

Klebsiella pneumoniae Volatile Organic Compounds (VOCs) Protect Artemia salina from Fish Pathogen Aeromonas sp.: A Combined In Vitro, In Vivo, and In Silico Approach

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Selection
2.2. In Vitro Antagonism Test with the Selected Strains
2.3. Biofilm Formation Assay of the Selected Strains
2.4. Inhibition of Biofilm Formation by CFSs of the Selected Strains
2.5. Scanning Electronic Microscope (SEM)
2.6. In Vivo Effect of Bacteria on Brine Shrimp
2.7. In Vivo Antagonism Test against Aeromonas sp.
2.8. Volatile Compounds Analysis through GC-MS
2.9. Ligand and Protein Preparation and Optimization
2.10. Molecular Docking Analysis
2.11. Pharmacokinetics Properties Analysis
2.12. Molecular Dynamics Simulation
2.13. Statistical Analysis
3. Results
3.1. In Vitro Antagonism Test of the Selected Bacterial Strains
3.2. Capability of Biofilm Formation by the Selected Bacterial Strain
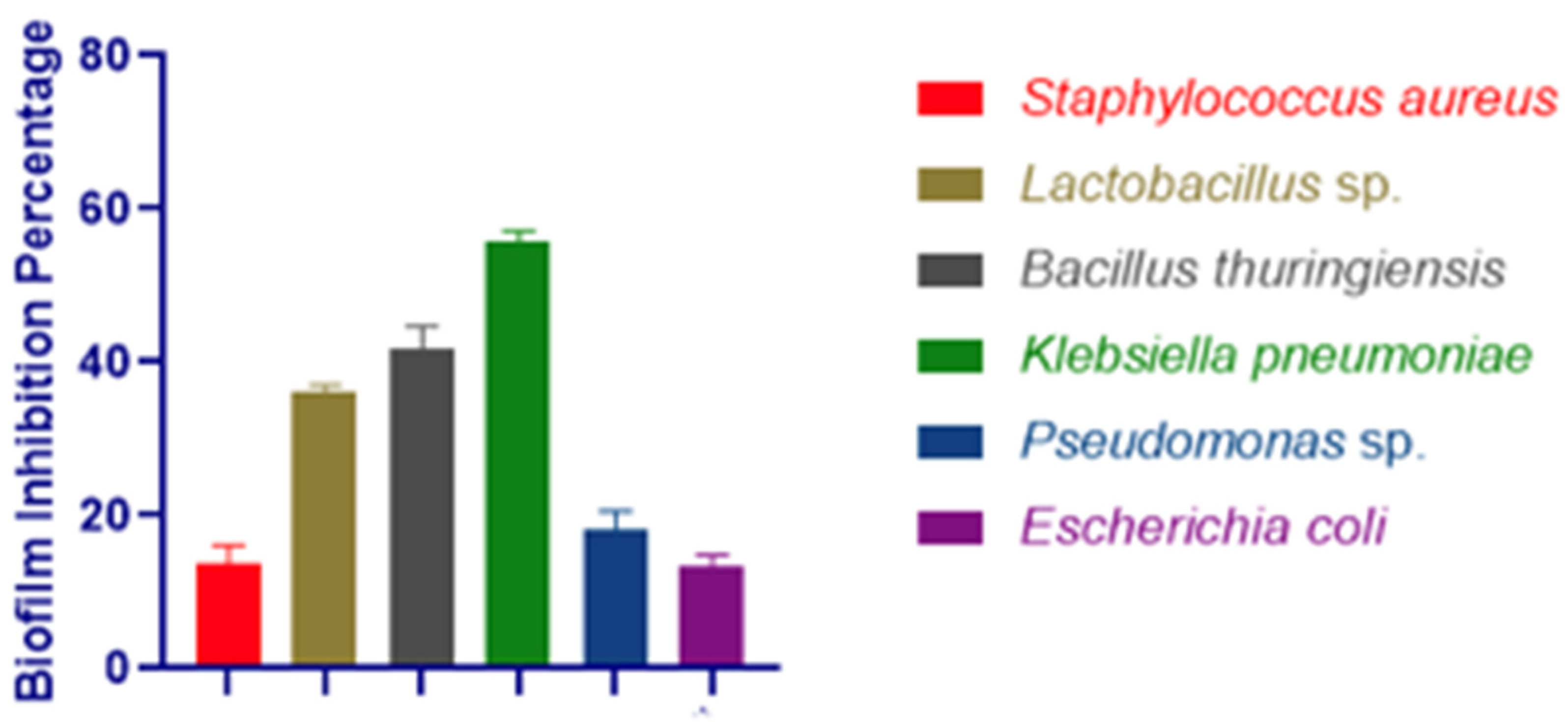
3.3. Biofilm Inhibition Assay
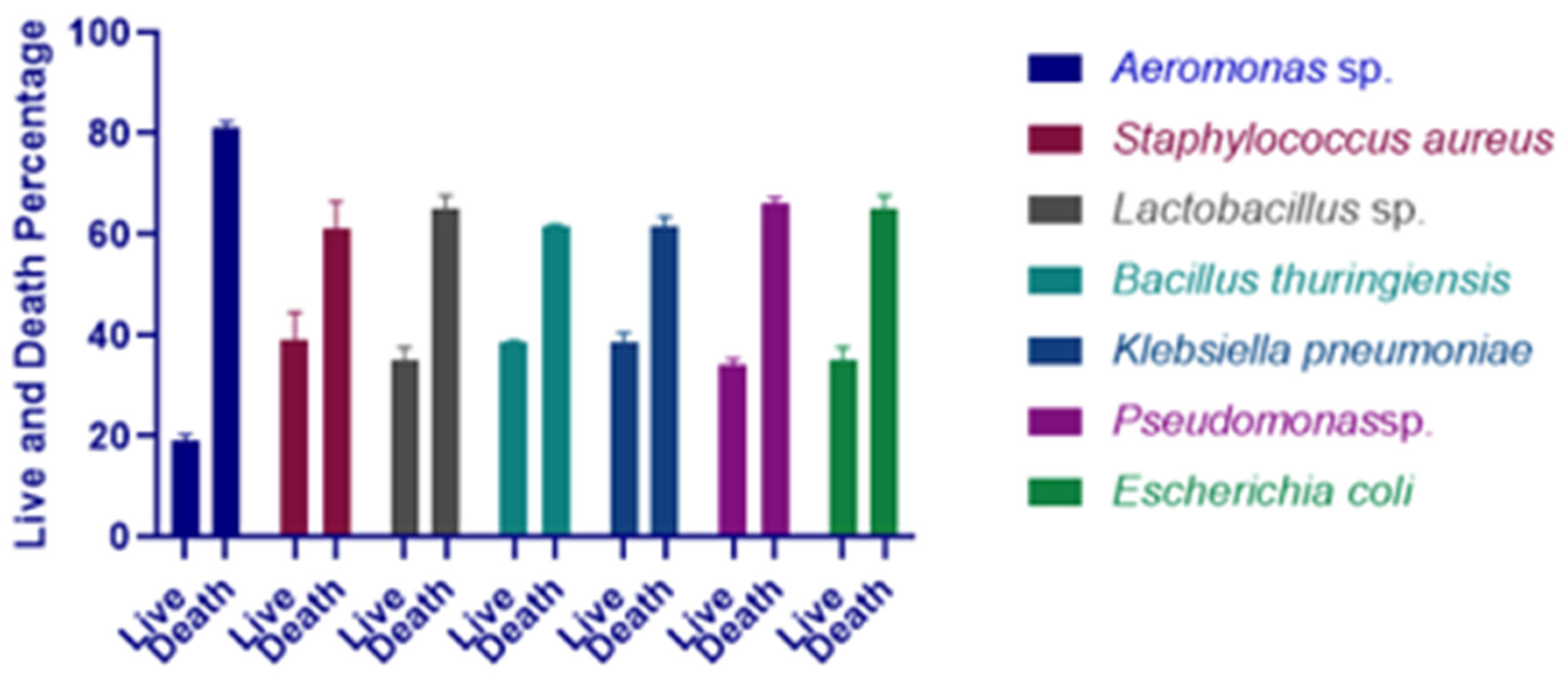
3.4. Effect of Selected Bacterial Strains on Artemia salina
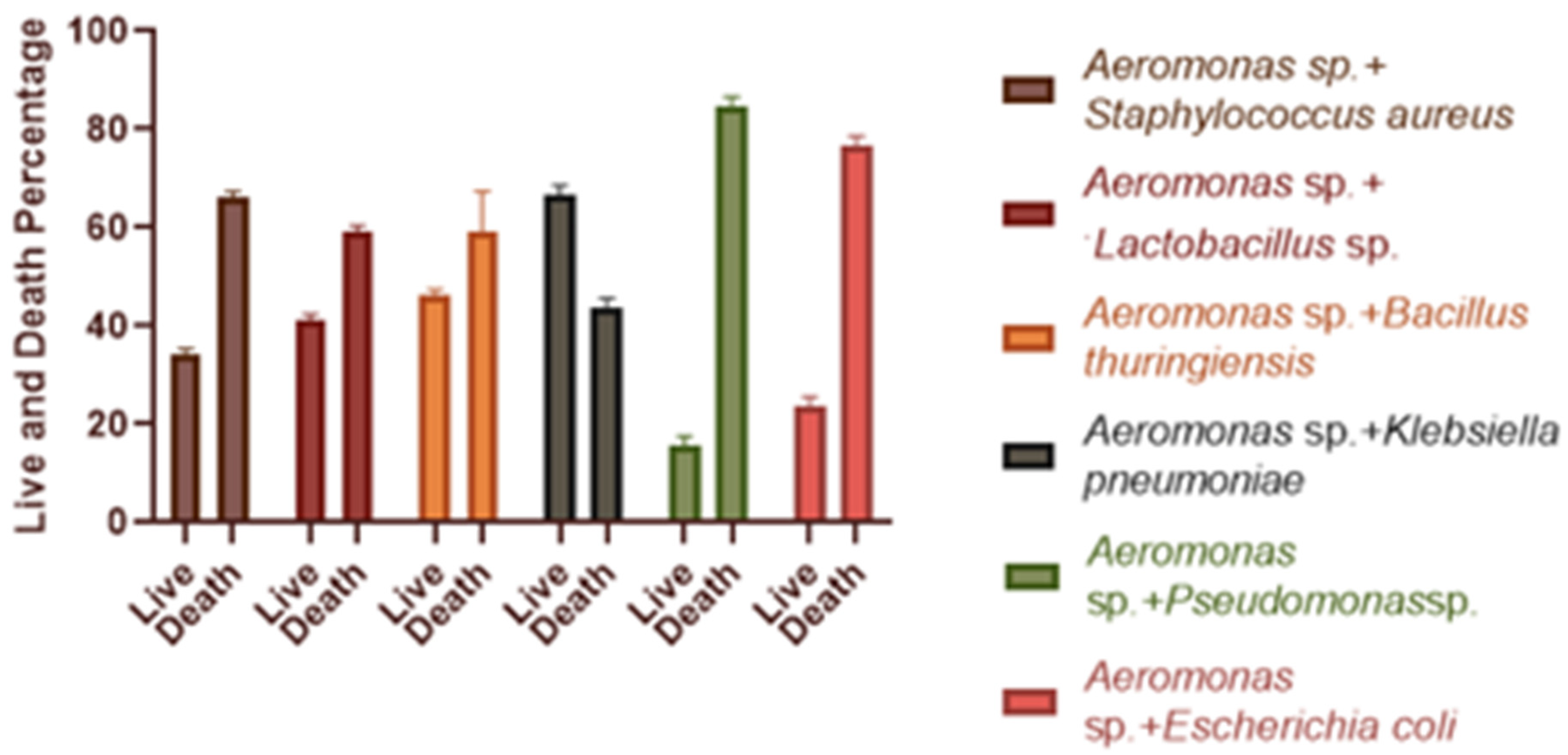
3.5. Effect of Single and Dual Bacteria on Artemia salina
3.6. Determination of Binding Energy
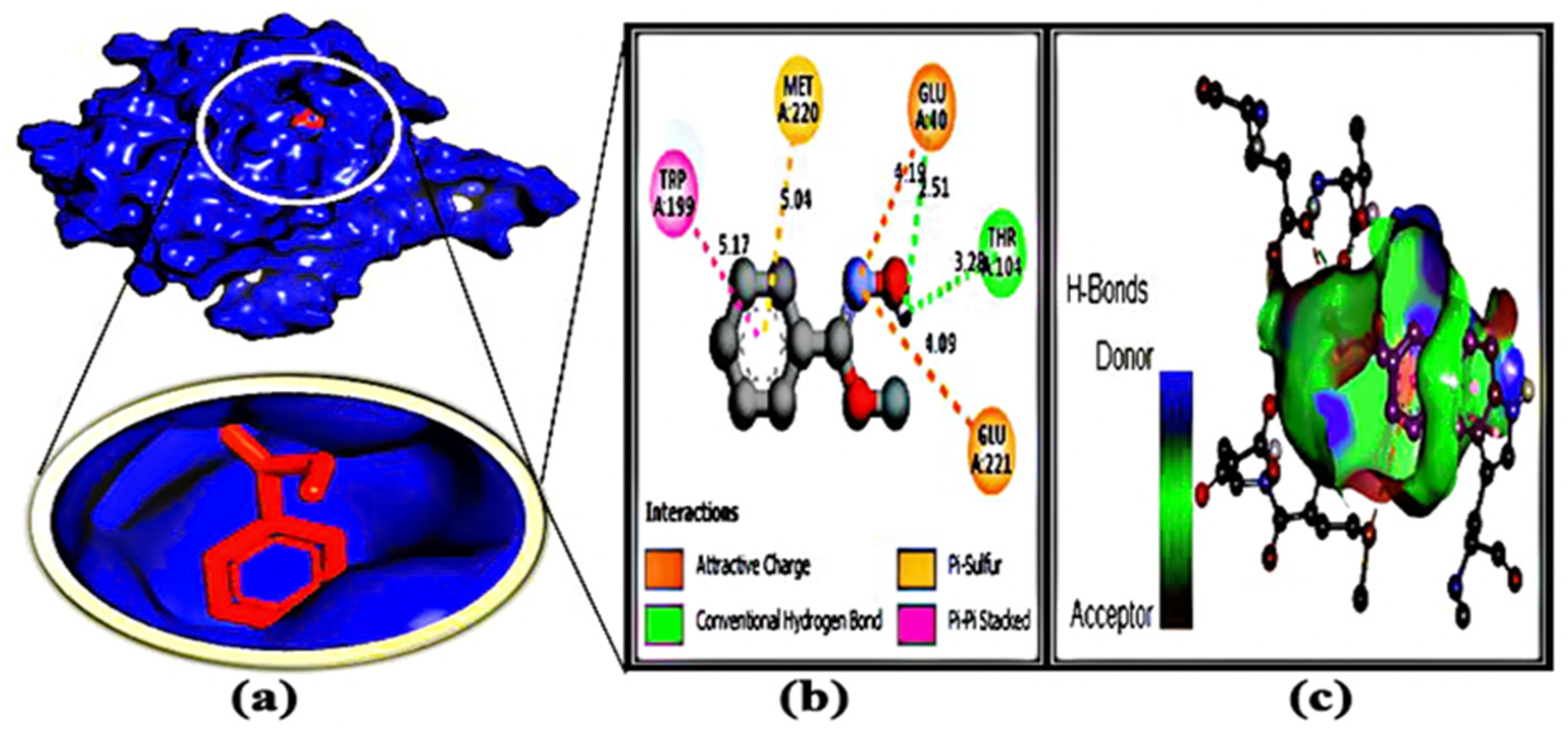
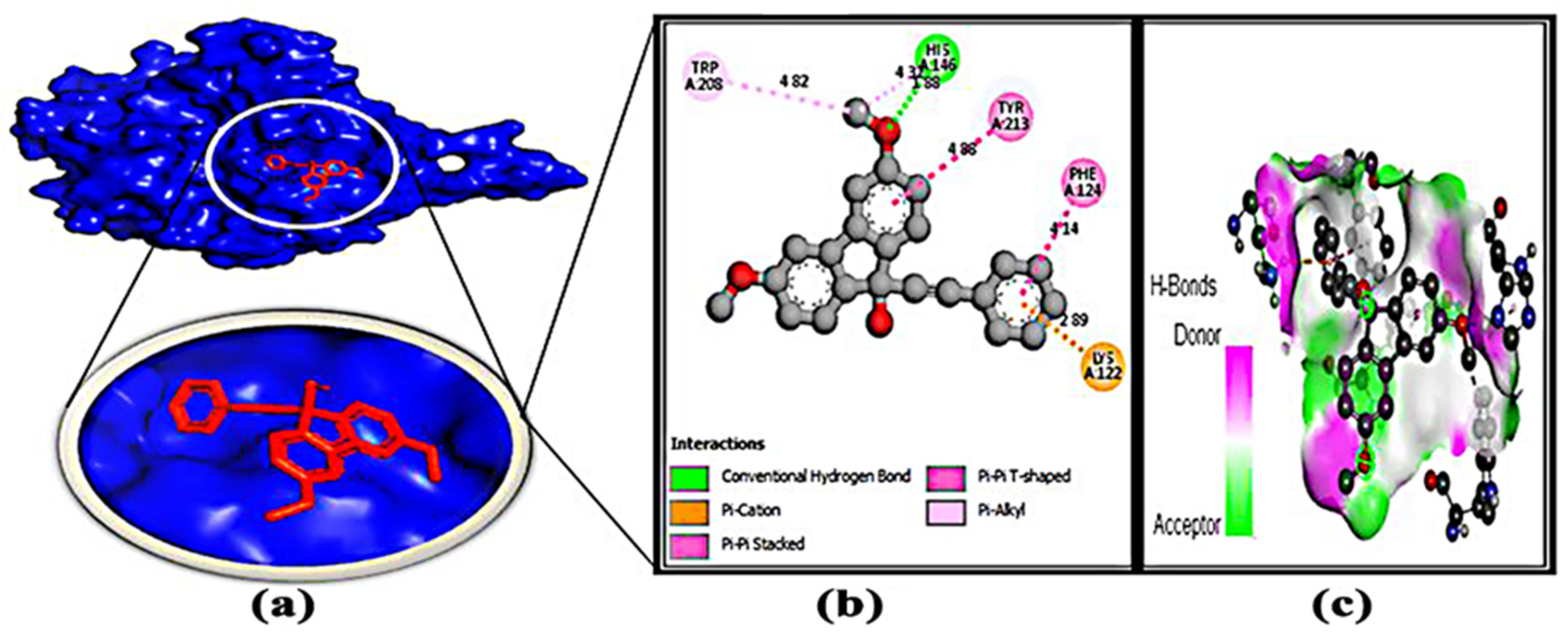
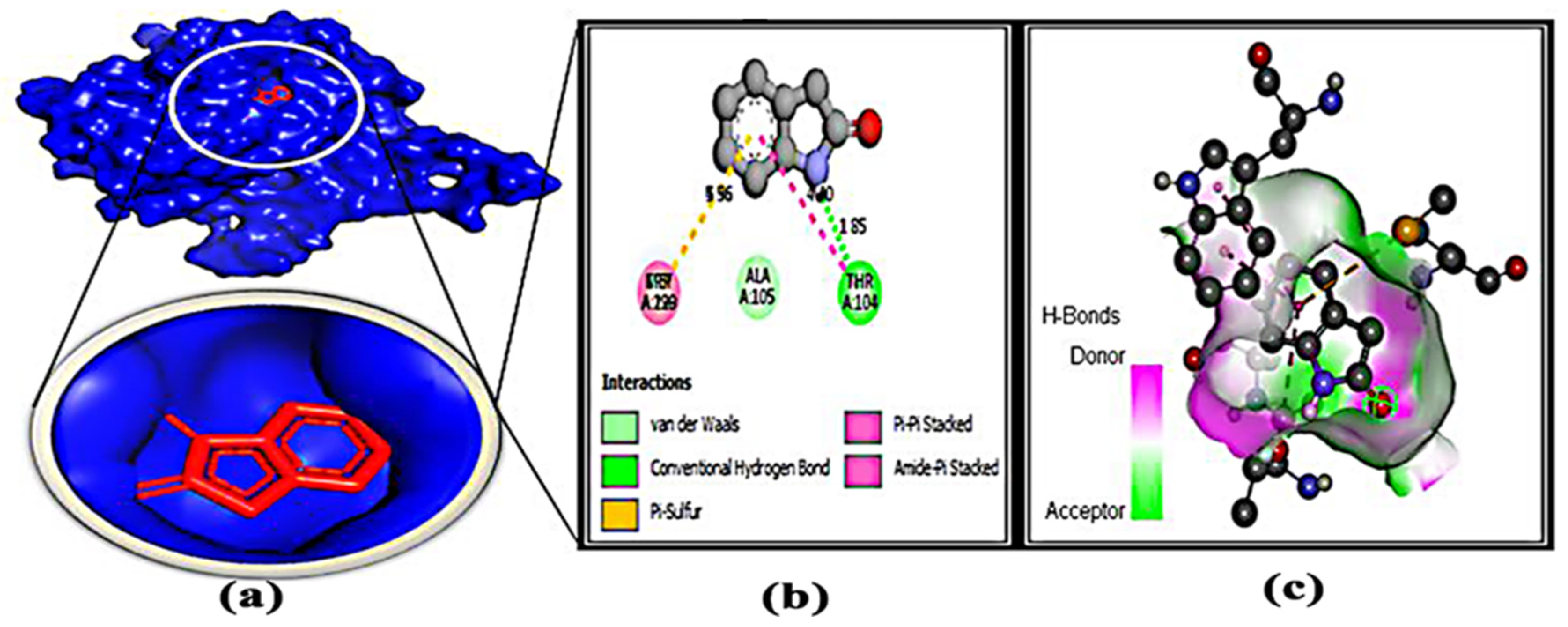
3.7. Protein–Ligand Interaction
3.8. Pharmacological Properties Assessment
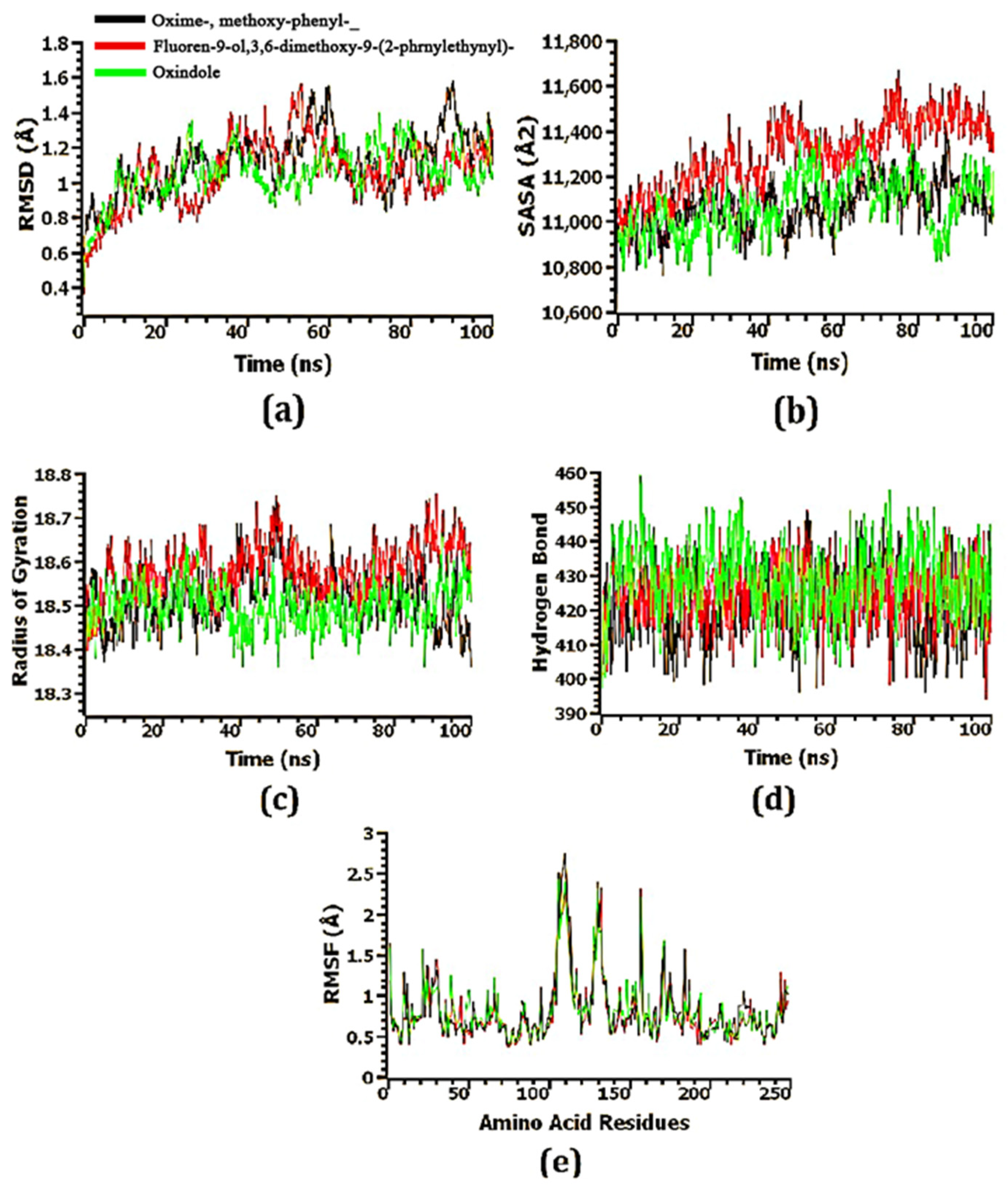
3.9. Confirmation of the Stability of Ligand–Protein Complexes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toja, Y.T.; Suprayitno, E.; Yanuhar, U. In Silico Potential Black Fruit Seeds (Haplolobus Monticola) Wondama Local Plant West Papua Barat as Antibacterial Aeromonas Hydrophila. Eurasian J. Biosci. 2020, 7131, 7125–7131. [Google Scholar]
- Moriel, B.; de Campos Prediger, K.; de Souza, E.M.; Pedrosa, F.O.; Fadel-Picheth, C.M.T.; Cruz, L.M. In Silico Comparative Analysis of Aeromonas Type VI Secretion System. Braz. J. Microbiol. 2021, 52, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bravo, A.; Figueras, M.J. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, J.M.; Abbott, S.L. The Genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teunis, P.; Figueras, M.J. Reassessment of the Enteropathogenicity of Mesophilic Aeromonas Species. Front. Microbiol. 2016, 7, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatasamy, V.; Durairaj, R.; Karuppaiah, P.; Sridhar, A.; Kamaraj, S.K.; Ramasamy, T. An In Silico Evaluation of Molecular Interaction Between Antimicrobial Peptide Subtilosin A of Bacillus Subtilis with Virulent Proteins of Aeromonas Hydrophila. Int. J. Pept. Res. Ther. 2021, 27, 1709–1718. [Google Scholar] [CrossRef]
- Yadav, S.K.; Panwar, D.; Singh, A.; Tellis, M.B.; Joshi, R.S.; Dixit, A. Molecular Phylogeny, Structure Modeling and in Silico Screening of Putative Inhibitors of Aerolysin of Aeromonas Hydrophila EUS112. J. Biomol. Struct. Dyn. 2021. [Google Scholar] [CrossRef]
- Vivekanandhan, G.; Savithamani, K.; Hatha, A.A.M.; Lakshmanaperumalsamy, P. Antibiotic Resistance of Aeromonas Hydrophila Isolated from Marketed Fish and Prawn of South India. Int. J. Food Microbiol. 2002, 76, 165–168. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Asmat, A.; Usup, G. Antibiotics Resistance and Putative Virulence Factors of Aeromonas Hydrophila Isolated from Estuary. J. Microbiol. 2012, 9, 1339–1357. [Google Scholar]
- Xu, Y.; Wang, L.; Chen, J.; Zhao, J.; Fan, S.; Dong, Y.; Ha, N.C.; Quan, C. Structural and Functional Analyses of Periplasmic 5′-Methylthioadenosine/S-Adenosylhomocysteine Nucleosidase from Aeromonas Hydrophila. Biochemistry 2017, 56, 5347–5355. [Google Scholar] [CrossRef]
- Hassan, M.; Kjos, M.; Nes, I.F.; Diep, D.B.; Lotfipour, F. Natural Antimicrobial Peptides from Bacteria: Characteristics and Potential Applications to Fight against Antibiotic Resistance. J. Appl. Microbiol. 2012, 113, 723–736. [Google Scholar] [CrossRef]
- Nascimento, G.G.F.; Locatelli, J.; Freitas, P.C.; Silva, G.L. Antibacterial Activity of Plant Extracts and Phytochemicals on Antibiotic-Resistant Bacteria. Braz. J. Microbiol. 2000, 31, 247–256. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The Antimicrobial Volatile Power of the Rhizospheric Isolate Pseudomonas Donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [Green Version]
- Islam1, S.; Mahmud1, M.L.; Almalki, W.H.; Biswas, S.; Islam, M.A.; Mortuza, M.G.; Hossain, M.A.; Ekram, M.A.-E.; Uddin, M.S.; Zaman, S.; et al. Cell-Free Supernatants (CFSs) from the Culture of Bacillus Subtilis Inhibit Pseudomonas Sp. Biofilm Formation. Microorganisms 2022, 10, 2105. [Google Scholar]
- Valença, C.A.S.; Barbosa, A.A.T.; Souto, E.B.; Caramão, E.B.; Jain, S. Volatile Nitrogenous Compounds from Bacteria: Source of Novel Bioactive Compounds. Chem. Biodivers. 2021, 18, e2100549. [Google Scholar] [CrossRef]
- Lammers, A.; Lalk, M.; Garbeva, P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics 2022, 11, 109. [Google Scholar] [CrossRef]
- Avalos, M.; van Wezel, G.P.; Raaijmakers, J.M.; Garbeva, P. Healthy Scents: Microbial Volatiles as New Frontier in Antibiotic Research? Curr. Opin. Microbiol. 2018, 45, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, G.; Motta, C.M.; Agnisola, C.; Venditti, P.; Fasciolo, G.; Ferrandino, I.; Capriello, T.; Vitale, E.; Costanzo, G.; Avallone, B.; et al. Commercial Red Food Dyes Preparations Modulate the Oxidative State in Three Model Organisms (Cucumis Sativus, Artemia Salina, and Danio Rerio). Environments 2022, 9, 63. [Google Scholar] [CrossRef]
- Mohd-Aris, A.; Muhamad-Sofie, M.H.N.; Zamri-Saad, M.; Daud, H.M.; Yasin Ina-Salwany, M. Live Vaccines against Bacterial Fish Diseases: A Review. Vet. World 2019, 12, 1806–1815. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Bio, A.M.; Amat, F.D.; Monteiro, N.M.; Vieira, N.M. Surviving an Invasion: Characterization of One of the Last Refugia for Artemia Diploid Parthenogenetic Strains. Wetlands 2012, 32, 1079–1090. [Google Scholar] [CrossRef] [Green Version]
- Overton, S.V.; Bland, C.E. Infection of Artemia Salina by Haliphthoros Milfordensis: A Scanning and Transmission Electron Microscope Study. J. Invertebr. Pathol. 1981, 37, 249–257. [Google Scholar] [CrossRef]
- Morya, V.K.; Choi, W.; Kim, E.K. Isolation and Characterization of Pseudoalteromonas Sp from Fermented Korean Food, as an Antagonist to Vibrio Harveyi. Appl. Microbiol. Biotechnol. 2014, 98, 1389–1395. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef]
- Guo, R.; Zhao, Y.; Zou, Q.; Fang, X.; Peng, S. Bioinformatics Applications on Apache Spark. Gigascience 2018, 7, giy098. [Google Scholar] [CrossRef]
- Dao, F.Y.; Lv, H.; Wang, F.; Feng, C.Q.; Ding, H.; Chen, W.; Lin, H. Identify Origin of Replication in Saccharomyces Cerevisiae Using Two-Step Feature Selection Technique. Bioinformatics 2019, 35, 2075–2083. [Google Scholar] [CrossRef]
- Ivanov, J.; Polshakov, D.; Kato-Weinstein, J.; Zhou, Q.; Li, Y.; Granet, R.; Garner, L.; Deng, Y.; Liu, C.; Albaiu, D.; et al. Quantitative Structure−activity Relationship Machine Learning Models and Their Applications for Identifying Viral 3Clpro- And RDRP-Targeting Compounds as Potential Therapeutics for Covid-19 and Related Viral Infections. ACS Omega 2020, 5, 27344–27358. [Google Scholar] [CrossRef]
- Mahmud, S.; Paul, G.K.; Afroze, M.; Islam, S.; Gupt, S.B.R.; Razu, M.H.; Biswas, S.; Zaman, S.; Uddin, M.S.; Khan, M.; et al. Efficacy of Phytochemicals Derived from Avicennia Officinalis for the Management of Covid-19: A Combined in Silico and Biochemical Study. Molecules 2021, 26, 2210. [Google Scholar] [CrossRef]
- Scieuzo, C.; Nardiello, M.; Farina, D.; Scala, A.; Cammack, J.A.; Tomberlin, J.K.; Vogel, H.; Salvia, R.; Persaud, K.; Falabella, P. Hermetia Illucens (L.) (Diptera: Stratiomyidae) Odorant Binding Proteins and Their Interactions with Selected Volatile Organic Compounds: An in Silico Approach. Insects 2021, 12, 814. [Google Scholar] [CrossRef]
- Karimi, S.; Rashidian, E.; Birjandi, M.; Mahmoodnia, L. Antagonistic Effect of Isolated Probiotic Bacteria from Natural Sources against Intestinal Escherichia Coli Pathotypes. Electron. Physician 2018, 10, 6534–6539. [Google Scholar] [CrossRef] [Green Version]
- Di Marco, N.I.; Pungitore, C.R.; Lucero-Estrada, C.S.M. Aporphinoid Alkaloids Inhibit Biofilm Formation of Yersinia Enterocolitica Isolated from Sausages. J. Appl. Microbiol. 2020, 129, 1029–1042. [Google Scholar] [CrossRef]
- Saito, Y.; Fujii, R.; Nakagawa, K.I.; Kuramitsu, H.K.; Okuda, K.; Ishihara, K. Stimulation of Fusobacterium Nucleatum Biofilm Formation by Porphyromonas Gingivalis. Oral Microbiol. Immunol. 2008, 23, 1–6. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2010, 47, e2437. [Google Scholar] [CrossRef]
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm Formation and Antibiotic Resistance of Klebsiella Pneumoniae Isolated from Clinical Samples in a Tertiary Care Hospital, Klaten, Indonesia. BMC Proc. 2019, 13. [Google Scholar] [CrossRef]
- Junfen, L.I.N.; Mengna, W.U.; Haocheng, W.U.; ZHANG, T.; Chen, W.U.; Fudong, L.I. Epidemiological Characteristics of Coronavirus Disease 2019 in Zhejiang Province. J. Prev. Med. 2020, 12, 217–223. [Google Scholar]
- Rakib, A.; Nain, Z.; Sami, S.A.; Mahmud, S.; Islam, A.; Ahmed, S.; Siddiqui, A.B.F.; Babu, S.M.O.F.; Hossain, P.; Shahriar, A.; et al. A Molecular Modelling Approach for Identifying Antiviral Selenium-Containing Heterocyclic Compounds That Inhibit the Main Protease of SARS-CoV-2: An in Silico Investigation. Brief. Bioinform. 2021, 22, 1476–1498. [Google Scholar] [CrossRef]
- Bueno, M.D.L.G.B. Molecular Docking, Pharmacokinetic, and DFT Calculation of Naproxen and Its Degradants. Biomed. J. Sci. Tech. Res. 2018, 9(5), 7360–7365. [Google Scholar] [CrossRef] [Green Version]
- Hashem, H.E.; Nath, A.; Kumer, A. Synthesis, Molecular Docking, Molecular Dynamic, Quantum Calculation, and Antibacterial Activity of New Schiff Base-Metal Complexes. J. Mol. Struct. 2021, 1250, 131915. [Google Scholar] [CrossRef]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Depicting the Inhibitory Potential of Polyphenols from Isatis Indigotica Root against the Main Protease of SARS CoV-2 Using Computational Approaches. J. Biomol. Struct. Dyn. 2020, 40, 4110–4121. [Google Scholar] [CrossRef]
- Nath, A.; Kumer, A.; Zaben, F.; Khan, M.W. Investigating the Binding Affinity, Molecular Dynamics, and ADMET Properties of 2,3-Dihydrobenzofuran Derivatives as an Inhibitor of Fungi, Bacteria, and Virus Protein. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 36. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.M.A.; Kumer, A. Computational Investigation of Methyl α-d-Glucopyranoside Derivatives as Inhibitor against Bacteria, Fungi and COVID-19 (Sars-2). J. Chil. Chem. Soc. 2021, 66, 5206–5214. [Google Scholar]
- Punjabi, M.; Bharadvaja, N.; Sachdev, A.; Krishnan, V. Molecular Characterization, Modeling, and Docking Analysis of Late Phytic Acid Biosynthesis Pathway Gene, Inositol Polyphosphate 6-/3-/5-Kinase, a Potential Candidate for Developing Low Phytate Crops. 3 Biotech 2018, 8, 344. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Krieger Elmar, G.V.; Spronk, C. YASARA–Yet Another Scientific Artificial Reality Application. YASARA Org 2013, 993, 51–78. [Google Scholar]
- Harrach, M.F.; Drossel, B. Structure and Dynamics of TIP3P, TIP4P, and TIP5P Water near Smooth and Atomistic Walls of Different Hydroaffinity. J. Chem. Phys. 2014, 140, 174501. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. New Ways to Boost Molecular Dynamics Simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Harvey, M.J.; De Fabritiis, G. An Implementation of the Smooth Particle Mesh Ewald Method on GPU Hardware. J. Chem. Theory Comput. 2009, 5, 2371–2377. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- Krieger, E.; Nielsen, J.E.; Spronk, C.A.E.M.; Vriend, G. Fast Empirical PKa Prediction by Ewald Summation. J. Mol. Graph. Model. 2006, 25, 481–486. [Google Scholar] [CrossRef]
- Mahmud, S.; Rafi, O.; Paul, G.K.; Promi, M.M.; Shimu, M.S.S.; Biswas, S.; Bin Emran, T.; Dhama, K.; Alyami, S.A.; Moni, M.A.; et al. Designing a Multi-Epitope Vaccine Candidate to Combat MERS-CoV by Employing an Immunoinformatics Approach. Sci. Rep. 2021, 11, 1–20. [Google Scholar] [CrossRef]
- Mahmud, S.; Hasan, M.R.; Biswas, S.; Paul, G.K.; Afrose, S.; Mita, M.A.; Sultana Shimu, M.S.; Promi, M.M.; Hani, U.; Rahamathulla, M.; et al. Screening of Potent Phytochemical Inhibitors Against SARS-CoV-2 Main Protease: An Integrative Computational Approach. Front. Bioinforma. 2021, 1, 717141. [Google Scholar] [CrossRef]
- Mahmud, S.; Paul, G.K.; Biswas, S.; Afrose, S.; Mita, M.A.; Hasan, M.R.; Shimu, M.S.S.; Hossain, A.; Promi, M.M.; Ema, F.K.; et al. Prospective Role of Peptide-Based Antiviral Therapy Against the Main Protease of SARS-CoV-2. Front. Mol. Biosci. 2021, 8, 628585. [Google Scholar] [CrossRef]
- Mahmud, S.; Biswas, S.; Paul, G.K.; Mita, M.A.; Promi, M.M.; Afrose, S.; Hasan, M.R.; Zaman, S.; Uddin, M.S.; Dhama, K.; et al. Plant-Based Phytochemical Screening by Targeting Main Protease of Sars-Cov-2 to Design Effective Potent Inhibitors. Biology 2021, 10, 589. [Google Scholar] [CrossRef]
- Kumar Paul, G.; Mahmud, S.; Aldahish, A.A.; Afroze, M.; Biswas, S.; Briti Ray Gupta, S.; Hasan Razu, M.; Zaman, S.; Salah Uddin, M.; Nahari, M.H.; et al. Computational Screening and Biochemical Analysis of Pistacia Integerrima and Pandanus Odorifer Plants to Find Effective Inhibitors against Receptor-Binding Domain (RBD) of the Spike Protein of SARS-Cov-2. Arab. J. Chem. 2022, 15, 103600. [Google Scholar] [CrossRef]
- Mahmud, S.; Mita, M.A.; Biswas, S.; Paul, G.K.; Promi, M.M.; Afrose, S.; Hasan, R.; Shimu, S.S.; Zaman, S.; Uddin, S.; et al. Molecular Docking and Dynamics Study to Explore Phytochemical Ligand Molecules against the Main Protease of SARS-CoV-2 from Extensive Phytochemical Datasets. Expert Rev. Clin. Pharmacol. 2021, 14, 1305–1315. [Google Scholar] [CrossRef]
- Mahmud, S.; Biswas, S.; Kumar Paul, G.; Mita, M.A.; Afrose, S.; Robiul Hasan, M.; Sharmin Sultana Shimu, M.; Uddin, M.A.R.; Salah Uddin, M.; Zaman, S.; et al. Antiviral Peptides against the Main Protease of SARS-CoV-2: A Molecular Docking and Dynamics Study. Arab. J. Chem. 2021, 14, 103315. [Google Scholar] [CrossRef]
- Jagannathan, R. Characterization of Drug-like Chemical Space for Cytotoxic Marine Metabolites Using Multivariate Methods. ACS Omega 2019, 4, 5402–5411. [Google Scholar] [CrossRef] [Green Version]
- Defoirdt, T.; Boon, N.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Alternatives to Antibiotics to Control Bacterial Infections: Luminescent Vibriosis in Aquaculture as an Example. Trends Biotechnol. 2007, 25, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Ali, F.; Yao, Z.; Li, W.; Sun, L.; Lin, W.; Lin, X. In-Silico Prediction and Modeling of the Quorum Sensing Luxs Protein and Inhibition of AI-2 Biosynthesis in Aeromonas Hydrophila. Molecules 2018, 23, 2627. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ali, F.; Cai, Q.; Yao, Z.; Sun, L.; Lin, W.; Lin, X. Quantitative Proteomic Analysis Reveals That Chemotaxis Is Involved in Chlortetracycline Resistance of Aeromonas Hydrophila. J. Proteom. 2018, 172, 143–151. [Google Scholar] [CrossRef]
- Bentzon-Tilia, M.; Sonnenschein, E.C.; Gram, L. Monitoring and Managing Microbes in Aquaculture – Towards a Sustainable Industry. Microb. Biotechnol. 2016, 9, 576–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skjermo, J.; Vadstein, O. Techniques for Microbial Control in the Intensive Rearing of Marine Larvae. Aquaculture 1999, 177, 333–343. [Google Scholar] [CrossRef]
- Orozco-Medina, C.; Maeda-Martínez, A.M.; López-Cortés, A. Effect of Aerobic Gram-Positive Heterotrophic Bacteria Associated with Artemia Franciscana Cysts on the Survival and Development of Its Larvae. Aquaculture 2002, 213, 15–29. [Google Scholar] [CrossRef]
- Rajabi, S.; Ramazani, A.; Hamidi, M.; Naji, T. Artemia Salina as a Model Organism in Toxicity Assessment of Nanoparticles. DARU, J. Pharm. Sci. 2015, 23, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamza, F.; Kumar, A.R.; Zinjarde, S. Efficacy of Cell Free Supernatant from Bacillus Licheniformis in Protecting Artemia Salina against Vibrio Alginolyticus and Pseudomonas Gessardii. Microb. Pathog. 2018, 116, 335–344. [Google Scholar] [CrossRef]
- Sofos, J.N.; Flick, G.; Nychas, G.-J.; O’Bryan, C.A.; Ricke, S.C.; Crandall, P.G. Meat, Poultry, and Seafood. Food Microbiol. 2014. [Google Scholar] [CrossRef]
- Hamza, F.; Kumar, A.R.; Zinjarde, S. Antibiofilm Potential of a Tropical Marine Bacillus Licheniformis Isolate: Role in Disruption of Aquaculture Associated Biofilms. Aquac. Res. 2016, 47, 2661–2669. [Google Scholar] [CrossRef]
- Padhi, S.; Masi, M.; Chourasia, R.; Rajashekar, Y.; Rai, A.K.; Evidente, A. ADMET Profile and Virtual Screening of Plant and Microbial Natural Metabolites as SARS-CoV-2 S1 Glycoprotein Receptor Binding Domain and Main Protease Inhibitors. Eur. J. Pharmacol. 2021, 890, 173648. [Google Scholar] [CrossRef]
- Mahanta, S.; Chowdhury, P.; Gogoi, N.; Goswami, N.; Borah, D.; Kumar, R.; Chetia, D.; Borah, P.; Buragohain, A.K.; Gogoi, B. Potential Anti-Viral Activity of Approved Repurposed Drug against Main Protease of SARS-CoV-2: An in Silico Based Approach. J. Biomol. Struct. Dyn. 2020, 39, 3802–3811. [Google Scholar] [CrossRef]
- Parvathaneni, V.; Kulkarni, N.S.; Muth, A.; Gupta, V. Drug Repurposing: A Promising Tool to Accelerate the Drug Discovery Process. Drug Discov. Today 2019, 24, 2076–2085. [Google Scholar] [CrossRef]
- Sayed, A.M.; Alhadrami, H.A.; El-Gendy, A.O.; Shamikh, Y.I.; Belbahri, L.; Hassan, H.M.; Abdelmohsen, U.R.; Rateb, M.E. Microbial Natural Products as Potential Inhibitors of SARS-CoV-2 Main Protease (Mpro). Microorganisms 2020, 8, 970. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Name | Zone of Inhibition (mm) | Status |
|---|---|---|
| Staphylococcus aureus | 6 | No inhibitory activity (−) |
| Lactobacillus sp. | 7.5 ± 1 | No inhibitory activity (−) |
| Bacillus thuringiensis | 12 ± 1 | Weak inhibition (+) |
| Klebsiella pneumoniae | 17 ± 1 | Moderate/average inhibition (++) |
| Pseudomonas sp. | 7.33 ± 0.57 | No inhibitory activity (−) |
| Escherichia coli | 6 | No inhibitory activity (−) |
| Name of the Bacteria | Range | Result |
|---|---|---|
| Aeromonas sp. | OD > 4 × ODcutoff | Strong |
| S/N | Ligand No | PubChem CID | Docking Score (Binding Affinity) Kcal/mol |
|---|---|---|---|
| 1 | L-32 | 631096 | −7.1 |
| 2 | L-8 | 9602988 | −6.7 |
| 3 | L-58 | 321710 | −6.4 |
| 4 | L-41 | 139743 | −6.3 |
| 5 | L-37 | 931 | −6.2 |
| 6 | L-17 | 548611 | −6.1 |
| 7 | L-31 | 6054 | −6.0 |
| 8 | L-76 | 545303 | −5.9 |
| SI NO | Compound | Hydrogen Bond | Hydrophobic Bond | ||
|---|---|---|---|---|---|
| Residues | Distance (A°) | Residues | Distance (A°) | ||
| 1. | L-32 | HIS-146 | 1.88 | TRP-208 TYR-213 PHE-124 LYS-122 | 4.82 4.88 4.14 2.89 |
| 2. | L-8 | THR-104 | 3.28 | MET-220 GLU-40 GLU-221 TRP-199 | 5.04 4.19 4.09 5.17 |
| 3. | L-58 | THR-104 | 1.85 | MET-220 | 5.96 |
| 4. | L-41 | SER-243 ASN-244 | 2.74 2.11 | VAL-78 ALA-36 GLU-218 TRP-199 MET-37 MET-27 MET-220 | 5.21 5.38 3.72 3.75 4.95 4.95 |
| 5. | L-37 | Absent | Absent | TRP-199 | 4.45 |
| 6. | L-17 | SER-28 VAL-71 GLY-64 | 2.55 2.20 2.45 | GLN-63 PRO-70 PRO-30 ALA-29 | 2.85 5.28 5.37 3.80 |
| 7. | L-31 | ARG-240 | 2.50 | MET-220 MET-220 TRP-199 ARG-240 | 1.39 5.05 5.05 2.50 |
| 8. | L-76 | TRP-145 | 2.11 | TRP-145 TRP-145 TRP-145 VAL-140 | 3.73 3.73 4.60 4.67 |
| Ligand | L\N | MW g/mol | H. Ac | H. Do | Log Po/w | Log S | Number of Lipinski violations | TPSA (Å2) | Human Intestinal Absorption) | BBB (+ve/−ve) |
|---|---|---|---|---|---|---|---|---|---|---|
| L-32 | 1 | 342.4 | 3 | 2 | 4.26 | −5.03 | Yes; 0 violation | 38.69 | High | +ve |
| L-8 | 2 | 151.16 | 3 | 1 | 2.03 | −2.33 | Yes; 2 violations | 41.82 | High | +ve |
| L-58 | 3 | 133.15 | 1 | 1 | 1.16 | −1.84 | Yes; 2 violations | 29.10 | High | +ve |
| L-41 | 4 | 156.22 | 2 | 0 | 2.79 | −2.37 | Yes; 2 violations | 26.30 | High | +ve |
| L-37 | 5 | 128.17 | 0 | 0 | 3.30 | −3.45 | Yes; 2 violations | 0.00 | Low | +ve |
| L-17 | 6 | 282.12 | 3 | 2 | 3.77 | −4.33 | Yes; 0 violation | 52.82 | High | +ve |
| L-31 | 7 | 122.16 | 1 | 1 | 1.36 | −1.82 | Yes; 2 violations | 20.23 | High | +ve |
| L-76 | 8 | 276.37 | 3 | 0 | 3.81 | −3.82 | Yes; 0 violation | 43.37 | High | +ve |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmud, M.L.; Islam, S.; Biswas, S.; Mortuza, M.G.; Paul, G.K.; Uddin, M.S.; Akhtar-E-Ekram, M.; Saleh, M.A.; Zaman, S.; Syed, A.; et al. Klebsiella pneumoniae Volatile Organic Compounds (VOCs) Protect Artemia salina from Fish Pathogen Aeromonas sp.: A Combined In Vitro, In Vivo, and In Silico Approach. Microorganisms 2023, 11, 172. https://doi.org/10.3390/microorganisms11010172
Mahmud ML, Islam S, Biswas S, Mortuza MG, Paul GK, Uddin MS, Akhtar-E-Ekram M, Saleh MA, Zaman S, Syed A, et al. Klebsiella pneumoniae Volatile Organic Compounds (VOCs) Protect Artemia salina from Fish Pathogen Aeromonas sp.: A Combined In Vitro, In Vivo, and In Silico Approach. Microorganisms. 2023; 11(1):172. https://doi.org/10.3390/microorganisms11010172
Chicago/Turabian StyleMahmud, Md. Liton, Shirmin Islam, Suvro Biswas, Md. Golam Mortuza, Gobindo Kumar Paul, Md. Salah Uddin, Md. Akhtar-E-Ekram, Md. Abu Saleh, Shahriar Zaman, Asad Syed, and et al. 2023. "Klebsiella pneumoniae Volatile Organic Compounds (VOCs) Protect Artemia salina from Fish Pathogen Aeromonas sp.: A Combined In Vitro, In Vivo, and In Silico Approach" Microorganisms 11, no. 1: 172. https://doi.org/10.3390/microorganisms11010172