; Spiro, S. The FNR modulon and FNR-regulated gene expression. In Regulation of Gene Expression in Escherichia coli; Lin, E.C.C., Lynch, A.S., Eds.; Champman and Hall: New York, NY, USA, 1996; pp. 317–342. [Google Scholar] [CrossRef]
- Lopes, L.D.; Wang, P.; Futrell, S.L.; Schachtman, D.P. Sugars and Jasmonic Acid Concentration in Root Exudates Affect Maize Rhizosphere Bacterial Communities. Appl. Environ. Microbiol. 2022, 88, e0097122. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Scher, F.M.; Laliberte, M.; Tipping, B. Emergence-promoting rhizobacteria: Description and implications for agricuture. In Iron, Siderophores, and Plant Diseases; Swinburne, T.R., Ed.; Plenum Press: New York, NY, USA, 1986; pp. 155–164. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Tafner, R.; Moreno, M.V.; Stenglein, S.A.; García de Salamone, I.E.; Nelson, L.M.; Novák, O.; Strnad, M.; van der Graaff, E.; Roitsch, T. Cytokinin production by Pseudomonas fluorescens G20-18 determines biocontrol activity against Pseudomonas syringae in Arabidopsis. Sci. Rep. 2016, 6, 23310. [Google Scholar] [CrossRef] [PubMed]
- Mekureyaw, M.F.; Pandey, C.; Hennessy, R.C.; Nicolaisen, M.H.; Liu, F.; Nybroe, O.; Roitsch, T. The cytokinin-producing plant beneficial bacterium Pseudomonas fluorescens G20-18 primes tomato (Solanum lycopersicum) for enhanced drought stress responses. J. Plant Physiol. 2022, 270, 153629. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for lowering of plant ethylene concentrations by plant growth promoting bacteria. J. Theoret. Biol. 1998, 190, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Yemelyanov, V.V.; Lastochkin, V.V.; Chirkova, T.V.; Lindberg, S.M.; Shishova, M.F. Indoleacetic Acid Levels in Wheat and Rice Seedlings under Oxygen Deficiency and Subsequent Reoxygenation. Biomolecules 2020, 10, 276. [Google Scholar] [CrossRef]
- Liu, W.H.; Chen, F.F.; Wang, C.E.; Fu, H.H.; Fang, X.Q.; Ye, J.R.; Shi, J.Y. Indole-3-Acetic Acid in Burkholderia pyrrocinia JK-SH007: Enzymatic Identification of the Indole-3-Acetamide Synthesis Pathway. Front. Microbiol. 2019, 10, 2559. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Formula | m/z * | Reference |
|---|---|---|---|
| Aspartic acid # | C4H7NO4 | 134.05 | This study |
| Valine # | C5H11NO2 | 118.08 | This study |
| Arginine # | C6H14N4O2 | 174.10 | This study |
| Isoleucine # | C6H13NO2 | 132.10 | This study |
| Alanine # | C3H7NO2 | 90.05 | [22] |
| Leucine # | C6H13NO2 | 132.10 | This study |
| Tryptophan | C11H12N2O2 | 205.09 | This study |
| Phenylalanine | C9H11NO2 | 166.08 | This study |
| Glucose # | C6H12O6 | 179.05 | This study |
| Fructose # | C6H12O6 | 179.05 | This study |
| Sucrose # | C12H22O11 | 341.10 | This study |
| Mannitol # | C6H14O6 | 221.04 | This study |
| L-Rhamnose # | C6H12O5 | 164.06 | [23] |
| α-Ketobutyric acid # | C4H6O3 | 102.09 | [24] |
| Fumaric acid # | C4H4O4 | 116.35 | This study |
| GABA | C4H9NO2 | 104.07 | This study |
| Malic acid | C4H6O5 | 133.01 | This study |
| Indole-3-acetic acid # | C10H9NO2 | 175.06 | This study |
| Salicylic acid | C7H6O3 | 137.02 | This study |
| Succinic acid | C4H6O4 | 101.02 | This study |
| Azelaic acid | C9H16O4 | 189.11 | This study |
| Trans-zeatin | C10H13N5O | 218.1 | This study |
| DIMBOA | C9H9NO5 | 210.04 | This study |
| DIBOA | C8H7NO4 | 182.04 | This study |
| Jasmonic acid | C12H18O3 | 211.13 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorty, A.M.; Ntana, F.; Hansen, M.; Stougaard, P. Plant-Root Exudate Analogues Influence Activity of the 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Gene in Pseudomonas hormoni G20-18T. Microorganisms 2023, 11, 2504. https://doi.org/10.3390/microorganisms11102504
Sorty AM, Ntana F, Hansen M, Stougaard P. Plant-Root Exudate Analogues Influence Activity of the 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Gene in Pseudomonas hormoni G20-18T. Microorganisms. 2023; 11(10):2504. https://doi.org/10.3390/microorganisms11102504
Chicago/Turabian StyleSorty, Ajay Madhusudan, Fani Ntana, Martin Hansen, and Peter Stougaard. 2023. "Plant-Root Exudate Analogues Influence Activity of the 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Gene in Pseudomonas hormoni G20-18T" Microorganisms 11, no. 10: 2504. https://doi.org/10.3390/microorganisms11102504