

TORC1 Signaling in Fungi: From Yeasts to Filamentous Fungi

Abstract
:1. Introduction
2. Operational Mode of TORC1 in Yeast Fungi
3. Upstream Signals of TORC1 in Yeast Fungi
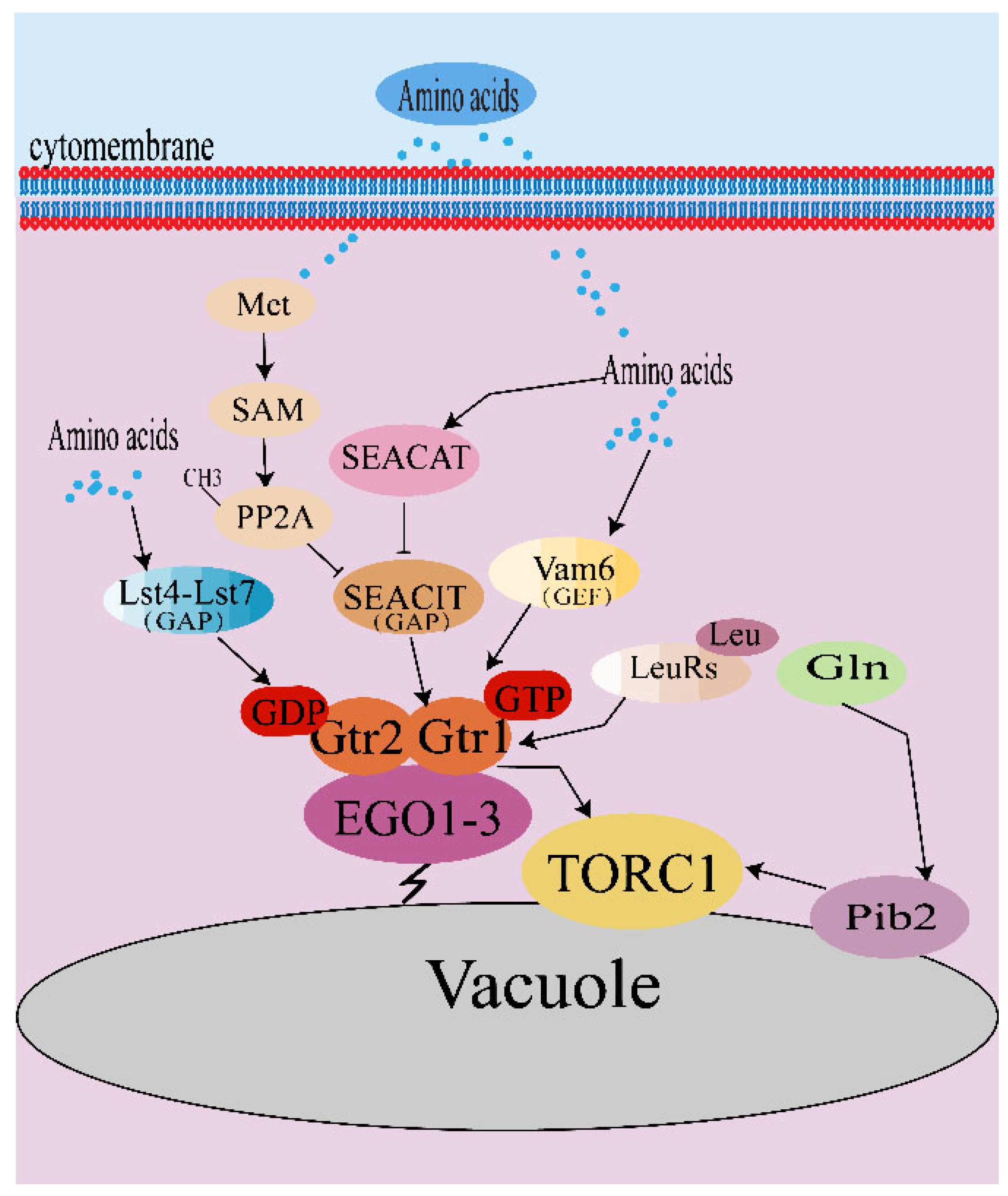
3.1. Amino Acid Signaling of TORC1 in Yeast Fungi
3.2. Carbon Signaling of TORC1 in Yeast Fungi
3.3. Phosphate Signaling of TORC1 in Yeast Fungi
4. Effects on Fungal Growth and Metabolism
4.1. Modulation of Protein Synthesis by TORC1
4.2. Modulation of Metabolic Programs by TORC1
4.3. Modulation of Macroautophagy by TORC1
4.4. Modulation of Stress Response by TORC1
4.5. Modulation of Pathogenicity and Virulence by TORC1
4.6. Modulation of Ubiquitination by TORC1
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muñoz-Muñoz, P.L.A.; Mares-Alejandre, R.E.; Meléndez-López, S.G.; Ramos-Ibarra, M.A. Bioinformatic Analysis of Two TOR (Target of Rapamycin)-Like Proteins Encoded by Entamoeba histolytica Revealed Structural Similarities with Functional Homologs. Genes 2021, 12, 1139. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, O.; Strilbytska, O.; Piskovatska, V.; Storey, K.B.; Koliada, A.; Vaiserman, A. The role of the TOR pathway in mediating the link between nutrition and longevity. Mech. Ageing Dev. 2017, 164, 127–138. [Google Scholar] [CrossRef]
- Loewith, R.; Hall, M.N. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Acosta-Zaldivar, M.; Flanagan, P.R.; Liu, N.N.; Jani, N.; Fierro, J.F.; Andrés, M.T.; Moran, G.P.; Köhler, J.R. Stress- and metabolic responses of Candida albicans require Tor1 kinase N-terminal HEAT repeats. PLoS Pathog. 2022, 18, e1010089. [Google Scholar] [CrossRef] [PubMed]
- Mallén-Ponce, M.J.; Pérez-Pérez, M.E.; Crespo, J.L. Deciphering the function and evolution of the target of rapamycin signaling pathway in microalgae. J. Exp. Bot. 2022, 73, 6993–7005. [Google Scholar] [CrossRef]
- Davie, E.; Petersen, J. Environmental control of cell size at division. Curr. Opin. Cell Biol. 2012, 24, 838–844. [Google Scholar] [CrossRef]
- Wullschleger, S.; Loewith, R.; Oppliger, W.; Hall, M.N. Molecular organization of target of rapamycin complex 2. J. Biol. Chem. 2005, 280, 30697–30704. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhu, T.; Song, Y.; Feng, L.; Kear, P.J.; Riseh, R.S.; Sitohy, M.; Datla, R.; Ren, M. Salicylic acid fights against Fusarium wilt by inhibiting target of rapamycin signaling pathway in Fusarium oxysporum. J. Adv. Res. 2022, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Hall, M.N. Nutrient sensing and TOR signaling in yeast and mammals. EMBO J. 2017, 36, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, R.T. Identification of TOR signaling complexes: More TORC for the cell growth engine. Cell 2002, 111, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Dobrenel, T.; Caldana, C.; Hanson, J.; Robaglia, C.; Vincentz, M.; Veit, B.; Meyer, C. TOR Signaling and Nutrient Sensing. Annu. Rev. Plant Biol. 2016, 67, 261–285. [Google Scholar] [CrossRef] [PubMed]
- Pancha, I.; Chokshi, K.; Tanaka, K.; Imamura, S. Microalgal Target of Rapamycin (TOR): A Central Regulatory Hub for Growth, Stress Response and Biomass Production. Plant Cell Physiol. 2020, 61, 675–684. [Google Scholar] [CrossRef]
- Powis, K.; Zhang, T.; Panchaud, N.; Wang, R.; De Virgilio, C.; Ding, J. Crystal structure of the Ego1-Ego2-Ego3 complex and its role in promoting Rag GTPase-dependent TORC1 signaling. Cell Res. 2015, 25, 1043–1059. [Google Scholar] [CrossRef] [Green Version]
- Powis, K.; De Virgilio, C. Conserved regulators of Rag GTPases orchestrate amino acid-dependent TORC1 signaling. Cell Discov. 2016, 2, 15049. [Google Scholar] [CrossRef] [Green Version]
- Rashida, Z.; Srinivasan, R.; Cyanam, M.; Laxman, S. Kog1/Raptor mediates metabolic rewiring during nutrient limitation by controlling SNF1/AMPK activity. Sci. Adv. 2021, 7, eabe5544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Péli-Gulli, M.P.; Yang, H.; De Virgilio, C.; Ding, J. Ego3 functions as a homodimer to mediate the interaction between Gtr1-Gtr2 and Ego1 in the ego complex to activate TORC1. Structure 2012, 20, 2151–2160. [Google Scholar] [CrossRef] [Green Version]
- Condon, K.J.; Sabatini, D.M. Nutrient regulation of mTORC1 at a glance. J. Cell Sci. 2019, 132, jcs222570. [Google Scholar] [CrossRef]
- Morozumi, Y.; Shiozaki, K. Conserved and Divergent Mechanisms That Control TORC1 in Yeasts and Mammals. Genes 2021, 12, 88. [Google Scholar] [CrossRef]
- Dokudovskaya, S.; Rout, M.P. SEA you later alli-GATOR--a dynamic regulator of the TORC1 stress response pathway. J. Cell Sci. 2015, 128, 2219–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Sofyantoro, F.; Tai, Y.T.; Chia, K.H.; Shiozaki, K. Tripartite suppression of fission yeast TORC1 signaling by the GATOR1-Sea3 complex, the TSC complex, and Gcn2 kinase. eLife 2021, 10, e60969. [Google Scholar] [CrossRef] [PubMed]
- Péli-Gulli, M.P.; Sardu, A.; Panchaud, N.; Raucci, S.; De Virgilio, C. Amino Acids Stimulate TORC1 through Lst4-Lst7, a GTPase-Activating Protein Complex for the Rag Family GTPase Gtr2. Cell Rep. 2015, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binda, M.; Péli-Gulli, M.P.; Bonfils, G.; Panchaud, N.; Urban, J.; Sturgill, T.W.; Loewith, R.; De Virgilio, C. The Vam6 GEF controls TORC1 by activating the EGO complex. Mol. Cell 2009, 35, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, R.L.; Lu, E.; Luo, X.; Capaldi, A.P. Ait1 regulates TORC1 signaling and localization in budding yeast. elife 2022, 11, e68773. [Google Scholar] [CrossRef]
- Varlakhanova, N.V.; Tornabene, B.A.; Ford, M.G.J. Ivy1 is a negative regulator of Gtr-dependent TORC1 activation. J. Cell Sci. 2018, 131, jcs218305. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, G.; Zhang, Y.; Dayhoff-Brannigan, M.; Diny, N.L.; Zhao, M.; He, G.; Sing, C.N.; Metz, K.A.; Stolp, Z.D.; et al. Whi2 is a conserved negative regulator of TORC1 in response to low amino acids. PLoS Genet. 2018, 14, e1007592. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Guo, S.; Yan, G.; Yuan, W.; Zheng, Y.; Jiang, Y. Ubiquitin regulates TORC1 in yeast Saccharomyces cerevisiae. Mol. Microbiol. 2016, 100, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Otsubo, Y.; Kamada, Y.; Yamashita, A. Novel Links between TORC1 and Traditional Non-Coding RNA, tRNA. Genes 2020, 11, 956. [Google Scholar] [CrossRef]
- Tessler, F.N.; Middleton, W.D.; Grant, E.G. Thyroid Imaging Reporting and Data System (TI-RADS): A User’s Guide. Radiology 2018, 287, 29–36. [Google Scholar] [CrossRef]
- Stracka, D.; Jozefczuk, S.; Rudroff, F.; Sauer, U.; Hall, M.N. Nitrogen source activates TOR (target of rapamycin) complex 1 via glutamine and independently of Gtr/Rag proteins. J. Biol. Chem. 2014, 289, 25010–25020. [Google Scholar] [CrossRef] [Green Version]
- Michel, A.H.; Hatakeyama, R.; Kimmig, P.; Arter, M.; Peter, M.; Matos, J.; De Virgilio, C.; Kornmann, B. Functional mapping of yeast genomes by saturated transposition. eLife 2017, 6, e23570. [Google Scholar] [CrossRef]
- Tanigawa, M.; Yamamoto, K.; Nagatoishi, S.; Nagata, K.; Noshiro, D.; Noda, N.N.; Tsumoto, K.; Maeda, T. A glutamine sensor that directly activates TORC1. Commun. Biol. 2021, 4, 1093. [Google Scholar] [CrossRef]
- Zhang, W.; Du, G.; Zhou, J.; Chen, J. Regulation of Sensing, Transportation, and Catabolism of Nitrogen Sources in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2018, 82, e00040-17. [Google Scholar] [CrossRef] [Green Version]
- Alfatah, M.; Wong, J.H.; Krishnan, V.G.; Lee, Y.C.; Sin, Q.F.; Goh, C.J.H.; Kong, K.W.; Lee, W.T.; Lewis, J.; Hoon, S.; et al. TORC1 regulates the transcriptional response to glucose and developmental cycle via the Tap42-Sit4-Rrd1/2 pathway in Saccharomyces cerevisiae. BMC Biol. 2021, 19, 95. [Google Scholar] [CrossRef]
- Clement, S.T.; Dixit, G.; Dohlman, H.G. Regulation of yeast G protein signaling by the kinases that activate the AMPK homolog Snf1. Sci. Signal. 2013, 6, ra78. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, X.; Jiang, H.Y.; Song, Z.M.; Lin, X.; Hu, X.P.; Li, C.F. Protein kinases Elm1 and Sak1 of Saccharomyces cerevisiae exerted different functions under high-glucose and heat shock stresses. Appl. Microbiol. Biotechnol. 2022, 106, 2029–2042. [Google Scholar] [CrossRef]
- Coccetti, P.; Nicastro, R.; Tripodi, F. Conventional and emerging roles of the energy sensor Snf1/AMPK in Saccharomyces cerevisiae. Microb. Cell 2018, 5, 482–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umekawa, M.; Ujihara, M.; Nakai, D.; Takematsu, H.; Wakayama, M. Ecm33 is a novel factor involved in efficient glucose uptake for nutrition-responsive TORC1 signaling in yeast. FEBS Lett. 2017, 591, 3721–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dechant, R.; Saad, S.; Ibáñez, A.J.; Peter, M. Cytosolic pH regulates cell growth through distinct GTPases, Arf1 and Gtr1, to promote Ras/PKA and TORC1 activity. Mol. Cell. 2014, 55, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Zinjarde, S.S.; Kale, B.V.; Vishwasrao, P.V.; Kumar, A.R. Morphogenetic behavior of tropical marine yeast Yarrowia lipolytica in response to hydrophobic substrates. J. Microbiol. Biotechnol. 2008, 18, 1522–1528. [Google Scholar] [PubMed]
- Ruiz-Herrera, J.; Sentandreu, R. Different effectors of dimorphism in Yarrowia lipolytica. Arch. Microbiol. 2002, 178, 477–483. [Google Scholar] [CrossRef]
- Liu, N.N.; Flanagan, P.R.; Zeng, J.; Jani, N.M.; Cardenas, M.E.; Moran, G.P.; Köhler, J.R. Phosphate is the third nutrient monitored by TOR in Candida albicans and provides a target for fungal-specific indirect TOR inhibition. Proc. Natl. Acad. Sci. USA 2017, 114, 6346–6351. [Google Scholar] [CrossRef] [Green Version]
- Mouillon, J.M.; Persson, B.L. New aspects on phosphate sensing and signalling in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomar, P.; Sinha, H. Conservation of PHO pathway in ascomycetes and the role of Pho84. J. Biosci. 2014, 39, 525–536. [Google Scholar] [CrossRef]
- Urban, J.; Soulard, A.; Huber, A.; Lippman, S.; Mukhopadhyay, D.; Deloche, O.; Wanke, V.; Anrather, D.; Ammerer, G.; Riezman, H.; et al. Sch9 is a major target of TORC1 in Saccharomyces cerevisiae. Mol. Cell. 2007, 26, 663–674. [Google Scholar] [CrossRef]
- Lageix, S.; Zhang, J.; Rothenburg, S.; Hinnebusch, A.G. Interaction between the tRNA-binding and C-terminal domains of Yeast Gcn2 regulates kinase activity in vivo. PLoS Genet. 2015, 11, e1004991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallinetti, J.; Harputlugil, E.; Mitchell, J.R. Amino acid sensing in dietary-restriction-mediated longevity: Roles of signal-transducing kinases GCN2 and TOR. Biochem. J. 2013, 449, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarumoto, Y.; Kanoh, J.; Ishikawa, F. Receptor for activated C-kinase (RACK1) homolog Cpc2 facilitates the general amino acid control response through Gcn2 kinase in fission yeast. J. Biol. Chem. 2013, 288, 19260–19268. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.; French, S.L.; Tekotte, H.; Yerlikaya, S.; Stahl, M.; Perepelkina, M.P.; Tyers, M.; Rougemont, J.; Beyer, A.L.; Loewith, R. Sch9 regulates ribosome biogenesis via Stb3, Dot6 and Tod6 and the histone deacetylase complex RPD3L. EMBO J. 2011, 30, 3052–3064. [Google Scholar] [CrossRef]
- González, A.; Shimobayashi, M.; Eisenberg, T.; Merle, D.A.; Pendl, T.; Hall, M.N.; Moustafa, T. TORC1 promotes phosphorylation of ribosomal protein S6 via the AGC kinase Ypk3 in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0120250. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, V.; Costa, V. Unraveling the role of the Target of Rapamycin signaling in sphingolipid metabolism. Prog. Lipid. Res. 2016, 61, 109–133. [Google Scholar] [CrossRef] [PubMed]
- Takahara, T.; Maeda, T. Evolutionarily conserved regulation of TOR signalling. J. Biochem. 2013, 154, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leśniewska, E.; Boguta, M. Novel layers of RNA polymerase III control affecting tRNA gene transcription in eukaryotes. Open Biol. 2017, 7, 170001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, A.; Otsubo, Y.; Yamashita, A.; Sato, T.; Yamamoto, M.; Tamanoi, F. Psk1, an AGC kinase family member in fission yeast, is directly phosphorylated and controlled by TORC1 and functions as S6 kinase. J. Cell Sci. 2012, 125, 5840–5849. [Google Scholar] [CrossRef] [Green Version]
- Ljungdahl, P.O. Amino-acid-induced signalling via the SPS-sensing pathway in yeast. Biochem. Soc. Trans. 2009, 37, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Andréasson, C.; Ljungdahl, P.O. Receptor-mediated endoproteolytic activation of two transcription factors in yeast. Genes Dev. 2002, 16, 3158–3172. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Mukai, Y. Yeast Cyc8p and Tup1p proteins function as coactivators for transcription of Stp1/2p-dependent amino acid transporter genes. Biochem. Biophys. Res. Commun. 2015, 468, 32–38. [Google Scholar] [CrossRef]
- Lleixà, J.; Martín, V.; Giorello, F.; Portillo, M.C.; Carrau, F.; Beltran, G.; Mas, A. Analysis of the NCR Mechanisms in Hanseniaspora vineae and Saccharomyces cerevisiae During Winemaking. Front. Genet. 2018, 9, 747. [Google Scholar] [CrossRef] [Green Version]
- Shamji, A.F.; Kuruvilla, F.G.; Schreiber, S.L. Partitioning the transcriptional program induced by rapamycin among the effectors of the Tor proteins. Curr. Biol. 2000, 10, 1574–1581. [Google Scholar] [CrossRef]
- Lu, J.Y.; Lin, Y.Y.; Sheu, J.C.; Wu, J.T.; Lee, F.J.; Chen, Y.; Lin, M.I.; Chiang, F.T.; Tai, T.Y.; Berger, S.L.; et al. Acetylation of yeast AMPK controls intrinsic aging independently of caloric restriction. Cell 2011, 146, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Zou, H.; Fu, J.; Zhou, J.; Du, G.; Chen, J. Metabolic engineering of the regulators in nitrogen catabolite repression to reduce the production of ethyl carbamate in a model rice wine system. Appl. Environ. Microbiol. 2014, 80, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Hahn, S.; Young, E.T. Transcriptional regulation in Saccharomyces cerevisiae: Transcription factor regulation and function, mechanisms of initiation, and roles of activators and coactivators. Genetics 2011, 189, 705–736. [Google Scholar] [CrossRef] [Green Version]
- Laor, D.; Cohen, A.; Kupiec, M.; Weisman, R. TORC1 Regulates Developmental Responses to Nitrogen Stress via Regulation of the GATA Transcription Factor Gaf1. mBio 2015, 6, e00959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swinnen, E.; Wilms, T.; Idkowiak-Baldys, J.; Smets, B.; De Snijder, P.; Accardo, S.; Ghillebert, R.; Thevissen, K.; Cammue, B.; De Vos, D.; et al. The protein kinase Sch9 is a key regulator of sphingolipid metabolism in Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 196–211. [Google Scholar] [CrossRef]
- Noda, T. Regulation of Autophagy through TORC1 and mTORC1. Biomolecules 2017, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Kohda, T.A.; Tanaka, K.; Konomi, M.; Sato, M.; Osumi, M.; Yamamoto, M. Fission yeast autophagy induced by nitrogen starvation generates a nitrogen source that drives adaptation processes. Genes Cells 2007, 12, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Fabrizio, P.; Hu, J.; Ge, H.; Cheng, C.; Li, L.; Longo, V.D. Life span extension by calorie restriction depends on Rim15 and transcription factors downstream of Ras/PKA, Tor, and Sch9. PLoS Genet. 2008, 4, e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.; Kim, M.S.; Paik, S.M.; Choi, S.H.; Cho, B.R.; Hahn, J.S. Rim15-dependent activation of Hsf1 and Msn2/4 transcription factors by direct phosphorylation in Saccharomyces cerevisiae. FEBS Lett. 2013, 587, 3648–3655. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Z.; Pinglay, S.; Ji, H.; Boeke, J.D. Msn2/4 regulate expression of glycolytic enzymes and control transition from quiescence to growth. eLife 2017, 6, e29938. [Google Scholar] [CrossRef]
- Lee, P.; Cho, B.R.; Joo, H.S.; Hahn, J.S. Yeast Yak1 kinase, a bridge between PKA and stress-responsive transcription factors, Hsf1 and Msn2/Msn4. Mol. Microbiol. 2008, 70, 882–895. [Google Scholar] [CrossRef]
- Liang, S.H.; Wu, H.; Wang, R.R.; Wang, Q.; Shu, T.; Gao, X.D. The TORC1-Sch9-Rim15 signaling pathway represses yeast-to-hypha transition in response to glycerol availability in the oleaginous yeast Yarrowia lipolytica. Mol. Microbiol. 2017, 104, 553–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Chow, E.W.L.; Wang, H.; Xu, X.; Cai, C.; Song, Y.; Wang, J.; Wang, Y. LncRNA DINOR is a virulence factor and global regulator of stress responses in Candida auris. Nat. Microbiol. 2021, 6, 842–851. [Google Scholar] [CrossRef]
- Yan, G.; Shen, X.; Jiang, Y. Rapamycin activates Tap42-associated phosphatases by abrogating their association with Tor complex 1. EMBO J. 2006, 25, 3546–3555. [Google Scholar] [CrossRef]
- Qian, B.; Liu, X.; Ye, Z.; Zhou, Q.; Liu, P.; Yin, Z.; Wang, W.; Zheng, X.; Zhang, H.; Zhang, Z. Phosphatase-associated protein MoTip41 interacts with the phosphatase MoPpe1 to mediate crosstalk between TOR and cell wall integrity signalling during infection by the rice blast fungus Magnaporthe oryzae. Environ. Microbiol. 2021, 23, 791–809. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Chen, C.; Yang, J.; Feng, W.; Liu, X.; Zuo, R.; Wang, J.; Yang, L.; Zhong, K.; Gao, C.; et al. Histone acetyltransferase MoHat1 acetylates autophagy-related proteins MoAtg3 and MoAtg9 to orchestrate functional appressorium formation and pathogenicity in Magnaporthe oryzae. Autophagy 2019, 15, 1234–1257. [Google Scholar] [CrossRef]
- Marroquin-Guzman, M.; Wilson, R.A. GATA-Dependent Glutaminolysis Drives Appressorium Formation in Magnaporthe oryzae by Suppressing TOR Inhibition of cAMP/PKA Signaling. PLoS Pathog. 2015, 11, e1004851. [Google Scholar] [CrossRef]
- Qiu, M.; Deng, Y.; Deng, Q.; Sun, L.; Fang, Z.; Wang, Y.; Huang, X.; Zhao, J. Cysteine Inhibits the Growth of Fusarium oxysporum and Promotes T-2 Toxin Synthesis through the Gtr/Tap42 Pathway. Microbiol. Spectr. 2022, 10, e0368222. [Google Scholar] [CrossRef] [PubMed]
- López-Berges, M.S.; Rispail, N.; Prados-Rosales, R.C.; Di Pietro, A. A nitrogen response pathway regulates virulence functions in Fusarium oxysporum via the protein kinase TOR and the bZIP protein MeaB. Plant Cell 2010, 22, 2459–2475. [Google Scholar] [CrossRef] [Green Version]
- Harata, K.; Nishiuchi, T.; Kubo, Y. Colletotrichum orbiculare WHI2, a Yeast Stress-Response Regulator Homolog, Controls the Biotrophic Stage of Hemibiotrophic Infection Through TOR Signaling. Mol. Plant. Microbe Interact. 2016, 29, 468–483. [Google Scholar] [CrossRef]
- Hu, G.; McQuiston, T.; Bernard, A.; Park, Y.D.; Qiu, J.; Vural, A.; Zhang, N.; Waterman, S.R.; Blewett, N.H.; Myers, T.G.; et al. A conserved mechanism of TOR-dependent RCK-mediated mRNA degradation regulates autophagy. Nat. Cell Biol. 2015, 17, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.D.; Jarvis, J.N.; Hu, G.; Davis, S.E.; Qiu, J.; Zhang, N.; Hollingsworth, C.; Loyse, A.; Gardina, P.J.; Valyi-Nagy, T.; et al. Transcriptional Profiling of Patient Isolates Identifies a Novel TOR/Starvation Regulatory Pathway in Cryptococcal Virulence. mBio 2018, 9, e02353-02318. [Google Scholar] [CrossRef] [Green Version]
- Alspaugh, J.A.; Pukkila-Worley, R.; Harashima, T.; Cavallo, L.M.; Funnell, D.; Cox, G.M.; Perfect, J.R.; Kronstad, J.W.; Heitman, J. Adenylyl cyclase functions downstream of the Galpha protein Gpa1 and controls mating and pathogenicity of Cryptococcus neoformans. Eukaryot. Cell. 2002, 1, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Stadtländer, K. The aspergilli: Genomics, medical aspects, biotechnology, and research methods. Aspergilli Genom. Med. Asp. Biotechnol. Res. Methods 2007, 4, 201. [Google Scholar] [CrossRef] [Green Version]
- Baldin, C.; Valiante, V.; Krüger, T.; Schafferer, L.; Haas, H.; Kniemeyer, O.; Brakhage, A.A. Comparative proteomics of a tor inducible Aspergillus fumigatus mutant reveals involvement of the Tor kinase in iron regulation. Proteomics 2015, 15, 2230–2243. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B.; Gangwar, G.; Nain, V.; Gupta, I.; Thakur, A.; Puria, R. Rapamycin and Torin2 inhibit Candida auris TOR: Insights through growth profiling, docking, and MD simulations. J. Biomol. Struct. Dyn. 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Romero-Aguilar, L.; Guerra-Sánchez, G.; Tenorio, E.P.; Tapia-Rodriguez, M.; Matus-Ortega, G.; Flores-Herrera, O.; González, J.; Pardo, J.P. Rapamycin induces morphological and physiological changes without increase in lipid content in Ustilago maydis. Arch. Microbiol. 2020, 202, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Freitag, J.; Lanver, D.; Böhmer, C.; Schink, K.O.; Bölker, M.; Sandrock, B. Septation of infectious hyphae is critical for appressoria formation and virulence in the smut fungus Ustilago maydis. PLoS Pathog. 2011, 7, e1002044. [Google Scholar] [CrossRef] [Green Version]
- Ghaddar, K.; Merhi, A.; Saliba, E.; Krammer, E.M.; Prévost, M.; André, B. Substrate-induced ubiquitylation and endocytosis of yeast amino acid permeases. Mol. Cell. Biol. 2014, 34, 4447–4463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merhi, A.; André, B. Internal amino acids promote Gap1 permease ubiquitylation via TORC1/Npr1/14-3-3-dependent control of the Bul arrestin-like adaptors. Mol. Cell. Biol. 2012, 32, 4510–4522. [Google Scholar] [CrossRef]
- Otsubo, Y.; Yamashita, A.; Ohno, H.; Yamamoto, M.S. pombe TORC1 activates the ubiquitin-proteasomal degradation of the meiotic regulator Mei2 in cooperation with Pat1 kinase. J. Cell Sci. 2014, 127, 2639–2646. [Google Scholar] [PubMed] [Green Version]
- Otsubo, Y.; Nakashima, A.; Yamamoto, M.; Yamashita, A. TORC1-Dependent Phosphorylation Targets in Fission Yeast. Biomolecules 2017, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Lee, N.N.; Pan, L.; Dhakshnamoorthy, J.; Sun, L.L.; Zofall, M.; Wheeler, D.; Grewal, S.I.S. TOR targets an RNA processing network to regulate facultative heterochromatin, developmental gene expression and cell proliferation. Nat. Cell Biol. 2021, 23, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Urano, J.; Tabancay, A.P.; Yang, W.; Tamanoi, F. The Saccharomyces cerevisiae Rheb G-protein is involved in regulating canavanine resistance and arginine uptake. J. Biol. Chem. 2000, 275, 11198–11206. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Regulatory Factors | Fungal Species | Human Homolog | Function or Functional Mechanisms |
|---|---|---|---|
| Rag-GTPase (Gtr1/Gtr2) | S. cerevisiae, S. pombe, F. oxysporum | Rag A/B, Rag C/D | recruits TORC1 to vacuolar membrane |
| EGO ternary complex | S. cerevisiae, S. pombe | Ragulator | as a scaffold tethering RAG heterodimer on vacuolar membrane |
| Vam6 | S. cerevisiae, S. pombe | Ragulator | has GEF activity that acts on Gtr1 GTP to positively regulate TORC1 |
| Seh1-associated complex (SEAC) (SEAIT and SEACT) | S. cerevisiae S. pombe (GATOR complex) | GATOR complex (GATOR1 and GATOR2) | SEAIT/GATOR1: has GAP activity that acts on Gtr1 GTP to negatively regulate TORC1 SEACT/GATOR2: antagonizes the GAP activity of SEAIT to participate in the activation of TORC1 |
| Lst4–Lst7 | S. cerevisiae S. pombe (Lst4– Bhd1) | FLCN–FNIP | acts as GAP of Gtr2 to stimulate the activity of TORC1 |
| Vps34–Vps15 | S. cerevisiae | Vps34–Vps15 | recruits Pib2 to the vacuolar membrane in TORC1 glutamine reaction |
| Pib2 | S. cerevisiae S. pombe (SPBC9B6.03 protein) | - | bound to the vacuolar membrane through its FYVE domain is targeted to TORC1 |
| Ait1 | S. cerevisiae | - | binds to the vacuolar membrane and interacts with EGO complex to inhibit Gtr-induced TORC1 activation |
| Ivy1 | S. cerevisiae | missing-in-metastasis (MIM) | a negative regulator of Gtr-dependent-TORC1 activation |
| Whi2 | S. cerevisiae | potassium channel tetramerization domain containing 11 (KCTD11) protein | a conserved negative regulator of TORC1 in response to low amino acids |
| Free uncharged tRNA | S. cerevisiae S. pombe | tRNA | deactivates TORC1 |
| Ubiquitin | S. cerevisiae | Ubiquitin | regulates the degradation of Kog1 and the stability of TORC1 through non-covalent binding to Kog1 |
| Fungi Species | Regulatory Factors | Regulation of Physiological Processes |
|---|---|---|
| S. cerevisiae | Tap42-PPase, Gcn2, and eIF2α | protein translation |
| Sch9, ypk3, MAF1, Ifh1, Stb3, Dot6, and Tod6 | ribosome biosynthesis | |
| PP2A or Sit4, Stp1, Gln3, and Gat1 | sensing, transport, and catabolic regulation of nitrogen sources | |
| Sch9, Ydc1, and Ypc1 ceramidase | sphingolipid metabolism | |
| Atg13, Rim15, and Msn2/4, Tap42-PP2A, and Ecm33 | macroautophagy | |
| Npr1 | ubiquitination of Gap1 | |
| Yak1 and Rim15, Msn2/4, and Gis1 | stress response | |
| S. pombe | Gcn2 and eIF2α | protein translation |
| AGC kinases and Psk1 | phosphorylation of Rps6 | |
| PP2A-like phosphatase Ppe1 and Gaf1 | nitrogen metabolism | |
| Atg13 | macroautophagy | |
| Mie2, Ste11 | sexual differentiation | |
| - | cell proliferation and gametogenesis | |
| Y. lipolytic | Sch9–Rim15 | the yeast-to-hypha dimorphic transition |
| Dictyostelium discoideum | 4EBP1 and eIF4E | protein translation |
| Candida auris | DINOR | stress response |
| - | G1 cell cycle arrest and cytokinesis defect | |
| Magnaporthe oryzae | PP6 catalytic subunit MoPpe1 and MoTap42 | toxicity and activation of the cell-wall integrity (CWI) pathway |
| MoATG3, MoAtg9, and Atg13 | autophagy, control appressoria function and infection growth | |
| F. oxysporum | Gtr/Tap42 pathway | the growth of hyphae and the synthesis of T-2 toxin |
| Coletothrichum orbiculare | CoWhi2 and CoMsn2 | pathogenicity and virulence |
| Cryptococcus neoformans | STL1 and VAD1–EM15–CAC2 | virulence |
| Aspergillus fumigatus | SchA | the regulation of high osmolarity, glycerol (HOG), and calcineurin/CrzA pathway calcineurin, as well as stress response and toxicity of mammalian host |
| - | production of iron carrier | |
| Ustilago maydis | Ras-like GTPases (Don1, Don3, and Cdc42) | diaphragm formation |
| Sch9, PP2A-B55, and PP2A-B55 | inhibit Cdk1; cytokinesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zheng, X.; Li, G.; Wang, X. TORC1 Signaling in Fungi: From Yeasts to Filamentous Fungi. Microorganisms 2023, 11, 218. https://doi.org/10.3390/microorganisms11010218
Wang Y, Zheng X, Li G, Wang X. TORC1 Signaling in Fungi: From Yeasts to Filamentous Fungi. Microorganisms. 2023; 11(1):218. https://doi.org/10.3390/microorganisms11010218
Chicago/Turabian StyleWang, Yuhua, Xi Zheng, Guohong Li, and Xin Wang. 2023. "TORC1 Signaling in Fungi: From Yeasts to Filamentous Fungi" Microorganisms 11, no. 1: 218. https://doi.org/10.3390/microorganisms11010218