
What Is Candida Doing in My Food? A Review and Safety Alert on Its Use as Starter Cultures in Fermented Foods

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Physiology and Metabolism
3. Clinical Relevance
4. Isolation and Cultivability
5. Candida Diversity in Cultured Foods and Biotechnological Importance
6. Candida Pathogenicity Evaluation
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Englezos, V.; Rantsiou, K.; Torchio, F.; Rolle, L.; Gerbi, V.; Cocolin, L. Exploitation of the Non-Saccharomyces Yeast Starmerella Bacillaris (Synonym Candida Zemplinina) in Wine Fermentation: Physiological and Molecular Characterizations. Int. J. Food Microbiol. 2015, 199, 33–40. [Google Scholar] [CrossRef]
- Van Wyk, N.; Pretorius, I.S.; von Wallbrunn, C. Assessing the Oenological Potential of Nakazawaea Ishiwadae, Candida Railenensis and Debaryomyces Hansenii Strains in Mixed-Culture Grape Must Fermentation with Saccharomyces Cerevisiae. Fermentation 2020, 6, 49. [Google Scholar] [CrossRef]
- Zohre, D.E.; Erten, H. The Influence of Kloeckera Apiculata and Candida Pulcherrima Yeasts on Wine Fermentation. Process Biochem. 2002, 38, 319–324. [Google Scholar] [CrossRef]
- Samagaci, L.; Ouattara, H.; Niamké, S.; Lemaire, M. Pichia Kudrazevii and Candida Nitrativorans Are the Most Well-Adapted and Relevant Yeast Species Fermenting Cocoa in Agneby-Tiassa, a Local Ivorian Cocoa Producing Region. Food Res. Int. 2016, 89, 773–780. [Google Scholar] [CrossRef]
- Santos, D.S.; Rezende, R.P.; dos Santos, T.F.; de Lima Silva Marques, E.; Ferreira, A.C.R.; de Cerqueira e Silva, A.B.; Romano, C.C.; da Cruz Santos, D.W.; Dias, J.C.T.; Bisneto, J.D.T. Fermentation in Fine Cocoa Type Scavina: Change in Standard Quality as the Effect of Use of Starters Yeast in Fermentation. Food Chem. 2020, 328, 7–12. [Google Scholar] [CrossRef]
- Alloue-Boraud, W.A.M.; N’Guessan, K.F.; Djeni, N.T.; Hiligsmann, S.; Djè, K.M.; Delvigne, F. Fermentation Profile of Saccharomyces Cerevisiae and Candida Tropicalis as Starter Cultures on Barley Malt Medium. J. Food Sci. Technol. 2015, 52, 5236–5242. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Bezawada, J.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Candida Krusei: Biotechnological Potentials and Concerns about Its Safety. Can. J. Microbiol. 2012, 58, 937–952. [Google Scholar] [CrossRef]
- Stefanini, I.; Cavalieri, D. Metagenomic Approaches to Investigate the Contribution of the Vineyard Environment to the Quality of Wine Fermentation: Potentials and Difficulties. Front. Microbiol. 2018, 9, 991. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; De Carvalho Neto, D.P.; Junqueira, A.C.D.O.; Karp, S.G.; Letti, L.A.J.; Magalhães Júnior, A.I.; Soccol, C.R. A Review of Selection Criteria for Starter Culture Development in the Food Fermentation Industry. Food Rev. Int. 2020, 36, 135–167. [Google Scholar] [CrossRef]
- Miranda, L.N.; van der Heijden, I.M.; Costa, S.F.; Sousa, A.P.I.; Sienra, R.A.; Gobara, S.; Santos, C.R.; Lobo, R.D.; Pessoa, V.P.; Levin, A.S. Candida Colonisation as a Source for Candidaemia. J. Hosp. Infect. 2009, 72, 9–16. [Google Scholar] [CrossRef]
- Rajkowska, K.; Kunicka-Styczyńska, A. Typing and Virulence Factors of Food-Borne Candida spp. Isolates. Int. J. Food Microbiol. 2018, 279, 57–63. [Google Scholar] [CrossRef]
- Koh, A.Y. Murine Models of Candida Gastrointestinal Colonization and Dissemination. Eukaryot. Cell 2013, 12, 1416–1422. [Google Scholar] [CrossRef]
- Kieliszek, M.; Kot, A.M.; Bzducha-Wróbel, A.; BŁażejak, S.; Gientka, I.; Kurcz, A. Biotechnological Use of Candida Yeasts in the Food Industry: A Review. Fungal Biol. Rev. 2017, 31, 185–198. [Google Scholar] [CrossRef]
- Schauer, F.; Hanschke, R. Zur Taxonomie Und Ökologie Der Gattung Candida: Taxonomy and Ecology of the Genus Candida. Mycoses 1999, 42, 12–21. [Google Scholar] [CrossRef]
- Ghazi, S.; Rafei, R.; Osman, M.; El Safadi, D.; Mallat, H.; Papon, N.; Dabboussi, F.; Bouchara, J.P.; Hamze, M. The Epidemiology of Candida Species in the Middle East and North Africa. J. Mycol. Med. 2019, 29, 245–252. [Google Scholar] [CrossRef]
- Alby, K.; Bennett, R.J. Sexual Reproduction in the Candida Clade: Cryptic Cycles, Diverse Mechanisms, and Alternative Functions. Cell Mol. Life Sci. 2010, 67, 3275–3285. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Beware of Alternate Names for Candida Species; CDC: Atlanta, GA, USA, 2021; Volume 106. [Google Scholar]
- Mitrea, L.; Ranga, F.; Fetea, F.; Dulf, F.V.; Rusu, A.; Trif, M.; Vodnar, D.C. Biodiesel-Derived Glycerol Obtained from Renewable Biomass—a Suitable Substrate for the Growth of Candida Zeylanoides Yeast Strain ATCC 20367. Microorganisms 2019, 7, 265. [Google Scholar] [CrossRef]
- Garg, M.; Priyanka; Chatterjee, M. Isolation, Characterization and Antibacterial Effect of Biosurfactant from Candida Parapsilosis. Biotechnol. Rep. 2018, 18, e00251. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Bezawada, J.; Ajila, C.M.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Mixed Culture of Kluyveromyces Marxianus and Candida Krusei for Single-Cell Protein Production and Organic Load Removal from Whey. Bioresour. Technol. 2014, 164, 119–127. [Google Scholar] [CrossRef]
- Douglass, A.P.; Offei, B.; Braun-Galleani, S.; Coughlan, A.Y.; Martos, A.A.R.; Ortiz-Merino, R.A.; Byrne, K.P.; Wolfe, K.H. RESEARCH ARTICLE Population Genomics Shows No Distinction between Pathogenic Candida Krusei and Environmental Pichia Kudriavzevii: One Species, Four Names. PLoS Pathog. 2018, 14, e1007138. [Google Scholar] [CrossRef] [Green Version]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of Novel Killer Toxins Secreted by Wine-Related Non-Saccharomyces Yeasts and Their Action on Brettanomyces spp. Int. J. Food Microbiol. 2014, 188, 83–91. [Google Scholar] [CrossRef]
- Büyüksırıt Bedir, T.; Kuleaşan, H. A Natural Approach, the Use of Killer Toxin Produced by Metschnikowia Pulcherrima in Fresh Ground Beef Patties for Shelf Life Extention. Int. J. Food Microbiol. 2021, 345, 109154. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef]
- Jacobsen, I.D.; Niemiec, M.J.; Kapitan, M.; Polke, M. Commensal to Pathogen Transition of Candida Albicans. Encycl. Mycol. 2021, 1, 507–525. [Google Scholar] [CrossRef]
- Hurley, D.L.; Fauci, A.S. Disseminated Candidiasis. I. An Experimental Model in the Guinea Pig. J. Infect. Dis. 1975, 131, 516–521. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, S.; Manichanh, C.; Nielsen, T.; Pons, N.; Yamada, T.; Mende, D.R.; et al. A Human Gut Microbial Gene Catalog Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.; Niu, S.; Zhang, C.; Xu, X.; Qin, M.; Huang, S.; Zhang, L. Epidemiology, Antifungal Susceptibilities, and Risk Factors for Invasive Candidiasis from 2011 to 2013 in a Teaching Hospital in Southwest China. J. Microbiol. Immunol. Infect. 2017, 50, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Strollo, S.; Lionakis, M.S.; Adjemian, J.; Steiner, C.A.; Prevots, D.R. Epidemiology of Hospitalizations Associated with Invasive Candidiasis, United States, 2002–2012. Emerg. Infect. Dis. 2017, 23, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.Q.; Tseng, K.Y.; Tsai, Y.H. Candida Gut Commensalism and Inflammatory Disease. Med. Microecol. 2020, 3, 100008. [Google Scholar] [CrossRef]
- Austermeier, S.; Kasper, L.; Westman, J.; Gresnigt, M.S. I Want to Break Free—Macrophage Strategies to Recognize and Kill Candida Albicans, and Fungal Counter-Strategies to Escape. Curr. Opin. Microbiol. 2020, 58, 15–23. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida Glabrata, Candida Parapsilosis and Candida Tropicalis: Biology, Epidemiology, Pathogenicity and Antifungal Resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [PubMed]
- Aaron, L.; Torsten, M. Candida Albicans in Celiac Disease: A Wolf in Sheep’s Clothing. Autoimmun. Rev. 2020, 19, 102621. [Google Scholar] [CrossRef] [PubMed]
- Bacher, P.; Hohnstein, T.; Beerbaum, E.; Röcker, M.; Blango, M.G.; Kaufmann, S.; Röhmel, J.; Eschenhagen, P.; Grehn, C.; Seidel, K.; et al. Human Anti-Fungal Th17 Immunity and Pathology Rely on Cross-Reactivity against Candida Albicans. Cell 2019, 176, 1340–1355.e15. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The Microbiota in Adaptive Immune Homeostasis and Disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Pérez, J.C. Fungi of the Human Gut Microbiota: Roles and Significance. Int. J. Med. Microbiol. 2021, 311, 151490. [Google Scholar] [CrossRef]
- Huët, M.A.L.; Lee, C.Z.; Rahman, S. A Review on Association of Fungi with the Development and Progression of Carcinogenesis in the Human Body. Curr. Res. Microb. Sci. 2022, 3, 100090. [Google Scholar] [CrossRef]
- Musumeci, S.; Coen, M.; Leidi, A.; Schrenzel, J. The Human Gut Mycobiome and the Specific Role of Candida Albicans: Where Do We Stand, as Clinicians? Clin. Microbiol. Infect. 2022, 28, 58–63. [Google Scholar] [CrossRef]
- Iliev, I.D.; Leonardi, I. Fungal Dysbiosis: Immunity and Interactions at Mucosal Barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef]
- De Luca, C.; Guglielminetti, M.; Ferrario, A.; Calabrò, M.; Casari, E. Candidemia: Species Involved, Virulence Factors and Antimycotic Susceptibility. New Microbiol. 2012, 35, 459–468. [Google Scholar]
- Pfaller, M.; Neofytos, D.; Diekema, D.; Azie, N.; Meier-Kriesche, H.U.; Quan, S.P.; Horn, D. Epidemiology and Outcomes of Candidemia in 3648 Patients: Data from the Prospective Antifungal Therapy (PATH Alliance®) Registry, 2004-2008. Diagn. Microbiol. Infect. Dis. 2012, 74, 323–331. [Google Scholar] [CrossRef]
- Hirayama, T.; Miyazaki, T.; Ito, Y.; Wakayama, M.; Shibuya, K.; Yamashita, K.; Takazono, T.; Saijo, T.; Shimamura, S.; Yamamoto, K.; et al. Virulence Assessment of Six Major Pathogenic Candida Species in the Mouse Model of Invasive Candidiasis Caused by Fungal Translocation. Sci. Rep. 2020, 10, 3814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome Definition Re-Visited: Old Concepts and New Challenges Concepts and New Challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bansal, A.; Chakrabarti, A.; Singhi, S. Evaluation of Efficacy of Probiotics in Prevention of Candida Colonization in a PICU-a Randomized Controlled Trial. Crit. Care Med. 2013, 41, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Precup, G.; Teleky, B.E.; Ranga, F.; Vodnar, D.C. Assessment of Physicochemical and Rheological Properties of Xylo-Oligosaccharides and Glucose-Enriched Doughs Fermented with BB-12. Biology 2022, 11, 553. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Wang, D.; Li, C.; Wang, Y.; Diao, Y.; Tang, Y. Isolation, Identification and Genotyping of Candida Albicans from Landes Geese. Transbound. Emerg. Dis. 2022, 69, 349–359. [Google Scholar] [CrossRef]
- Idelevich, E.A.; Grunewald, C.M.; Wüllenweber, J.; Becker, K. Rapid Identification and Susceptibility Testing of Candida spp. from Positive Blood Cultures by Combination of Direct MALDI-TOF Mass Spectrometry and Direct Inoculation of Vitek 2. PLoS ONE 2014, 9, e114834. [Google Scholar] [CrossRef]
- Sourav, D.; Shreya, S.; Yamini, T.; Chakrabarti, A.; Rudramurthy, S.; Harsimran, K.; Shamanth, A.S.; Ghosh, A. A Selective Medium for Isolation and Detection of Candida. J. Clin. Microbiol. 2021, 59, e00326-20. [Google Scholar]
- Odds, F.C.; Bernaerts, R. CHROMagar Candida, a New Differential Isolation Medium for Presumptive Identification of Clinically Important Candida Species. J. Clin. Microbiol. 1994, 32, 1923–1929. [Google Scholar] [CrossRef]
- Byadarahally Raju, S.; Rajappa, S. Isolation and Identification of Candida from the Oral Cavity. ISRN Dent. 2011, 2011, 487921. [Google Scholar] [CrossRef]
- Gasparoto, T.H.; Dionísio, T.J.; De Oliveira, C.E.; Porto, V.C.; Gelani, V.; Santos, C.F.; Campanelli, A.P.; Lara, V.S. Isolation of Candida Dubliniensis from Denture Wearers. J. Med. Microbiol. 2009, 58, 959–962. [Google Scholar] [CrossRef]
- Bernal, S.; Mazuelos, E.M.; García, M.; Aller, A.I.; Martínez, M.A.; Gutiérrez, M.J. Evaluation of CHROMagar Candida Medium for the Isolation and Presumptive Identification of Species of Candida of Clinical Importance. Diagn. Microbiol. Infect. Dis. 1996, 24, 201–204. [Google Scholar] [CrossRef]
- Manfredi, M.; McCullough, M.J.; Al-Karaawi, Z.M.; Hurel, S.J.; Porter, S.R. The Isolation, Identification and Molecular Analysis of Candida spp. Isolated from the Oral Cavities of Patients with Diabetes Mellitus. Oral Microbiol. Immunol. 2002, 17, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Welsh, R.M.; Bentz, M.L.; Shams, A.; Houston, H.; Lyons, A.; Rose, L.J.; Litvintseva, P. Survival, Persistence, and Isolation of the Emerging Multidrug-Resistant Pathogenic Yeast Candida Auris on a Plastic Health Care Surface. J. Clin. Microbiol. 2017, 55, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Horvath, L.L.; Hospenthal, D.R.; Murray, C.K.; Dooley, D.P. Direct Isolation of Candida spp. from Blood Cultures on the Chromogenic Medium CHROMagar Candida. J. Clin. Microbiol. 2003, 41, 2629–2632. [Google Scholar] [CrossRef] [PubMed]
- Eraso, E.; Moragues, M.D.; Villar-Vidal, M.; Sahand, I.H.; González-Gómez, N.; Pontón, J.; Quindós, G. Evaluation of the New Chromogenic Medium Candida ID 2 for Isolation and Identification of Candida Albicans and Other Medically Important Candida Species. J. Clin. Microbiol. 2006, 44, 3340–3345. [Google Scholar] [CrossRef]
- Darwazeh, A.M.-G.; Sulieman, A.-R.; Al-Mojaiwe, S. Isolation of Candida Species from the Oral Cavity and Fingertips of Complete Denture Wearers. J. Prosthet. Dent. 2001, 86, 420–423. [Google Scholar] [CrossRef]
- Nnadi, N.E.; Ayanbimpe, G.M.; Okolo, M.O.; Enweani, I.B.; Criseo, G.; Romeo, O. Isolation and Molecular Characterization of Candida Africana from Jos Nigeria. Med. Mycol. 2012, 50, 765–767. [Google Scholar] [CrossRef]
- Álvarez, M.I.; Suárez, B.L.; Caicedo, L.D. Isolation of Candida Dubliniensis for the First Time in Cali, Colombia, and Its Identification with Phenotyping Methods. Mycopathologia 2009, 167, 19–24. [Google Scholar] [CrossRef]
- Lotfali, E.; Mardani, M.; Abolghasemi, S.; Darvishnia, D.; Rabiei, M.M.; Ghasemi, R.; Fattahi, A. Isolation of Candida Africana in Oral Candidiasis: First Report among Cancer Patients in Iran. Curr. Med. Mycol. 2020, 6, 58–62. [Google Scholar] [CrossRef]
- Adjapong, G.; Bartlett, M.; Hale, M.; Garrill, A. The Isolation of Candida Rugosa and Candida Mesorugosa from Clinical Samples in Ghana. Med. Mycol. 2016, 54, 322–326. [Google Scholar] [CrossRef]
- Khan, Z.; Ahmad, S.; Joseph, L.; Al-Obaid, K. Isolation of Cholesterol-Dependent, Multidrug-Resistant Candida Glabrata Strains from Blood Cultures of a Candidemia Patient in Kuwait. BMC Infect. Dis. 2014, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Okulicz, J.F.; Rivard, R.G.; Conger, N.G.; Nguyen, M.X.; Hospenthal, D.R. Primary Isolation of Candida Species from Urine Specimens Using Chromogenic Medium. Mycoses 2008, 51, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Krishnasamy, L.; Krishnakumar, S.; Santharam, P.; Saikumar, C. Isolation and Identification of Candida Species in Patients with Vulvovaginal Candidiasis. J. Pure Appl. Microbiol. 2018, 12, 2269–2273. [Google Scholar] [CrossRef]
- Taj-Aldeen, S.J.; Al-Ansari, H.I.; Boekhout, T.; Theelen, B. Co-Isolation of Trichosporon Inkin and Candida Parapsilosis from a Scalp White Piedra Case. Med. Mycol. 2004, 42, 87–92. [Google Scholar] [CrossRef]
- De Sousa, L.V.N.F.; Santos, V.L.; de Souza Monteiro, A.; Dias-Souza, M.V.; Marques, S.G.; de Faria, E.S.; de O. Assunção, E.A.; dos Santos, S.G.; Zonis, J.M.; de Alvarenga, D.G.; et al. Isolation and Identification of Candida Species in Patients with Orogastric Cancer: Susceptibility to Antifungal Drugs, Attributes of Virulence in Vitro and Immune Response Phenotype. BMC Infect. Dis. 2016, 16, 86. [Google Scholar] [CrossRef]
- Sharma, U.; Patel, K.; Shah, V.; Sinha, S.; Rathore, V.P.S. Isolation and Speciation of Candida in Type Ii Diabetic Patients Using Chrom Agar: A Microbial Study. J. Clin. Diagn. Res. 2017, 11, DC09–DC11. [Google Scholar] [CrossRef]
- Kanna, B.; Kumar, G.; Swapna, M.; Easow, J.M. Isolation and Identification of Candida Species from Various Clinical Samples in a Tertiary Care Hospital. Int. J. Res. Med. Sci. 2017, 5, 3520. [Google Scholar] [CrossRef]
- Alonso-Vargas, R.; Elorduy, L.; Eraso, E.; Cano, J.F.; Guarro, J.; Pontón, J.; Quindós, G. Isolation of Candida Africana, Probable Atypical Strains of Candida Albicans, from a Patient with Vaginitis. Med. Mycol. 2008, 46, 167–170. [Google Scholar] [CrossRef]
- Gauch, L.M.R.; Pedrosa, S.S.; Silveira-Gomes, F.; Esteves, R.A.; Marques-da-Silva, S.H. Isolation of Candida spp. from Denture-Related Stomatitis in Pará Brazil. Braz. J. Microbiol. 2018, 49, 148–151. [Google Scholar] [CrossRef]
- Pekard-Amenitsch, S.; Schriebl, A.; Posawetz, W.; Willinger, B.; Kölli, B.; Buzina, W. Isolation of Candida Auris from Ear of Otherwise Healthy Patient, Austria, 2018. Emerg. Infect. Dis. 2018, 24, 1596–1597. [Google Scholar] [CrossRef]
- Carrasco, L.; Ramos, M.; Galisteo, R.; Pisa, D.; Fresno, M.; González, M.E. Isolation of Candida Famata from a Patient with Acute Zonal Occult Outer Retinopathy. J. Clin. Microbiol. 2005, 43, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Ghajari, A.; Lotfali, E.; Ahmadi, N.A.; Fassihi, P.N.; Shahmohammadi, N.; Ansari, S.; Norouzi, M.; Arab-Mazar, Z. Isolation of Different Species of Candida in Patients with Vulvovaginal Candidiasis from Damavand, Iran. Arch. Clin. Infect. Dis. 2018, 13, e59291. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.P.; Zinchuk, R.; Larone, D.H. CHROMagar Candida as the Sole Primary Medium for Isolation of Yeasts and as a Source Medium for the Rapid-Assimilation-of-Trehalose Test. J. Clin. Microbiol. 2005, 43, 1210–1212. [Google Scholar] [CrossRef] [PubMed]
- Ghelardi, E.; Pichierri, G.; Castagna, B.; Barnini, S.; Tavanti, A.; Campa, M. Efficacy of Chromogenic Candida Agar for Isolation and Presumptive Identification of Pathogenic Yeast Species. Clin. Microbiol. Infect. 2008, 14, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Giammanco, G.M.; Melilli, D.; Pizzo, G. Candida Pararugosa Isolation from the Oral Cavity of an Italian Denture Wearer. Res. Microbiol. 2004, 155, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Chattopadhyay, U.; Naskar, K. Prevalence of Candida Albicans in Raw Chicken and Mutton Meat Samples Sold in the Open Markets of Kolkata City of West Bengal. Int. J. Livest. Res. 2017, 7, 243–249. [Google Scholar] [CrossRef]
- De Casia Dos Santos, R.; Marin, J.M. Isolation of Candida spp. from Mastitic Bovine Milk in Brazil. Mycopathologia 2005, 159, 251–253. [Google Scholar] [CrossRef]
- Eldesouky, I.; Mohamed, N.; Khalaf, D.; Salama, A.; Elsify, A.; Ombarak, R.; El-Ballal, S.; Effat, M.; Al Shabrawy, M. Candida Mastitis in Dairy Cattle with Molecular Detection of Candida Albicans. Kafkas Univ. Vet. Fak. Derg. 2016, 22, 461–464. [Google Scholar] [CrossRef]
- Ramin, A.G.; Shoorijeh, S.J.; Ashtiani, H.R.A.; Naderi, M.M.; Behzadi, M.A.; Tamadon, A.; Sofi, A.H.; Wani, S.A.; Salahuddin, M.; Malik, A.H.; et al. Removal of Metallic Objects from Animal Feeds: Development and Studies on a New Machine. VetScan 2008, 3, 1–6. [Google Scholar]
- Zhai, L.; Zhou, Y.; Wu, Y.; Jin, Y.; Zhu, Q.; Gao, S.; Li, X.; Sun, Z.; Xiao, Y.; Huang, B.; et al. Isolation and Identification of Candida Tropicalis in Sows with Fatal Infection: A Case Report. BMC Vet. Res. 2021, 17, 108. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, Q.; Xue, W.; Li, Y.; Guo, Y.; Wu, X.; Huo, S.; Li, Y.; Li, C. The Isolation and Identification of Candida Glabrata from Avian Species and a Study of the Antibacterial Activities of Chinese Herbal Medicine in Vitro. Poult. Sci. 2021, 100, 101003. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Piergili Fioretti, D.; Boncio, L.; Pasquali, P.; Del Rossi, E. Isolation of Candida Rugosa from Turkeys. J. Vet. Med. Ser. B 2000, 47, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.; Sulieman, A.M.E.; Elkhalifa, E.A. Isolation Identification and Characterization of Candida Utilis from Some of the Sudanese Traditionally Fermented Food Products Isolation. Gezira J. Eng. Appl. 2015, 4, 104–114. [Google Scholar]
- Arora, P.; Singh, P.; Wang, Y.; Yadav, A.; Pawar, K.; Singh, A.; Padmavati, G.; Chowdhary, A.; Xu, J. Environmental Isolation of Candida Auris from the Coastal Wetlands of Andaman Islands, India. mBio 2021, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cook, W.L.; Schlitzer, R.L. Isolation of Candida Albicans from Freshwater and Sewage. Appl. Environ. Microbiol. 1981, 41, 840–842. [Google Scholar] [CrossRef] [PubMed]
- Alwaely, W.A.S.; Ghadban, A.K.; Alrubayae, I.M.N. Production and Properties of Biosurfactant from the Local Isolation of Candida spp. Drug Invent. Today 2019, 12, 948–953. [Google Scholar] [CrossRef]
- Patel, M.J.; Tipre, D.R.; Dave, S.R. Isolation and Identification of a Candida Digboiensis Strain from an Extreme Acid Mine Drainage of the Lignite Mine, Gujarat. J. Basic Microbiol. 2009, 49, 564–571. [Google Scholar] [CrossRef]
- Wang, L.; Chi, Z.; Wang, X.; Ju, L.; Chi, Z.; Guo, N. Isolation and Characterization of Candida Membranifaciens Subsp. Flavinogenie W14-3, a Novel Riboflavin-Producing Marine Yeast. Microbiol. Res. 2008, 163, 255–266. [Google Scholar] [CrossRef]
- Pinjon, E.; Sullivan, D.; Salkin, I.; Shanley, D.; Coleman, D. Simple, Inexpensive, Reliable Method for Differentiation of Candida Dubliniensis from Candida Albicans. J. Clin. Microbiol. 1998, 36, 2093–2095. [Google Scholar] [CrossRef]
- Ibrahim, A.; Peyclit, L.; Abdallah, R.; Khelaifia, S.; Chamieh, A.; Rolain, J.M.; Bittar, F. Sca Medium: A New Culture Medium for the Isolation of All Candida Auris Clades. J. Fungi 2021, 7, 433. [Google Scholar] [CrossRef]
- Sahand, I.H.; Moragues, M.D.; Eraso, E.; Villar-Vidal, M.; Quindós, G.; Pontón, J. Supplementation of CHROMagar Candida Medium with Pal’s Medium for Rapid Identification of Candida Dubliniensis. J. Clin. Microbiol. 2005, 43, 5768–5770. [Google Scholar] [CrossRef] [PubMed]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on Major Food-Borne Zoonotic Bacterial Pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef] [PubMed]
- Käferstein, F.K. Food Safety: The Fourth Pillar in the Strategy to Prevent Infant Diarrhoea. Bull. World Health Organ. 2003, 81, 842–843. [Google Scholar]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpędowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J. Chemical Signaling and Insect Attraction Is a Conserved Trait in Yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef]
- Vadkertiová, R.; Molnárová, J.; Vránová, D.; Sláviková, E. Yeasts and Yeast-like Organisms Associated with Fruits and Blossoms of Different Fruit Trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Freilich, S.; Bartuv, R.; Zhimo, V.Y.; Kumar, A.; Biasi, A.; Salim, S.; Feygenberg, O.; Burchard, E.; Dardick, C.; et al. Global Analysis of the Apple Fruit Microbiome: Are All Apples the Same? Environ. Microbiol. 2021, 23, 6038–6055. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Gond, S.K.; Kumar, A.; Sharma, V.K.; Verma, S.K.; Kharwar, R.N.; Sieber, T.N. Season and Tissue Type Affect Fungal Endophyte Communities of the Indian Medicinal Plant Tinospora Cordifolia More Strongly than Geographic Location. Microb. Ecol. 2012, 64, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Zhimo, Y.; Biasi, A.; Salim, S.; Feygenberg, O.; Wisniewski, M.; Droby, S. Endophytic Microbiome in the Carposphere and Its Importance in Fruit Physiology and Pathology. In Postharvest Pathology; Spadaro, D., Droby, S., Gullino, M.L., Eds.; Springer: Cham, Switzerland, 2021; pp. 73–88. ISBN 9783030565305. [Google Scholar]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R.; Fukami, T.; Re, I.; Sheppard, J.; et al. The Ecology of Insect—Yeast Relationships and Its Relevance to Human Industry. Proc. R. Soc. B 2018, 285, 20172733. [Google Scholar] [CrossRef]
- Jones, R.; Fountain, M.T.; Andreani, N.A.; Günther, C.S.; Goddard, M.R. The Relative Abundances of Yeasts Attractive to Drosophila Suzukii Differ between Fruit Types and Are Greatest on Raspberries. Sci. Rep. 2022, 12, 10382. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Medina, E.; Ruiz-Bellido, M.Á.; Romero-Gil, V.; Montes-Borrego, M.; Landa, B.B. Enhancement of the Knowledge on Fungal Communities in Directly Brined Aloreña de Málaga Green Olive Fermentations by Metabarcoding Analysis. PLoS ONE 2016, 11, e0163135. [Google Scholar] [CrossRef]
- Abdo, H.; Catacchio, C.R.; Ventura, M.; D’Addabbo, P.; Alexandre, H.; Guilloux-Bénatier, M.; Rousseaux, S. The Establishment of a Fungal Consortium in a New Winery. Sci. Rep. 2020, 10, 7962. [Google Scholar] [CrossRef] [PubMed]
- Palareti, G.; Legnani, C.; Cosmi, B.; Antonucci, E.; Erba, N.; Poli, D.; Testa, S.; Tosetto, A. Comparison between Different D-Dimer Cutoff Values to Assess the Individual Risk of Recurrent Venous Thromboembolism: Analysis of Results Obtained in the DULCIS Study. Int. J. Lab. Hematol. 2016, 38, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Candida Zemplinina Sp. Nov., an Osmotolerant and Psychrotolerant Yeast That Ferments Sweet Botrytized Wines. Int. J. Syst. Evol. Microbiol. 2003, 53, 2079–2083. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Z.; Long, F.; Guo, C.; Yuan, Y.; Yue, T. Detection of Zygosaccharomyces Rouxii and Candida Tropicalis in a High-Sugar Medium by a Metal Oxide Sensor-Based Electronic Nose and Comparison with Test Panel Evaluation. J. Food Prot. 2015, 78, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, A.S.S. Fermentation of Fruit Juices by the Osmotolerant Yeast Candida Magnoliae. Master’s Dissertation, Universidade Nova de Lisboa, Lisbon, Portugal, 2014. [Google Scholar]
- Qi, W.; Zhang, W.T.; Lu, F.P. Carbon Metabolism and Transcriptional Variation in Response to Salt Stress in the Genome Shuffled Candida Versatilis and a Wild-Type Salt Tolerant Yeast Strain. RSC Adv. 2017, 7, 1646–1653. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.-H.; Lee, D.-H.; Park, Y.-C.; Lee, M.-G.; Kim, D.-O.; Ryu, Y.-W.; Seo, J.-H. Proteomic Analysis of Fructophilic Properties of Osmotolerant Candida Magnoliae. J. Microbiol. Biotechnol. 2008, 18, 248–254. [Google Scholar]
- Raymond Eder, M.L.; Rosa, A.L. Genetic, Physiological, and Industrial Aspects of the Fructophilic Non-Saccharomyces Yeast Species, Starmerella Bacillaris. Fermentation 2021, 7, 87. [Google Scholar] [CrossRef]
- Viesser, J.A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Rogez, H.; Góes-Neto, A.; Azevedo, V.; Brenig, B.; Aburjaile, F.; Soccol, C.R. Co-Culturing Fructophilic Lactic Acid Bacteria and Yeast Enhanced Sugar Metabolism and Aroma Formation during Cocoa Beans Fermentation. Int. J. Food Microbiol. 2021, 339, 109015. [Google Scholar] [CrossRef]
- De Oliveira Junqueira, A.C.; de Melo Pereira, G.V.; Viesser, J.A.; de Carvalho Neto, D.P.; Querne, L.B.P.; Soccol, C.R. Isolation and Selection of Fructose-Consuming Lactic Acid Bacteria Associated with Coffee Bean Fermentation. Food Biotechnol. 2022, 36, 58–75. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Studies on Acetate Ester Production by Non-Saccharomyces Wine Yeasts. Int. J. Food Microbiol. 2001, 70, 283–289. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; Van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces Cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Masneuf-Pomarede, I.; Juquin, E.; Miot-Sertier, C.; Renault, P.; Laizet, Y.; Salin, F.; Alexandre, H.; Capozzi, V.; Cocolin, L.; Colonna-Ceccaldi, B.; et al. The Yeast Starmerella Bacillaris (Synonym Candida Zemplinina) Shows High Genetic Diversity in Winemaking Environments. FEMS Yeast Res. 2015, 15, fov045. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-maréchal, R.; Morge, C.; Sparrow, C. Non-Saccharomyces Yeasts Nitrogen Source Preferences: Impact on Sequential Fermentation and Wine Volatile Compounds Profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef]
- Perpetuini, G.; Rossetti, A.P.; Tittarelli, F.; Battistelli, N.; Arfelli, G.; Suzzi, G.; Tofalo, R. Promoting Candida Zemplinina Adhesion on Oak Chips: A Strategy to Enhance Esters and Glycerol Content of Montepulciano d’Abruzzo Organic Wines. Food Res. Int. 2021, 150, 110772. [Google Scholar] [CrossRef]
- Tufariello, M.; Capozzi, V.; Spano, G.; Cantele, G.; Venerito, P.; Mita, G.; Grieco, F. Effect of Co-Inoculation of Candida Zemplinina, Saccharomyces Cerevisiae and Lactobacillus Plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy). Microorganisms 2020, 8, 726. [Google Scholar] [CrossRef]
- Mahazar, N.H.; Sufian, N.F.; Meor Hussin, A.S.; Norhayati, H.; Mathawan, M.; Rukayadi, Y. Candida Sp. as a Starter Culture for Cocoa (Theobroma Cacao L.) Beans Fermentation. Int. Food Res. J. 2015, 22, 1783–1787. [Google Scholar]
- Bressani, A.P.P.; Martinez, S.J.; de Figueiredo Vilela, L.; Dias, D.R.; Schwan, R.F. Coffee Protein Profiles during Fermentation Using Different Yeast Inoculation Methods. Pesqui. Agropecuária Bras. 2020, 55, e01159. [Google Scholar] [CrossRef]
- Bressani, A.P.P.; Martinez, S.J.; Sarmento, A.B.I.; Borém, F.M.; Schwan, R.F. Organic Acids Produced during Fermentation and Sensory Perception in Specialty Coffee Using Yeast Starter Culture. Food Res. Int. 2020, 128, 108773. [Google Scholar] [CrossRef]
- Bressani, A.P.P.; Martinez, S.J.; Evangelista, S.R.; Dias, D.R.; Schwan, R.F. Characteristics of Fermented Coffee Inoculated with Yeast Starter Cultures Using Different Inoculation Methods. LWT Food Sci. Technol. 2018, 92, 212–219. [Google Scholar] [CrossRef]
- Sulistyaningtyas, A.R.; Lunggani, A.T.; Kusdiyantini, E. Kefir Produced from Red Rice Milk by Lactobacillus Bulgaricus and Candida Kefir Starter. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012038. [Google Scholar] [CrossRef]
- Vasile, M.-A.; Cotârlet, M.; Bahrim, G.-E. Metabolic Functionality of the Synergistically Ameliorated Consortium Based on Wild Kefir Grains and Selected Microorganisms for Bovine Colostrum Fermentation. Innov. Rom. Food Biotechnol. 2020, 19, 33–38. [Google Scholar]
- Biagiotti, C.; Ciani, M.; Canonico, L.; Comitini, F. Occurrence and Involvement of Yeast Biota in Ripening of Italian Fossa Cheese. Eur. Food Res. Technol. 2018, 244, 1921–1931. [Google Scholar] [CrossRef]
- Lanza, B.; Di Marco, S.; Bacceli, M.; Di Serio, M.G.; Di Loreto, G.; Cellini, M.; Simone, N. Lactiplantibacillus Plantarum Used as Single, Multiple, and Mixed Starter Combined with Candida Boidinii for Table Olive Fermentations: Chemical, Textural, and Sensorial Characterization of Final Products. Fermentation 2021, 7, 239. [Google Scholar] [CrossRef]
- Purriños, L.; Carballo, J.; Lorenzo, J.M. The Influence of Debaryomyces Hansenii, Candida Deformans and Candida Zeylanoides on the Aroma Formation of Dry-Cured “Lacón”. Meat Sci. 2013, 93, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.Y.; Chua, J.Y.; Liu, S.Q. Exploring the Feasibility of Biotransforming Salted Soy Whey into a Soy Sauce-like Condiment Using Wine Yeast Torulaspora Delbrueckii and Soy Sauce Yeasts Zygosaccharomyces Rouxii and Candida Versatilis as Single Starter Cultures. Food Res. Int. 2022, 156, 111350. [Google Scholar] [CrossRef]
- Silva, C.F.; Vilela, D.M.; de Souza Cordeiro, C.; Duarte, W.F.; Dias, D.R.; Schwan, R.F. Evaluation of a Potential Starter Culture for Enhance Quality of Coffee Fermentation. World J. Microbiol. Biotechnol. 2013, 29, 235–247. [Google Scholar] [CrossRef]
- Hay, R.J. Fungal Infections. Clin. Dermatol. 2006, 24, 201–212. [Google Scholar] [CrossRef]
- Samaranayake, Y.H.; MacFarlane, T.W.; Samaranayake, L.P.; Aitchison, T. The in Vitro Proteolytic and Saccharolytic Activity of Candida Species Cultured in Human Saliva. Oral Microbiol. Immunol. 1994, 9, 229–235. [Google Scholar] [CrossRef]
- Segal, E.; Gottlieb, S.; Altboum, Z.; Gov, Y.; Berdicevsky, I. Adhesion of Candida Albicans to Epithelial Cells—Effect of Nikkomycin. Mycoses 1997, 40, 33–39. [Google Scholar] [CrossRef]
- Jin, Y.; Yip, H.K.; Samaranayake, Y.H.; Yau, J.Y.; Samaranayake, L.P. Biofilm-Forming Ability of Candida Albicans Is Unlikely to Contribute to High Levels of Oral Yeast Carriage in Cases of Human Immunodeficiency Virus Infection. J. Clin. Microbiol. 2003, 41, 2961–2967. [Google Scholar] [CrossRef]
- Naglik, J.R.; Rodgers, C.A.; Shirlaw, P.J.; Dobbie, J.L.; Fernandes-Naglik, L.L.; Greenspan, D.; Agabian, N.; Challacombe, S.J. Differential Expression of Candida Albicans Secreted Aspartyl Proteinase and Phospholipase B Genes in Humans Correlates with Active Oral and Vaginal Infections. J. Infect. Dis. 2003, 188, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Price, M.F.; Wilkinson, A.N.D.; Gentry, L.O. Plate Method for Detection of Propspholipase Acvivity in Candida Albicans. Sabouraudia 1982, 20, 7–14. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, E.M.; Monteiro, L.C.; da Silva Fernandes, M.B.; Martelli Junior, H.; Bonan, P.R.F.; Nobre, S.A.M. Antifungal Susceptibility in Vitro Determined by the Etest® for Candida Obtained from the Oral Cavity of Irradiated and Elderly Individuals. Braz. Dent. J. 2015, 26, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida Albicans Pathogenicity Mechanisms. Virulence 2018, 4, 119–128. [Google Scholar] [CrossRef]
- Yang, H.; Tsang, P.C.S.; Pow, E.H.N.; Lam, O.L.T.; Tsang, P.W.K. Potential Role of Candida Albicans Secreted Aspartic Protease 9 in Serum Induced-Hyphal Formation and Interaction with Oral Epithelial Cells. Microb. Pathog. 2020, 139, 103896. [Google Scholar] [CrossRef]
- Brieland, J.; Essig, D.; Jackson, C.; Frank, D.; Loebenberg, D.; Menzel, F.; Arnold, B.; DiDomenico, B.; Hare, R. Comparison of Pathogenesis and Host Immune Responses to Candida Glabrata and Candida Albicans in Systemically Infected Immunocompetent Mice. Infect. Immun. 2001, 69, 5046–5055. [Google Scholar] [CrossRef] [Green Version]


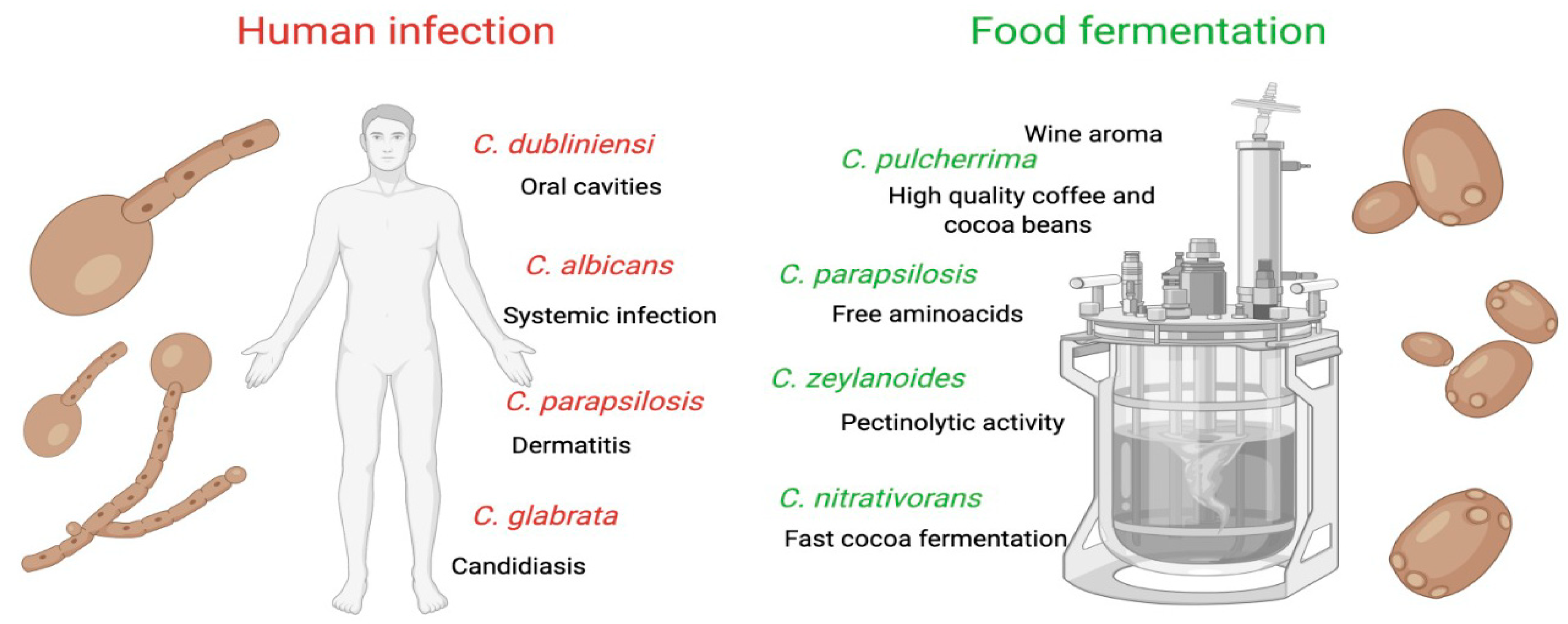{kind=link}
| Species | Isolation Source | Medium | Cultivation Conditions | References |
|---|---|---|---|---|
| Clinical Samples | ||||
| C. dubliniensis | Denture wearers | SAB and CAC | Room temperature | [51] |
| C. albicans, N. (Candida)glabrata, P. (Candida)krusei C. tropicalis, M. (Candida)parapsilosis, C. (Candida) lusitaniae, M. (Candida) guilliermondii, D. (Candida)famata, C. lambica, K. (Candida) zeylanoides, and Candida humicola (teleomorph Vanrija humicola) | Clinical samples | SAB and CAC | 48 h, 37 °C | [52] |
| Candida spp. | Human oral cavities | SAB with chloramphenicol; | NI | [53] |
| C. auris | Clinical samples | CAC | 40 °C | [54] |
| Candida spp. | Oral cavity | SDA and CAC | 24–48 h, 37 °C | [50] |
| C. dubliniensis | Denture stomatitis | Candida ID2 | 48 h,37 °C | [51] |
| Candida spp. | Blood sample | CAC | 30 °C | [55] |
| C. albicans, C. dubliniensis, C. tropicalis, and P. (Candida)krusei | Clinical samples | Candida ID, Albicans ID 2, CAC | 10 days, 37 °C | [56] |
| Candida spp. | Oral cavity | SAB | 48 h, 37 °C | [57] |
| C. africana | Vagina sample | CAC | NI | [58] |
| C. dubliniensis | Oral cavity | CAC | 48 h, 30 °C | [59] |
| C. africana | Oral candidiasis sample | SAB | 24 h, 30 °C | [60] |
| C. rugosa (teleomorph Diutina rugosa) and C. mesorugosa (teleomorph Diutina mesorugosa) | Clinical samples | SAB and brilliance Candida agar | NI | [61] |
| N. (Candida)glabrata | Blood samples | Blood agar, chocolate agar, SAB, and MH agar | NI | [62] |
| C. albicans, P. (Candida)krusei, C. tropicalis, and N. (Candida)glabrata | Urine | CAC, McConkey and blood agar | 48 h, 30–37 °C | [63] |
| Candida spp. | Vagina | SAB and Hichrome Candida differential agar | 24–48 h, 37 °C | [64] |
| M. (Candida)parapsilosis | Scalp | SAB with streptomycin, SAB, and BHI with streptomycin and penicillin | 3 days, 37 °C/room temperature | [65] |
| Candida spp. | Clinical samples | SAB with chloramphenicol | 72 h, 28 °C | [66] |
| C. albicans, N. (Candida)glabrata, C. dubliniensis, P. (Candida)krusei, and M. (Candida) parapsilosi | Oral cavity | CAC | 3–4 days, 37 °C | [67] |
| C. albicans, C. tropicalis, P. (Candida)krusei, N. (Candida)glabrata, and C. dubliniensis | Clinical specimens | CAC | 24 h, 37 °C | [68] |
| C. africana | Vagina | CHROMagar and Candida ID2 | NI | [69] |
| Candida spp. | Denture-related stomatitis patients | SAB | 7 days, 35 °C | [70] |
| C. auris | Healthy human ear | CAC, Candida ID, SAB, and malt extract agar | 48 h, 37 °C | [71] |
| D. (Candida)famata | Eye infection | YEPD | 30 °C | [72] |
| C. albicans, N. (Candida)glabrata and C. kefyr (teleomorph Kluyveromyces marxianus) | Vulvovaginal candidiasis patients | SAB with chloramphenicol and cycloheximide | 48 h, 30 °C | [73] |
| N. (Candida)glabrata | Blood | CAC, SAB IMA, and MYC | 48 h/4–7 days, 30–37 °C | [74] |
| C. albicans, C. tropicalis and P. (Candida) krusei | Blood | CAC | 48 h, 37 °C | [75] |
| C. pararugosa (teleomorph Diutina pararugosa) | Oral cavity | SAB with chloramphenicol and CAC | 37/25 °C | [76] |
| Food Samples | ||||
| M. (Candida)parapsilosis | Contaminated dairy products | PDB | 24 h, 37 °C | [19] |
| C. albicans | Raw chicken meat and mutton meat | SAB | 7–10 days, 30 °C | [77] |
| Candida spp. | Mastitic bovine milk | Blood agar, Mac Conkey agar, and SAB | 72 h, 37 °C | [78] |
| C. albicans | Landes geese | CAC, YPD, TTC-Sha Baoluo, Corn meat agar medium | 72 h, 35 °C | [46] |
| C. albicans | Milk | SAB with chloramphenicol and CAC | 24 h, 37 °C | [79] |
| Candida spp. | Mastitic cows and milkers | SAB | 72 h, 37 °C | [80] |
| C. tropicalis | Infected sows | SAB and TSA with newborn bovine serum | 3–5 days, 28/37 °C, respectively | [81] |
| N. (Candida)glabrata | Avian species | SAB | 48 h, 37 °C | [82] |
| D. (Candida) rugosa | Turkeys | SAB | 2 weeks, 27 °C | [83] |
| Environmental Samples | ||||
| C. auris and N. (Candida) glabrata | Plastic health care surface | SAB; YNB | 40 °C | [54] |
| C. (Candida) utilis | Hulu Mur, Kissra, and Marisa (Sudanese traditionally cereal-based fermented foods) | YM with rose bengal | 3 days, 28 °C | [84] |
| C. auris | Sediment soil and seawater Coastal wetlands (e.g., rocky shores, sandy beaches, tidal marshes, and mangrove swamps) | SAB with chloramphenicol and gentamicin, CAC | 7 days, 28 °C | [85] |
| C. albicans | Fresh water and sewage | Ágar MacConkey | 24 h, 22 °C | [86] |
| C. albicans and Y. (Candida) lipolytica | Soil samples | PDA with chloramphenicol, CAC | 3–5 days, 28 ± 2 °C | [87] |
| C. digboiensis | Acid mine drainage | GPYE | 32 ± 2 °C | [88] |
| C. (Candida) membranifaciens subsp. flavinogenie strain W14-3 | Seawater and sediments | YPD with chloramphenicol | 5 days, 20–25 °C | [89] |
| Fermented Product | Starter Culture | Isolation Source | Desirable Traits/ Inoculation Effects | References |
|---|---|---|---|---|
| Wine | S. (Candida) zemplinina | Grapefruits and musts | Dominance observed during sequential inoculation with S. cerevisiae. Higher aroma complexity during co-inoculation (↑↑ production of aliphatic alcohols and free terpenes); | [1] |
| Organic wine must | Synergism when co-inoculated with S. cerevisiae. ↑↑ quantitative and qualitative content of esters | [119] | ||
| Spontaneous wine fermentation | ↓↓ concentration of residual sugar at the end of fermentation. ↑↑ glycerol production on trials containing the S. (Candida) zemplinina; Improvement of fruity and floral sensory notes when co-inoculated with S. cerevisiae and Lactiplantibacillus plantarum | [120] | ||
| C. railenensis | Vineyards | Decrease in ethyl and acetate esters production. Higher production of glycerol, acetate, free terpenes, and norisoprenoids | [2] | |
| W. (Candida) pulcherrina | Spontaneous wine fermentation | Slow rate of sugar consumption. ↑ production of ethyl acetate in comparison to pure S. cerevisae wine fermentation | [3] | |
| Cocoa | C. nitrativorans | Spontaneous cocoa beans fermentation | Natural adaptation to the cocoa fermentative process. Superior sugar metabolism. Killer phenotype trait. ↑↑ polygalacturonic and pectin lyase production | [4] |
| C. parapsilosis | Uniformity of well-fermented seeds (>95%); Lower acidity of the cotyledon in comparison to other inoculums. ↑↑ residual sugar at the end of fermentation. ↑ free amino acids content inside the cotyledon | [5] | ||
| Candida sp. | NI | Growth inhibition of pathogenic microorganisms. No significant alterations on cocoa quality parameters | [121] | |
| Coffee | M. (Candida) parapsilosis | Coffee fruits | ↑ proteolytic activity during semi-dry processing. Final beverage with prevalence of caramel- and fruity-like flavor, and dense body. Highest number of aroma-active compounds in semi-dry processing | [122,123,124] |
| Kefir | K. (Candida) kefyr | NI | ↑↑ proteolytic activity. Decrease on total fat content. ↑↑ residual sugar at the end of fermentation; | [125] |
| Bovine colostrum | Y. (Candida) lipolytica | NI | Increased antioxidant and antibacterial activity. Inhibition of Aspergillus niger sporulation | [126] |
| Cheese | K. (Candida) zeylanoides and H. (Candida). homilentoma | Spontaneous Italian Fossa cheese | Lower contamination by molds. ↑↑ sensorial attributes in comparison to spontaneous fermentation | [127] |
| Table olive | C. boidinii | Leucocarpa cv. olives brine | Alteration of color parameters. ↑↑ enzymatic activity, resulting in the texture loss of the fermented olives. ↓↓ acidification of the brine and presence of spoilage microorganisms at the end of the fermentation | [128] |
| Meat | C. deformans and K. (Candida) zeylanoides | Native dry-cured lacón | Lower production of flavor- and odor-active compounds associated to dry-cured meat | [129] |
| Beer | C. tropicalis | Traditional sub-Saharan sorghum beer | ↑↑ growth rate and cell concentration in comparison to S. cerevisiae. Lower glucose consumption rate. ↑ production of lactic acid in the final product. High viability rate in superior concentrations of ethanol | [6] |
| Soy sauce-like condiment | W. (Candida) versatilis | NI | Superior osmotic tolerance and growth rate at 10% NaCl soy whey medium. Superior acidification of the final product. ↑↑ residual sugar at the end of fermentation. ↑↑ contribution to umami taste Production of volatile compounds associated with desirable flavor in soy sauce | [130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Melo Pereira, G.V.; Maske, B.L.; de Carvalho Neto, D.P.; Karp, S.G.; De Dea Lindner, J.; Martin, J.G.P.; de Oliveira Hosken, B.; Soccol, C.R. What Is Candida Doing in My Food? A Review and Safety Alert on Its Use as Starter Cultures in Fermented Foods. Microorganisms 2022, 10, 1855. https://doi.org/10.3390/microorganisms10091855
de Melo Pereira GV, Maske BL, de Carvalho Neto DP, Karp SG, De Dea Lindner J, Martin JGP, de Oliveira Hosken B, Soccol CR. What Is Candida Doing in My Food? A Review and Safety Alert on Its Use as Starter Cultures in Fermented Foods. Microorganisms. 2022; 10(9):1855. https://doi.org/10.3390/microorganisms10091855
Chicago/Turabian Stylede Melo Pereira, Gilberto Vinícius, Bruna Leal Maske, Dão Pedro de Carvalho Neto, Susan Grace Karp, Juliano De Dea Lindner, José Guilherme Prado Martin, Bianca de Oliveira Hosken, and Carlos Ricardo Soccol. 2022. "What Is Candida Doing in My Food? A Review and Safety Alert on Its Use as Starter Cultures in Fermented Foods" Microorganisms 10, no. 9: 1855. https://doi.org/10.3390/microorganisms10091855