3.1. Carbon Source Use by Xanthomonas Strains
Carbon source use was analyzed for bacterial strains listed in
Table 1 with Biolog GN2 Microplate TM (Biolog Inc. Hayward, CA, USA) following the manufacturer instructions. Readings were made at 0 and 24 h post-inoculation (hpi) to detect the earliest metabolic response. The use of carbon sources that differentiate
Xanthomonas strains studied is presented in
Table 3.
Tween 40, N-acetyl-D-glucosamine, D-cellobiose, D-fructose, D-galactose, gentiobiose, α-D-glucose, maltose, D-mannose, D-psicose, D-trehalose, pyruvic acid methyl-ester, α-keto glutaric acid, succinic acid, bromosuccinic acid, L-glutamic acid, L-proline, L-serine, L-threonine, and glycerol were used by all strains tested. The compounds not metabolized by any of the strains were α-cyclodextrin, N-acetyl-D-galactosamine, adonitol, m-inositol, D-mannitol, β-methyl-D-glucoside, L-rhamnose, D-sorbitol, xylitol, citric acid, formic acid, D-galactonic acid lactone, D-galacturonic acid, D-glucosaminic acid, D-glucuronic acid, γ-hidroxybutyric acid, ρ-hydroxy phenylacetic acid, itaconic acid, quinic acid, sebacic acid, glucuronamide, L-histidine, L-leucine, L-ornithine, L-phenylalanine, L-pyroglutamic acid, D-serine, D,L-carnitine, γ-amino butyr acid, inosine, thymidine, phenyethyl-amine, and 2-aminoethanol. Compared with Xc, citrus strains used cis-aconitic acid and L-alanyl-glycine. Xc specifically used Tween 80, D-saccharic acid, and uridine. Among the citrus strains, Xcc A, A*, and Aw strains used sucrose. Xcc 306 was atypical compared with all other Xcc A strains in that no activity was detected for dextrin, L-fucose, lactulose, and α-keto butyric acid; meanwhile, Xcc A* Iran10 was the only strain that responded to uridine. Glycyl-L-aspartic acid, propionic acid, D-alanine, and L-alanine were used by A* and Aw but not by wide-host-range A strains. Xcc A* strains were the only one that responded to α-D-lactose, turanose, and L-aspartic acid. In addition, Xcc A*, as did Xec and Xc, used D-melobiose, α-hydroxybutiric acid, and D,L-lactic acid.
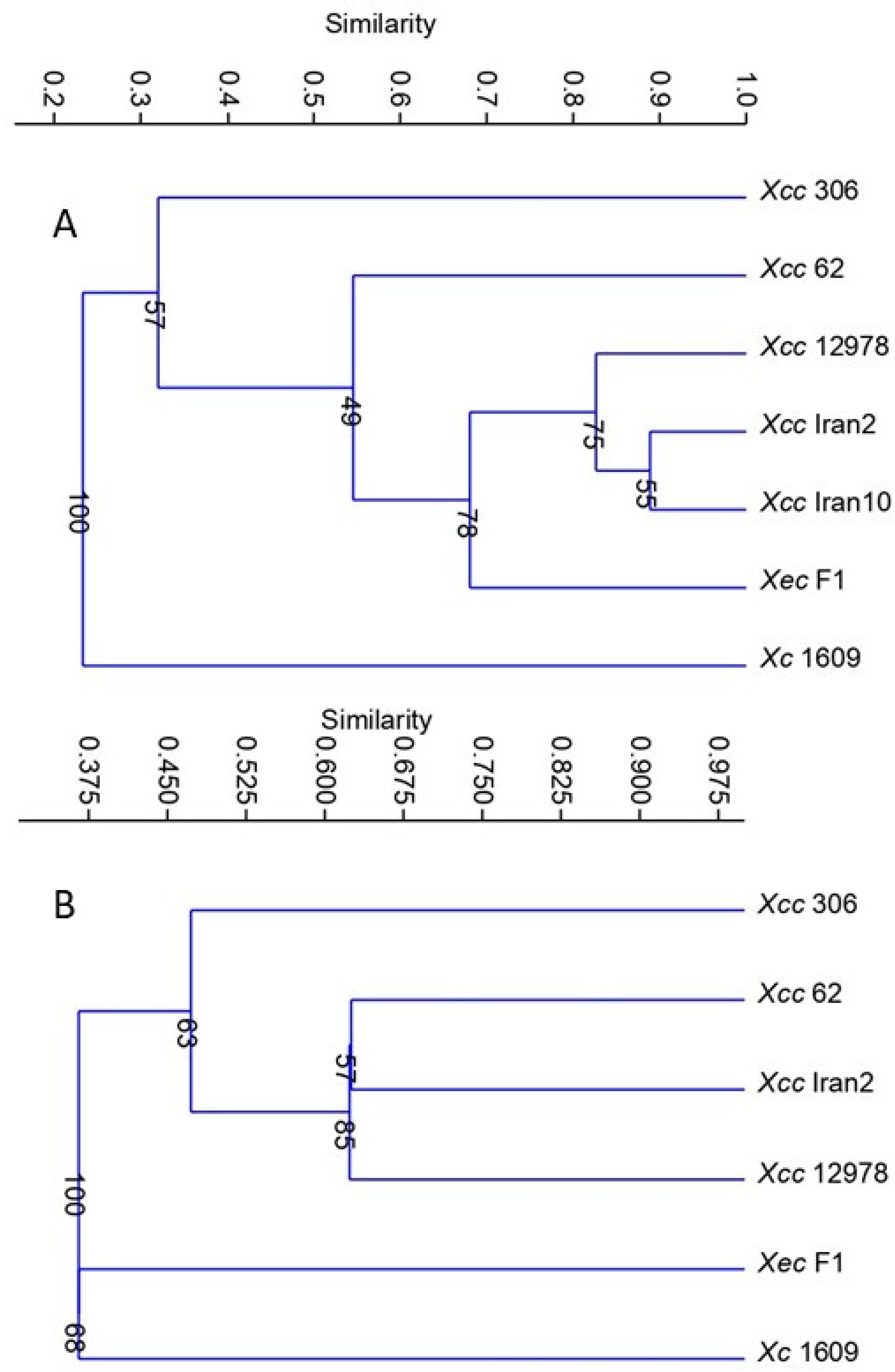
To study the overall relatedness of the metabolic response among the xanthomonads evaluated, cluster analysis was performed by transforming the data from carbon source use to binary form (uninformative carbon sources were dropped from the analysis). The analysis demonstrated that citrus strains were grouped in the same cluster and separated from
Xc. Moreover,
Xcc A strains were clustered according to the host range, i.e., separated from strains
Xcc, A*, A
w, and
Xec (
Figure 1A), and the two
Xcc strains showed their diversity.
Because a possible relationship between carbon source use and host range was elucidated, the putative role of chemical compounds in chemotaxis was studied later.
3.2. Chemotactic Response of Xanthomonas Strains to Carbon Compounds
To define the chemotactic profile of
Xanthomonas strains, a new chemotaxis assay, in which several compounds were concurrently tested with a large number of technical replicates, was developed. In this assay, the quantity of bacteria entering a pipette tip containing the carbon source was used to assess the chemotactic response independently of bacterial growth. This experimental approach has the same principle as the protocols described previously [
33,
36,
37]. This assay was validated with
D. dadantii 3937, and the chemotactic response obtained matched those previously reported: 10 mM cysteine was repellent, and 10 mM sodium citrate, 10 mM glucose, and 1 and 200 mM serine were attractants [
14,
34].
To determine the chemotactic responses of the
Xanthomonas studied, 19 compounds were tested (see
Table 4); from these chemicals, the metabolic response was determined using Biolog GN2 for 14 of them, and therefore, solely sodium citrate, xylose, arginine, cumaric acid, and cysteine’s metabolic responses were not considered.
All
Xanthomonas strains evaluated responded similarly to 10 mM cysteine as a repellent and 10 mM sucrose, 0.2% glycerol, and 200 mM serine as attractants (
Table 4).
Interestingly, the repellent cysteine has not been detected in the phloem sap of most
Citrus spp. [
38], and sucrose was previously reported as an attractant for other
Xanthomonas spp. [
36,
39].
Xc differed from citrus strains in that 10 mM alanine and 10 mM leucine acted as repellents and 10 mM glucuronic acid as an attractant. The responses that differentiated
Xcc strains from
Xec and
Xc were 150 mM leucine and 0.2% mannitol as attractants for
Xcc strains (repellent for
Xec and no response for
Xc) and 10 mM xylose and 10 mM serine as repellents for
Xec and
Xc, while no response was observed for
Xcc strains. In addition, among citrus pathogenic strains,
Xec was the sole strain showing a repellent response toward fructose and glucose, two well-known carbon sources for bacteria; 200 mM alanine did not show any chemotactic effect in
Xec, while it was an attractant for the
Xcc strains tested. As previously reported for
Ralstonia solanacearum strains [
40], chemotactic responses varied within
Xcc strains;
Xcc 306 and
Xcc 62 were the only strains attracted to 10 mM galacturonic acid, along with
Xec.
Xcc 62 was the only
Xcc A strain attracted to 10 mM arginine, and
Xcc 306 was the only showing no response to 100 mM arginine or being attracted by leucine at 10 mM. Cluster analysis based on chemoattraction grouped
Xcc A strains with the narrow-host-range strains
Xcc A* Iran2 and
Xcc A
w 12879 and separated them from
Xec and
Xc. Within the
Xcc subgroup,
Xcc 62 was more closely related to
Xcc A* Iran2 and
Xcc A
w 12879 than to
Xcc 306 (
Figure 1B). Chemotactic responses were more similar for narrow-host-range strains, while the wide-host-range strains responses were variable.
3.3. Identification of MCPs in Xanthomonas Species Used in the Study
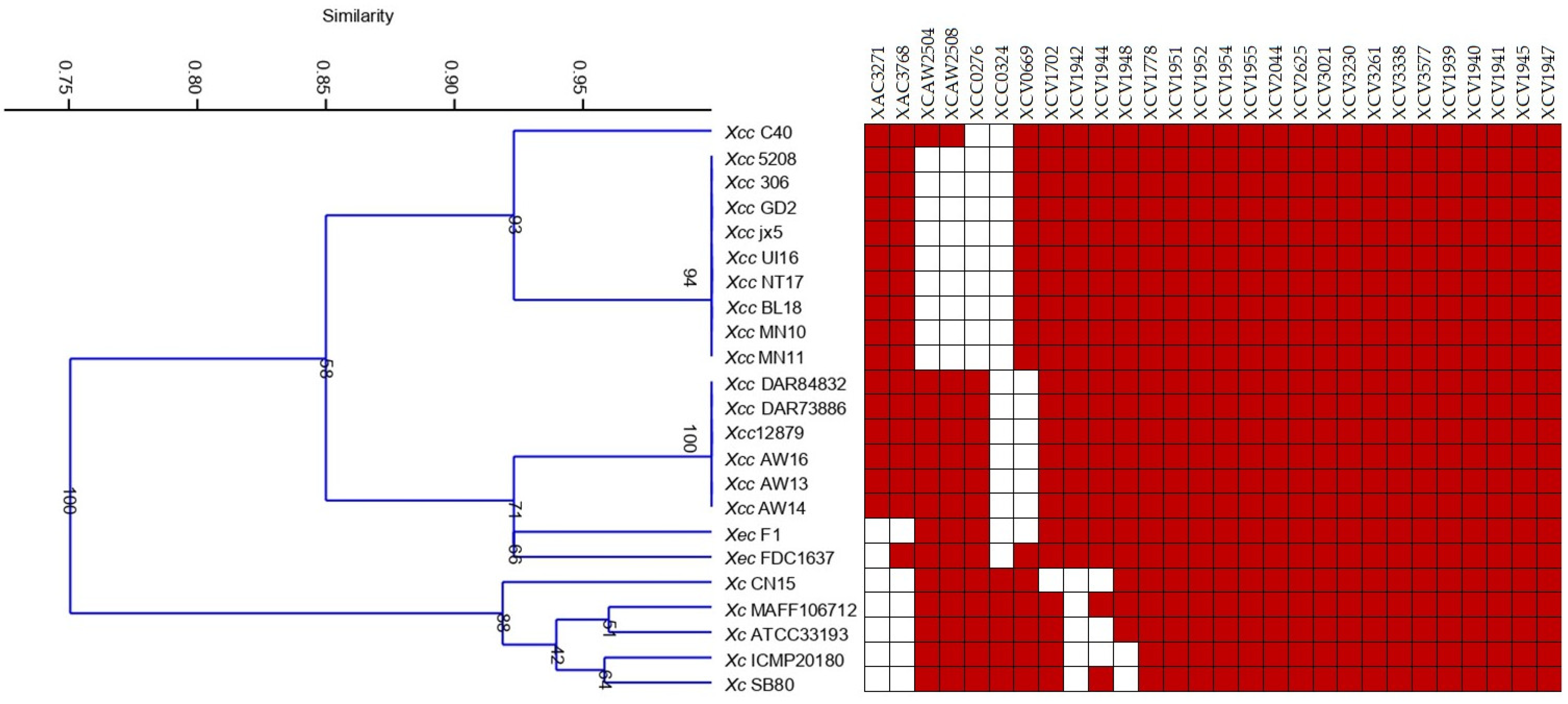
The analysis of the complete genomes of different
Xanthomonas species, pathovars, and pathotypes revealed variants in their MCP profiles. Although 28 different MCPs were found in the genome sequences, the number of MCPs varied from 24 in most of the type A
Xcc strains to 26 in all A*/A
w Xcc and
Xec FDC1637 strains. An MCP pattern composed of 18 genes was shared by all genomes analyzed; meanwhile, citrus-associated and brassica-associated strains shared 22 and 24 MCPs, respectively. Among those common MCPs, XCV1942, XAC3768, and XAC3271 were only present in citrus-associated xanthomonads and XCC0324 was only found in brassica-associated ones (
Figure 2). Results also showed that the MCP content differed among citrus xanthomonads; thereby, XAC3271 was only identified in
Xcc, but it was not found in
Xec, and although XCAW2504 and XCAW2508 were detected in all
Xcc A*/A
w strains, they were found in just one
Xcc A strain and were not identified in any
Xec.
Cluster analysis of the binary data obtained from MCP analysis revealed major groups according to pathotype and
Xanthomonas spp. (
Figure 2). One cluster included all
Xcc type A strains separated from A
w/A* that grouped together with
Xec and more separated from
Xc (
Figure 2).
PCR using primers previously described [
19] in addition to those for XCAW2504 and XCAW2508 results confirmed findings from the genomic analysis (
Table 5). XCV1942, XAC3768, and XAC3271 were identified in citrus strains but not in
Xc 1609, and XCC0324 was only found in
Xc 1609. In addition, some other MCPs were universally distributed in all the strains, in line with genomic results. As well, either A* or A
w Xcc strains showed the same MCP/PCR profile; meanwhile, variability among
Xcc A strains was found in the MCP content (
Figure 2,
Table 5).
The difference in the presence of specific MCPs was related to the host (citrus vs. crucifer) and the citrus pathogenic species (Xec vs. Xcc strains); moreover, the minor differences revealed within the Xcc A strains were in concordance with their different chemotactic responses to carbon compounds.
3.4. Xanthomonas Strains Are Attracted by Leaf Apoplastic Fluids
To confirm the role of chemotaxis at an early stage of leaf infection, chemotaxis of the different strains was assessed in response to apoplastic fluids from sweet orange (
C. sinensis) var. ‘Valencia Late’, Mexican lime (
Citrus aurantifolia), and Chinese cabbage (
Brassica pekinensis) var. Kasumi. Our results showed that all apoplastic fluids act as chemoattractants (
Table 6). Both cabbage and citrus apoplastic fluids were attractive for all
Xanthomonas strains.
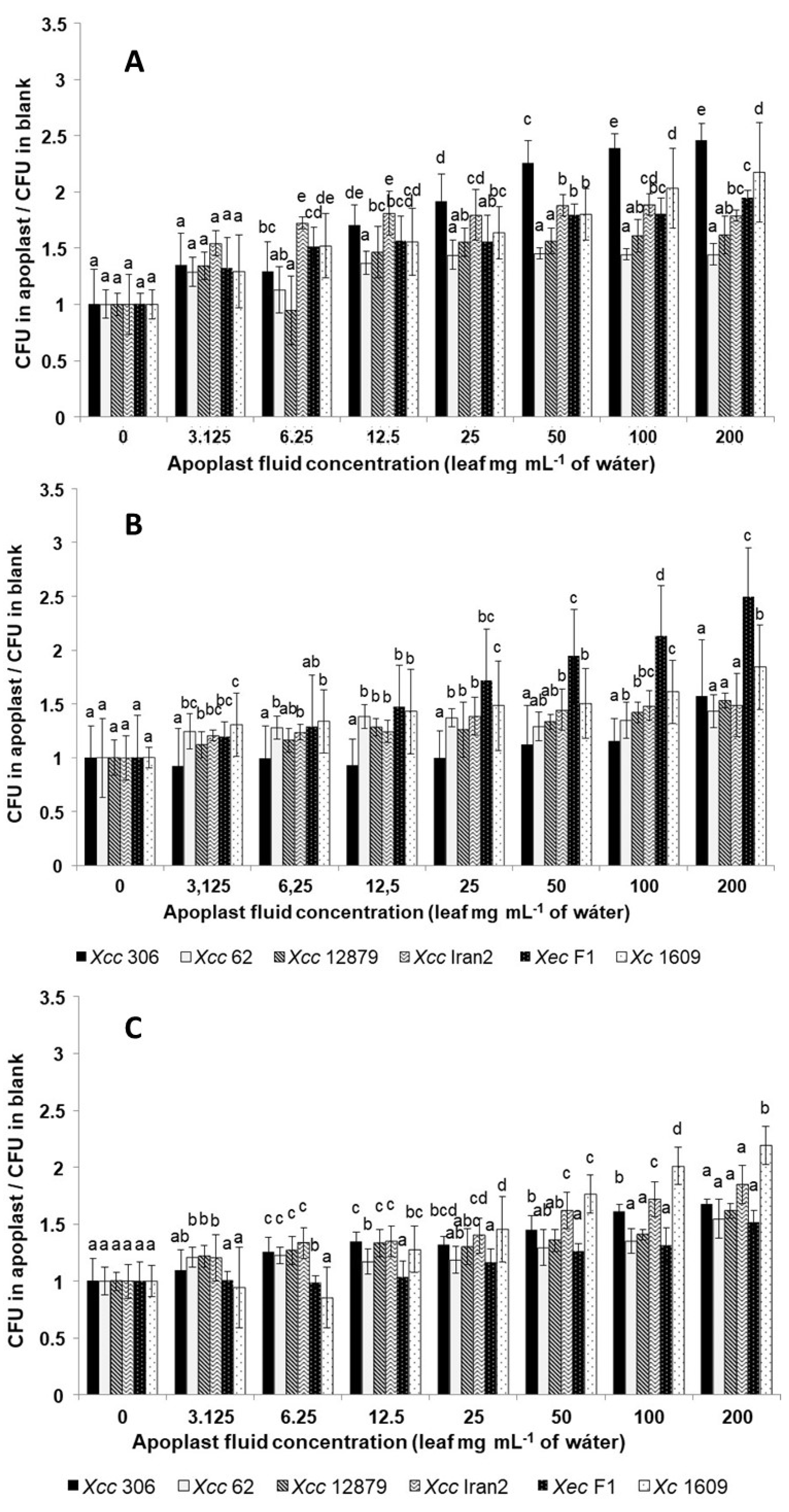
Nevertheless, the response differed among strains:
Xcc A 306 was more responsive to sweet orange,
Xec F1 to Mexican lime, and
Xc 1609 to Chinese cabbage, indicating a clear difference in the response between citrus and crucifer strains (
Figure 3). Moreover, although these strains weakly responded to the lowest concentrations of apoplast fluids from these species (
Table 6), their chemoattractive response increased markedly with the apoplastic fluid concentration. The same occurred for the interaction between
Xcc A 306 and Mexican lime (
Figure 3).
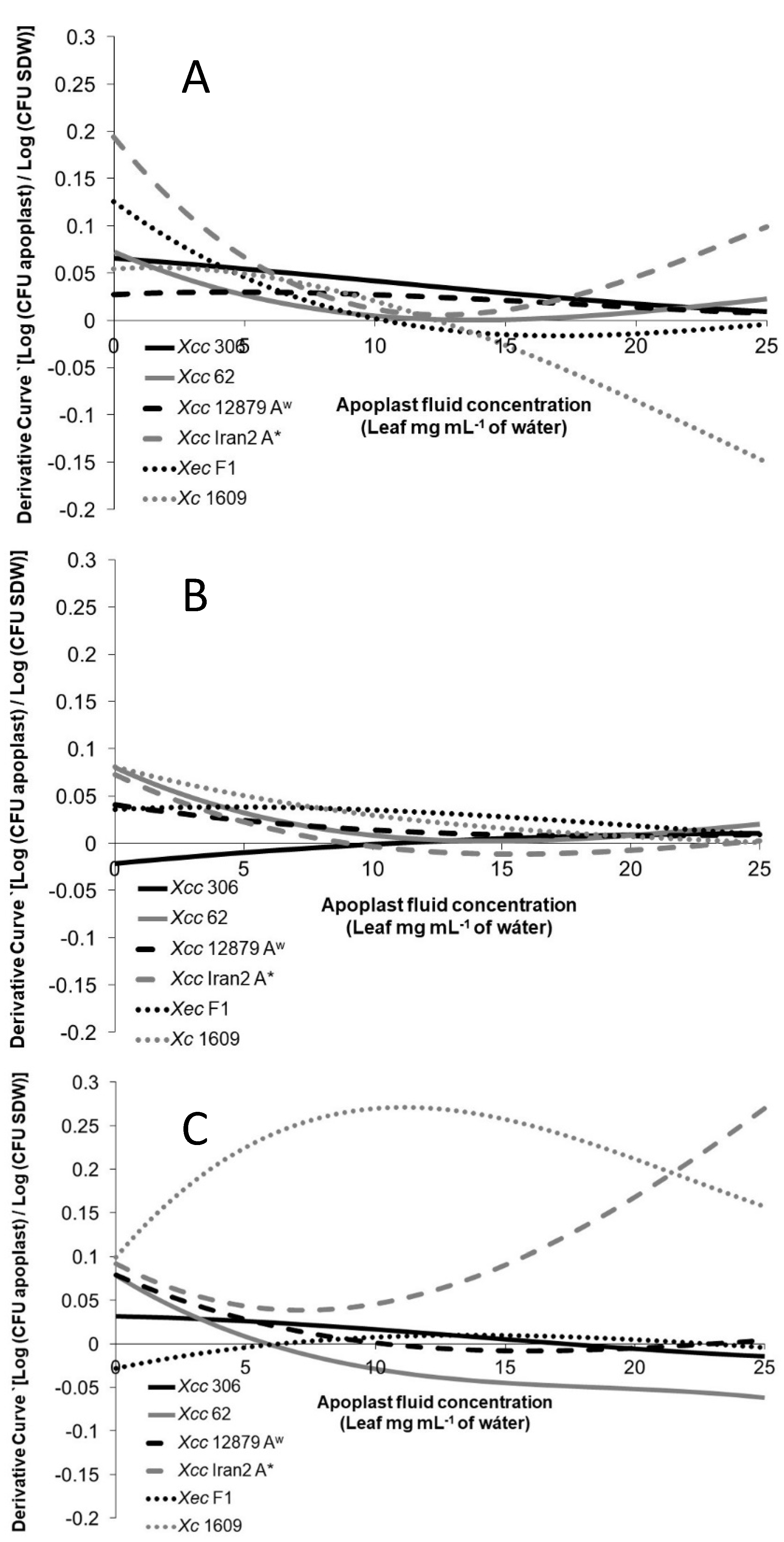
To evaluate more precisely the differences among the strains on the different hosts, the variation of the chemotactic response related to the apoplast concentration increase was analyzed. The chemotactic derivative curves in
Figure 4 show how the chemotactic response changed as the apoplastic fluid concentration increased.
The chemotactic responses of the
Xanthomonas strains tested toward citrus apoplastic fluids (
Figure 4A,B) showed higher response changes at low concentrations for most of the strains. Usually, the chemoattractive response diminished or even declined as the apoplastic fluid concentration increased. However, it is important to note that this reduction in the chemoattractive response does not mean a negative response (chemorepellent) but fewer bacterial cells entering the tip with apoplastic concentration increments.
Citrus pathogenic strains’ response toward the Chinese cabbage apoplastic fluid was constant or even negative when the concentration increased, with the exception of
Xcc Iran2 A* (
Figure 4C). The same behavior was observed in
Xc 1609 toward most citrus apoplastic fluids. This result suggests that on a non-host-plant leaf surface, the xanthomonad chemotactic response would not be as efficient as the pathogen approaches the stomata.
The orange leaf apoplastic fluid produced the most variable response among the
Xanthomonas strains tested (
Figure 4A). The highest variation of the response associated with the concentration was observed for
Xcc 306, presenting
Xec F1 and
Xcc Iran2, A* an intermediate phenotype; meanwhile, the lowest variation was found in
Xcc 62 and
Xcc A
w 12879. Moreover, the response of
Xcc Iran2 A* showed a reduction in the variation at concentrations over 6.25 mg mL
−1.
No differences in the chemotactic response toward Mexican lime was observed among
Xcc 62,
Xcc A
w 12879, and
Xcc Iran2 A* (
Figure 4B). However, less reaction was observed for
Xcc 306, although the response increased with the apoplastic fluid concentration.
Xc 1609 was highly responsive toward the Chinese cabbage apoplastic fluid compared with citrus
Xanthomonas (
Figure 4C) and less reactive to citrus apoplastic fluids (
Figure 4A,B).


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}