




A New Strain of Lecanicillium uredinophilum Isolated from Tibetan Plateau and Its Insecticidal Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens Collection and Isolation
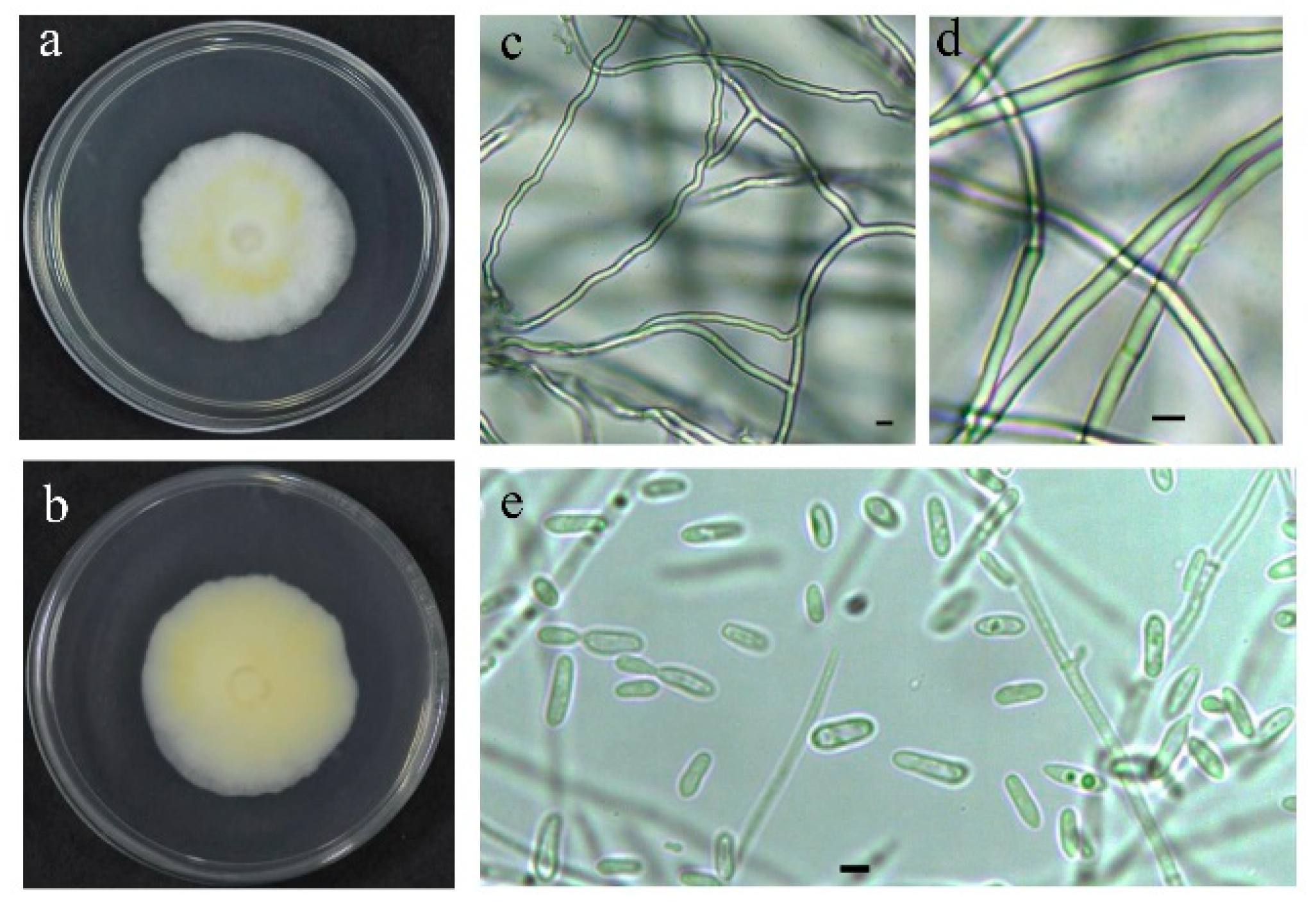
2.1.1. Morphological Observation of Strain
2.1.2. Extraction and Amplification of Genomic DNA
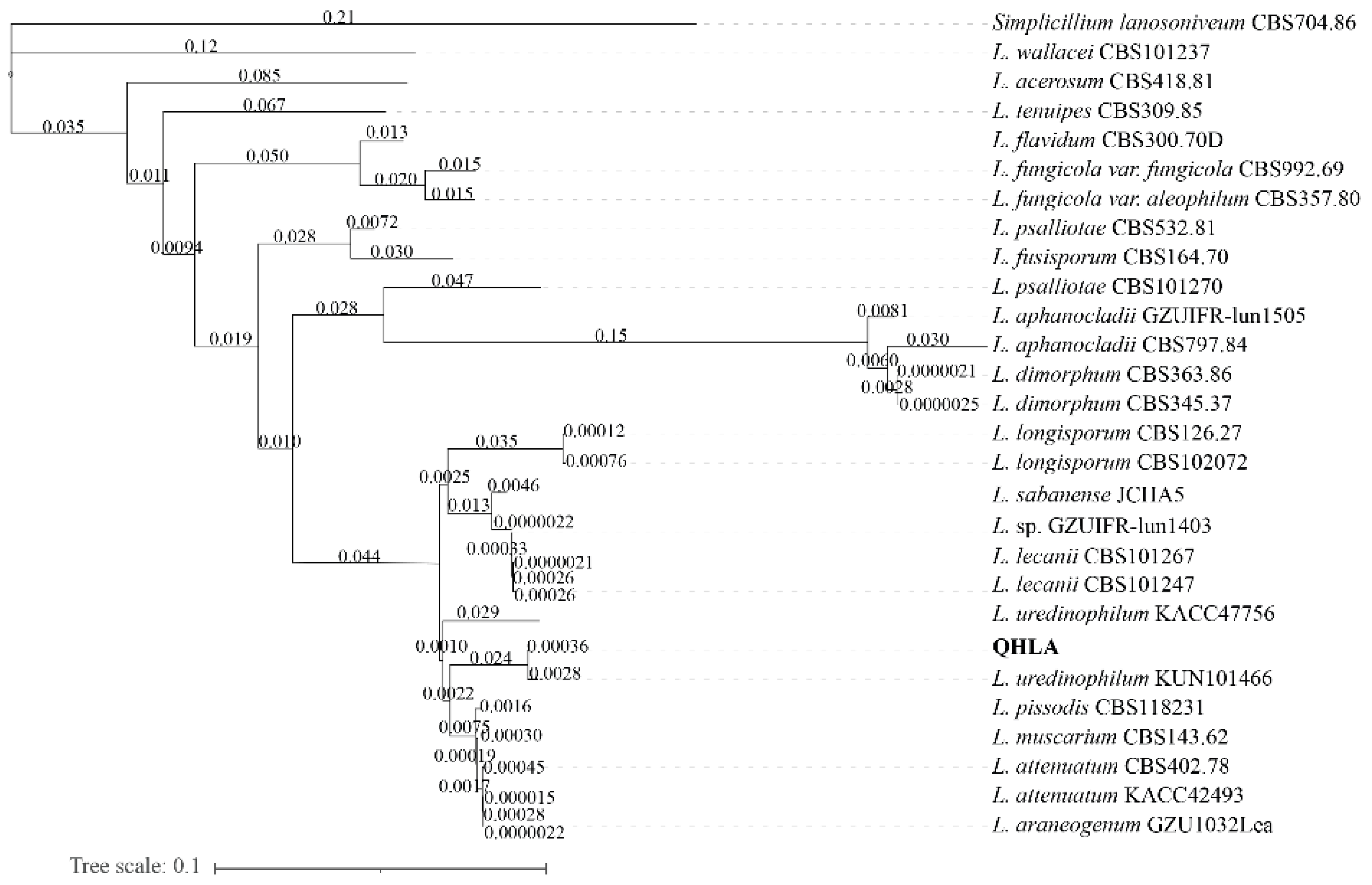
2.1.3. Phylogenetic Analysis
2.2. Biological Characteristics of the QHLA
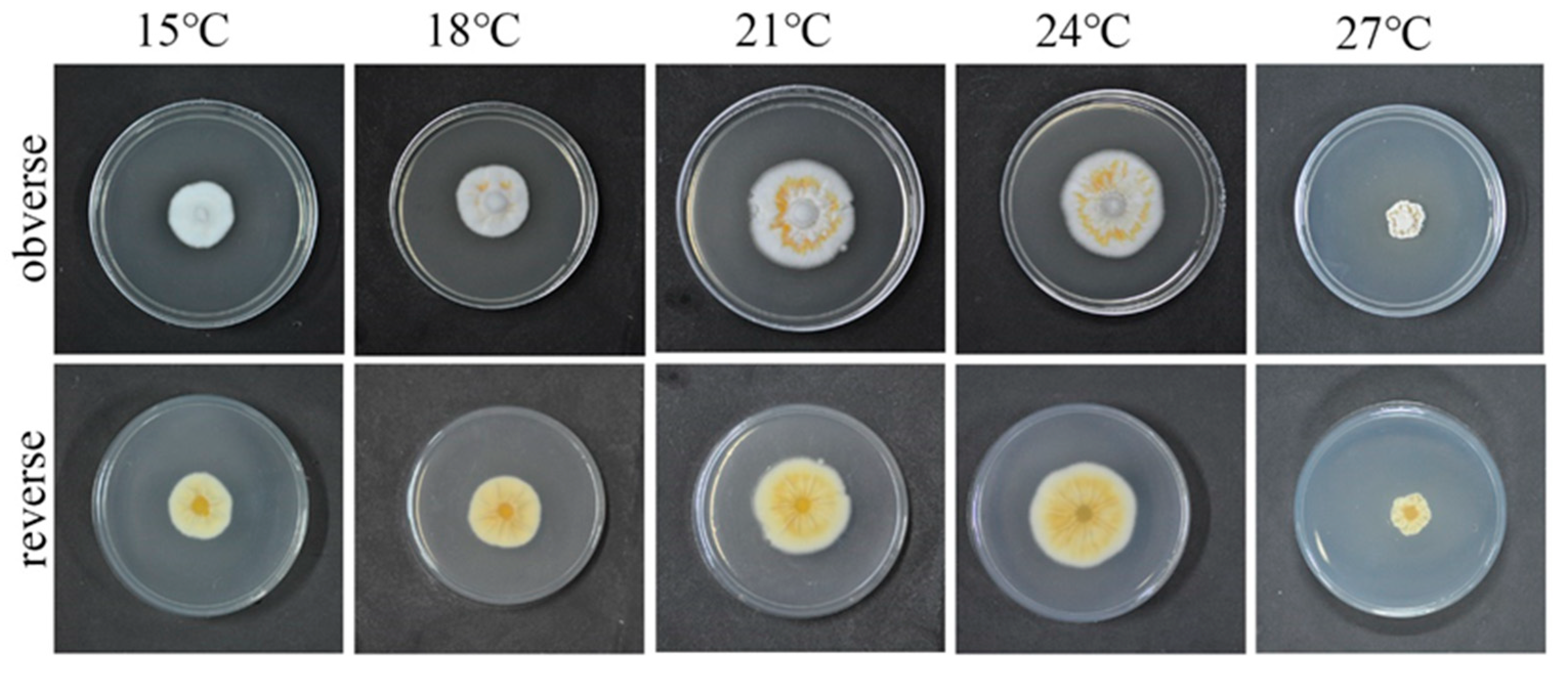
2.2.1. Determination of Optimized Culture Temperature for the Fungus
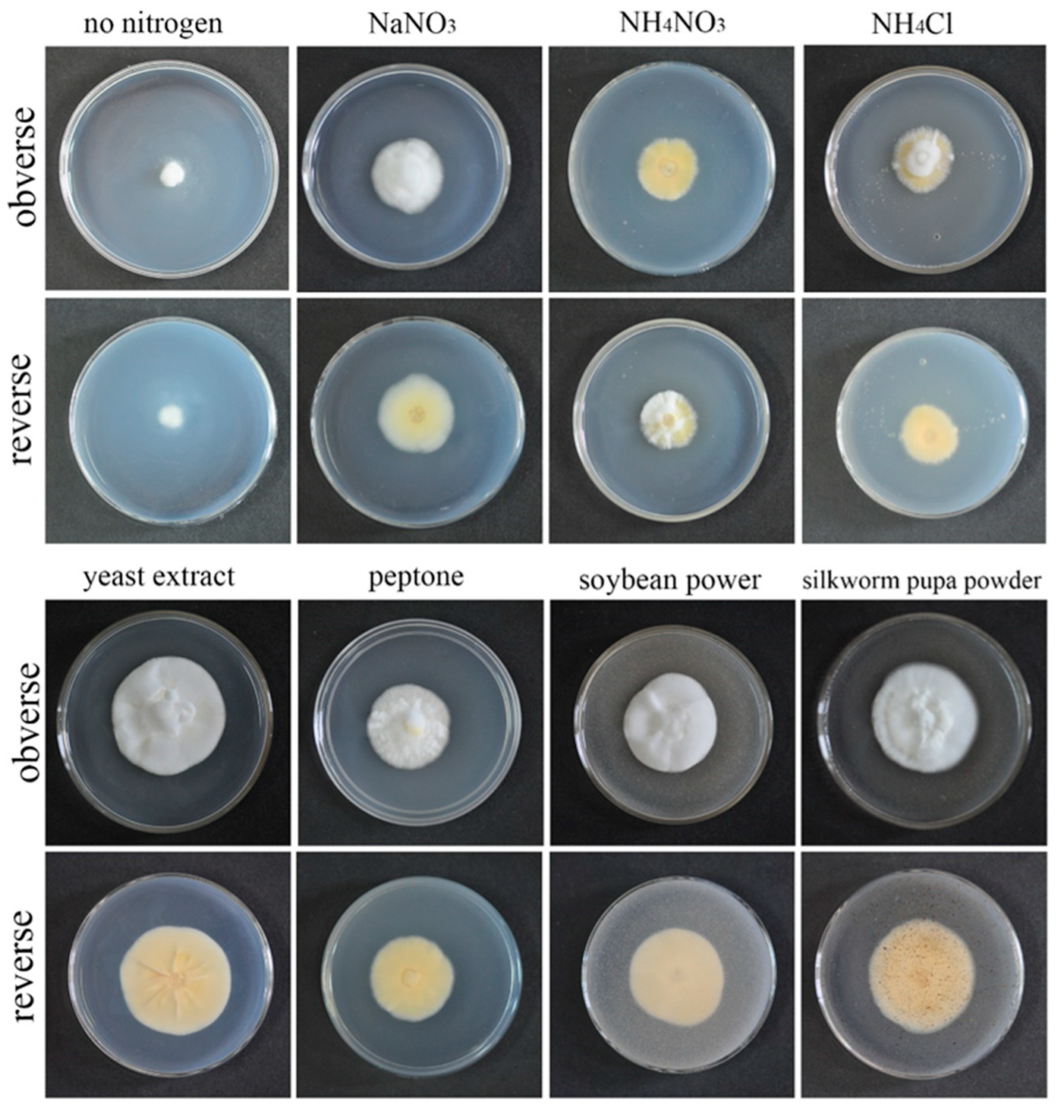
2.2.2. Determination of Optimized Nitrogen Sources for the Fungus
2.2.3. Determination of Optimized Carbon Sources for the Fungus
2.2.4. Determination of Optimized Liquid Medium for the Fungus
2.3. Bioassay for Insecticidal Activity
2.4. Statistical Analysis
3. Results
3.1. Isolation and Identification of Strain
3.2. Optimum Culture Temperature for the QHLA
3.3. Effects of Different Nitrogen Sources on Growth Rate and Sporulation Quantity of the Strain QHLA
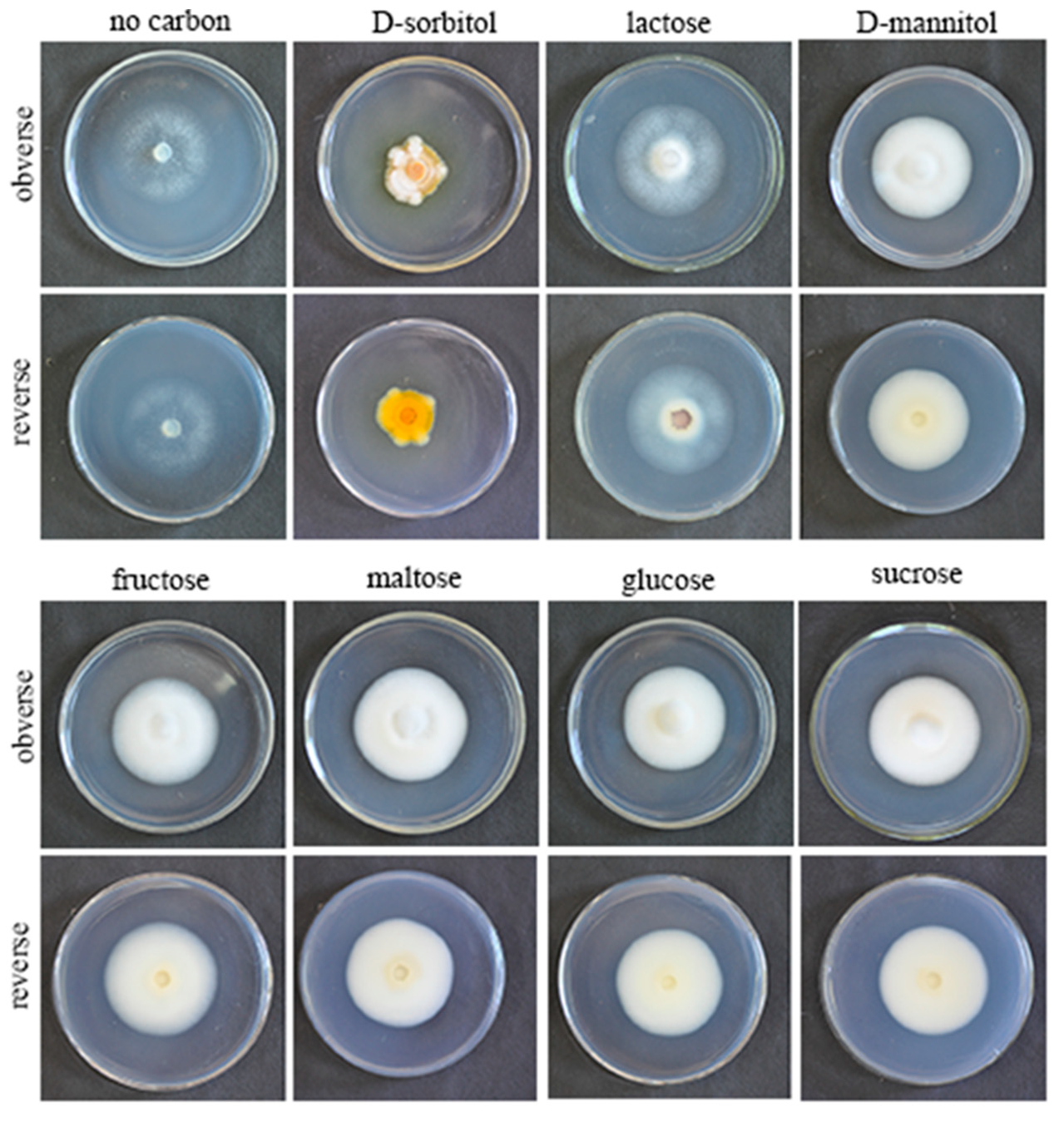
3.4. Effects of Different Carbon Sources on Growth Rate and Sporulation Quantity of the Strain QHLA
3.5. Optimum Liquid Medium for the QHLA
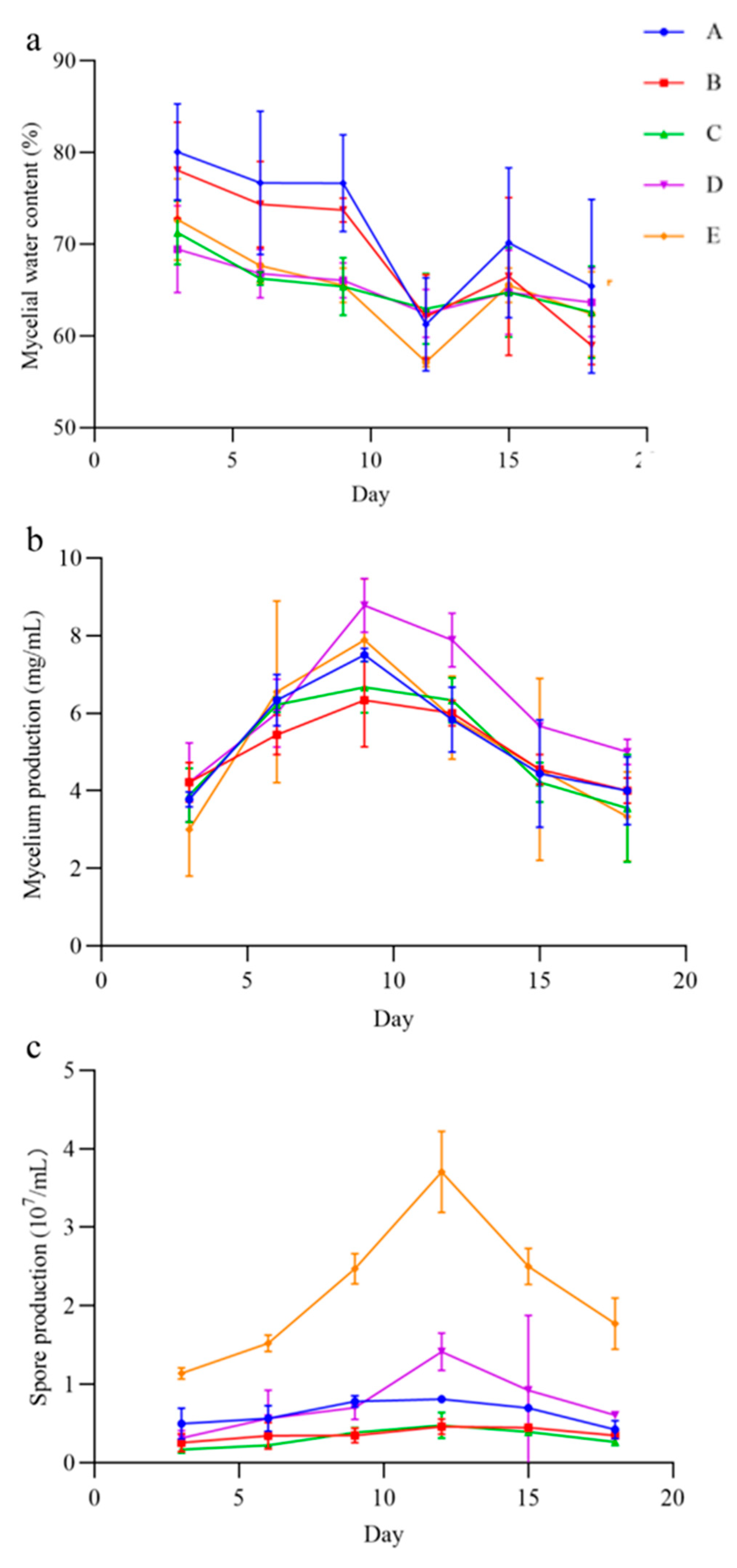
3.6. Bioassay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, S.K.; Maharachchikumbura, S.; Jeewon, R.; Bhat, D.J.; Al-Sadi, A.M. Lecanicillium subprimulinum (Cordycipitaceae, Hypocreales), a novel species from Baoshan, Yunnan. Phytotaxa 2018, 348, 99–108. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, Q.; Wang, D.; Zou, W.Q.; Tang, D.X.; Hongthong, P.; Yu, H. Species diversity and virulence potential of the Beauveria bassiana complex and Beauveria scarabaeidicola complex. Front. Microbiol. 2022, 13, 841604. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, X.; Tian, B.; Wang, M.; Niu, Q.; Zhang, K. Isolation and characterization of a serine protease from the nematophagous fungus, Lecanicillium psalliotae, displaying nematicidal activity. Biotechnol. Lett. 2005, 27, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Zhu, H.; Guo, Y.; Du, X.; Guo, J.; Zhang, L.; Qin, C. Lecanicillium coprophilum (Cordycipitaceae, Hypocreales), a new species of fungus from the feces of Marmota monax in China. Phytotaxa 2019, 387, 55–62. [Google Scholar] [CrossRef]
- Ponizovskaya, V.B.; GrumGrzhimaylo, A.A.; Georgieva, M.L.; Kokaeva, L.T.; Bilanenko, E.N. Lecanicillium gracile (Cordycipitaceae), a new species isolated from mineral building materials. Phytotaxa 2020, 443, 265–278. [Google Scholar] [CrossRef]
- Moyo, D.; Ishikura, S.; Rakotondrafara, A.; Clayton, M.; Kinoshita, R.; Tani, M.; Koike, M.; Aiuchi, D. Behavioral change of Bemisia tabaci and Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) infected by Lecanicillium muscarium (Hypocreales: Cordycipitaceae). Appl. Entomol. Zool. 2021, 56, 327–336. [Google Scholar] [CrossRef]
- Mitina, G.V.; Stepanycheva, E.A.; Choglokova, A.A.; Cherepanova, M.A. Features of behavioral reactions of the peach aphid Myzus persicae (Sulzer, 1776) (Hemiptera, Aphididae) to volatile organic compounds of entomopathogenic fungi of the genus Lecanicillium. Entomol. Rev. 2022, 101, 1015–1023. [Google Scholar] [CrossRef]
- Woo, R.M.; Park, M.G.; Choi, J.Y.; Park, D.H.; Kim, J.Y.; Wang, M.; Kim, H.J.; Woo, S.D.; Kim, J.S.; Je, Y.H. Insecticidal and insect growth regulatory activities of secondary metabolites from entomopathogenic fungi, Lecanicillium attenuatum. J. Appl. Entomol. 2020, 144, 655–663. [Google Scholar] [CrossRef]
- Nguyen, D.N.; Wang, S.L.; Nguyen, A.D.; Doan, M.D.; Tran, D.M.; Nguyen, T.H.; Ngo, V.A.; Doan, C.T.; Tran, T.N.; Do, V.C.; et al. Potential application of rhizobacteria isolated from the Central Highland of Vietnam as an effective biocontrol agent of Robusta Coffee nematodes and as a bio-fertilizer. Agronomy 2021, 11, 1887. [Google Scholar] [CrossRef]
- Gonalves, M.F.M.; Abreu, A.C.; Sandra, H.; Alves, A. Diversity of marine fungi associated with wood baits in the estuary Ria de Aveiro, with descriptions of Paralulworthia halima, comb. nov. Remispora submersa, sp. nov. and Zalerion pseudomaritima, sp. Nov. Mycologia 2021, 113, 664–683. [Google Scholar] [CrossRef]
- Abdulle, A.Y.; Nazir, T.; Keerio, A.U.; Ali, H.; Zaman, S.; Anwar, T.; Nam, T.D.; Qiu, D. In vitro virulence of three Lecanicillium lecanii strains against the whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt. J. Biol. Pest Control 2020, 30, 129. [Google Scholar] [CrossRef]
- Abdulle, A.Y.; Nazir, T.; Sayed, S.; Mahmoud, F.S.; Majeed, Z.M.; Aslam, U.M.H.; Iqbal, Z.; Nisar, S.M.; Keerio, U.A.; Ali, H.; et al. Sub-Lethal Effects of Lecanicillium lecanii (Zimmermann)-derived partially purified protein and its potential implication in Cotton (Gossypium hirsutum L.) defense against Bemisia tabaci Gennadius (Aleyrodidae: Hemiptera). Agriculture 2021, 11, 778. [Google Scholar] [CrossRef]
- Mochi, D.A.; Monteiro, A.C.; Simi, L.D.; Sampaio, M.A.A. Susceptibility of adult and larval stages of the horn fly, Haematobia irritans, to the entomopathogenic fungus Metarhizium anisopliae under field conditions. Vet. Parasitol. 2009, 166, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Sumalatha, B.V.; Selvaraj, K.; Poomesha, B.; Ramanujam, B. Pathogenicity of entomopathogenic fungus Isaria fumosorosea on rugose spiralling whitefly Aleurodicus rugioperculatus and its effect on parasitoid Encarsia guadeloupae. Biocontrol Sci. Technol. 2020, 30, 1150–1161. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Mathers, J.J.; Northing, P. The integrated use of chemical insecticides and the entomopathogenic nematode, Steinernema carpocapsae (Nematoda: Steinernematidae), for the control of sweetpotato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Sci. 2008, 15, 447–453. [Google Scholar] [CrossRef]
- Park, M.J.; Hong, S.B.; Shin, H.D. Lecanicillium uredinophilum sp. nov. associated with rust fungi from Korea. Mycotaxon 2015, 12, 997–1005. [Google Scholar] [CrossRef]
- Wei, D.P.; Wanasinghe, D.N.; Chaiwat, T.A.; Hyde, K.D. Lecanicillium uredinophilum known from rusts, also occurs on animal hosts with chitinous bodies. Asian J. Mycol. 2018, 1, 63–73. [Google Scholar] [CrossRef]
- He, W.; Zhao, X.; Ali, A.; Ge, S.; Zhang, H.; He, L.; Wu, K. Population dynamics and reproductive developmental analysis of Helicoverpa armigera (Lepidoptera: Noctuidae) trapped using food attractants in the field. J. Econ. Entomol. 2021, 114, 1533–1541. [Google Scholar] [CrossRef]
- Garlet, C.G.; Moreira, R.P.; Gubiani, P.D.S.; Palharini, R.B.; Farias, J.R.; Bernardi, O. Fitness cost of chlorpyrifos resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae) on different host plants. Environ. Entomol. 2021, 50, 898–908. [Google Scholar] [CrossRef]
- Wang, J.; Song, J.; Wu, X.; Deng, Q.; Zhu, Z.; Ren, M.; Ye, M.; Zeng, R. Seed priming with calcium chloride enhances wheat resistance against wheat aphid Schizaphis graminum Rondani. Pest Manag. Sci. 2021, 77, 4709–4718. [Google Scholar] [CrossRef]
- Rimantas, R.; Jekaterina, H.; Audrius, Z. Mitochondrial COI and morphological specificity of the mealy aphids (Hyalopterus ssp.) collected from different hosts in Europe (Hemiptera, Aphididae). ZooKeys 2013, 319, 255–267. [Google Scholar] [CrossRef]
- Li, S.; Lv, M.; Li, T.; Hao, M.; Xu, H. Spirodiclofen ether derivatives: Semisynthesis, structural elucidation, and pesticidal activities against Tetranychus cinnabarinus Boisduval, Aphis citricola Van der Goot and Mythimna separata Walker. Pest Manag. Sci. 2021, 77, 2395–2402. [Google Scholar] [CrossRef] [PubMed]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Sun, B.L. Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of ribosomal DNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 4599–4603. [Google Scholar] [CrossRef] [PubMed]
- Guhr, A.; Weig, A.R. Assessment of prokaryote to eukaryote ratios in environmental samples by SSU rDNA length polymorphism. Antonie Leeuwenhoek 2020, 113, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.H.; Hyten, A.S.; Spatafora, J.W. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef]
- Liu, Y.; Whelen, S.; Hall, B. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Zhou, Y.; Zou, X.; Zhi, J.; Xie, J.; Jiang, T. Fast recognition of Lecanicillium spp. and its virulence against Frankliniella occidentalis. Front. Microbiol. 2020, 11, 561381. [Google Scholar] [CrossRef]
- Zhang, D.F.; Gao, I.; Jakovlić, H.; Zou, J.; Zhang, W.; Li, X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, M.Y.; Liu, S.; Liu, M.K.; Ding, C.Y.; Qi, Y.Z.; Yan, X.L.; Qian, Y.B.; Xu, Y.F.; Yan, F.F. Optimization of culture conditions for a highly virulent Lecanicillium psalliotae and its pathogenicity against Geen Peach Aphid. Chin. J. Biol. Control 2021, 37, 349–355. [Google Scholar] [CrossRef]
- Russell, R.M.; Robertson, J.L.; Savin, N.E. POLO: A New Computer Program for Probit Analysis. Bull. Entomol. Soc. Am. 1977, 23, 209–213. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Wang, M.; Zhang, H.; Huang, Z.; Ma, J. Comparative study of the composition of cultivated, naturally grown Cordyceps sinensis, and stiff worms across different sampling years. PLoS ONE 2019, 14, e0225750. [Google Scholar] [CrossRef]
- Yang, X.; Chen, L.; Zhao, L.; Yang, Y.; Yan, L.; Tai, G.; Zhang, H. Cordyceps sinensis-derived fungus Isaria felina ameliorates experimental autoimmune thyroiditis in mice. Biomed. Pharmacother. 2021, 140, 111733. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Shi, J.D.; Wang, D. Three fungal strains isolated from stroma of Ophiocordyceps sinensis and their culture conditions. Mycosystema 2021, 40, 1991–2007. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Zhi, J.R.; Ye, M.; Zhang, Z.Y.; Yue, W.B.; Zou, X. Lecanicillium cauligalbarum sp. nov. (Cordycipitaceae, Hypocreales), a novel fungus isolated from a stemborer in the Yao Ren National Forest Mountain Park, Guizhou. Mycokeys 2018, 43, 59–74. [Google Scholar] [CrossRef]
- Mseddi, J.; FarhatTouzri, D.B.; Azzouz, H. Selection and characterization of thermotolerant Beauveria bassiana isolates and with insecticidal activity against the cotton-melon aphid Aphis gossypii (Glover) (Hemiptera: Aphididae). Pest Manag. Sci. 2022, 78, 2183–2195. [Google Scholar] [CrossRef]
- Kaifuchi, S.; Nonaka, K.; Mori, M.; Shiomi, K.; Satoshi, O.; Masuma, R. Lecanicillium primulinum, a new hyphomycete (Cordycipitaceae) from soils in the Okinawa’s main island and the Bonin Islands, Japan. Mycoscience 2013, 54, 291–296. [Google Scholar] [CrossRef]
- Omuse, E.R.; Niassy, S.; Wagacha, J.M.; Ong’amo, G.O.; Azrag, A.G.A.; Dubois, T. Suitable models to describe the effect of temperature on conidial germination and mycelial growth of Metarhizium anisopliae and Beauveria bassiana. Biocontrol Sci. Technol. 2022, 32, 281–298. [Google Scholar] [CrossRef]
- Tang, H.; Yoshizawa, S. Influences of temporal variations in environmental temperature and humidity on the fungus groethrate. Biocontrol Sci. 2000, 5, 51–55. [Google Scholar] [CrossRef]
- Zare, R.; Gams, W. A revision of the Verticillium fungicola species complex and its affinity with the genus Lecanicillium. Mycol. Res. 2008, 112, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Gao, L. Optimization of nutritional and environmental conditions for biomass production and sporulation of entomogenous fungus Lecanicillium lecanii CA-1-G using two-stage cultivation system. J. Yeast Fungal Res. 2018, 9, 14–20. [Google Scholar] [CrossRef]
- Wang, H.; Lei, Z.; Reitz, S.; Li, Y.; Xu, X. Production of microsclerotia of the fungal entomopathogen Lecanicillium lecanii (Hypocreales: Cordycipitaceae) as a biological control agent against soil-dwelling stages of Frankliniella occidentalis (Thysanoptera: Thripidae). Biocontrol Sci. Technol. 2013, 23, 234–238. [Google Scholar] [CrossRef]
- Gan, Z.; Yang, J.; Tao, N.; Liang, L.; Mi, Q.; Li, J.; Zhang, K.-Q. Cloning of the gene Lecanicillium psalliotae chitinase Lpchi1 and identification of its potential role in the biocontrol of root-knot nematode Meloidogyne incognita. Appl. Microbiol. Biotechnol. 2007, 76, 1309–1317. [Google Scholar] [CrossRef]
- Radwan, O.; Gunasekera, T.S.; Ruiz, O.N. Draft Genome Sequence of Lecanicillium sp. isolate LEC01, a fungus capable of hydrocarbon degradation. Microbiol. Resour. Announc. 2019, 8, e01744-18. [Google Scholar] [CrossRef]
- Montalva, C.; Valenzuela, E.; Barta, M.; Rojas, E.; Arismendi, N.; Rodrigues, J.; Humber, R.A. Lecanicillium attenuatum isolates affecting the invasive cypress aphid (Cinara cupressi) in Chile. Biocontrol 2017, 62, 625–637. [Google Scholar] [CrossRef]
- Kim, J.J.; Roberts, D.W. The relationship between conidial dose, moulting and insect developmental stage on the susceptibility of cotton aphid, Aphis gossypii, to conidia of Lecanicillium attenuatum, an entomopathogenic fungus. Biocontrol Sci. Technol. 2012, 22, 319–331. [Google Scholar] [CrossRef]
- Kim, J.J.; Goettel, M.S.; Gillespie, D.R. Potential of Lecanicillium species for dual microbial control of aphids and the cucumber powdery mildew fungus, Sphaerotheca fuliginea. Biol. Control 2007, 40, 327–332. [Google Scholar] [CrossRef]
- Askary, H.; Ajam, H.M.; Yarmand, H. Investigation on survival of Aphidius nigripes Ashmead (Hymenoptera: Aphidiidae) reared on infected potato aphid by Lecanicillium muscarium (Deut.: Moniliaceae). Commun. Agric. Appl. Biol. Sci. 2006, 71, 375–385. [Google Scholar]
- Biryol, S.; Araz, N.; Eski, A.; Aktürk, R.; Aksu, Y.; Çelik Göktürk, B.; Bilgin, L.; Demir, İ. Biodiversity and pathogenicity of entomopathogenic fungi associated with the lesser spruce sawfly, Pristiphora abietina. Entomol. Exp. Appl. 2021, 169, 414–423. [Google Scholar] [CrossRef]
- Juliya, R.F. Phylogeny, chitinase activity, and pathogenicity of Beauveria, Metarhizium and Lecanicillium species against cowpea aphid, Aphis craccivora Koch. Int. J. Trop. Insect Sci. 2019, 40, 309–314. [Google Scholar] [CrossRef]
- Keerio, A.U.; Nazir, T.; Abdulle, Y.A.; Jatoi, G.H.; Gadhi, M.A.; Anwar, T.; Sokea, T.; Qiu, D. In vitro pathogenicity of the fungi Beauveria bassiana and Lecanicillium lecanii at different temperatures against the whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt. J. Biol. Pest Control 2020, 30, 41. [Google Scholar] [CrossRef]
- Broumandnia, F.; Rajabpour, A. Efficacies of some isolates of Lecanicillium lecanii to control Tribolium castaneum (Col. Tenebrionidae). J. Plant Dis. Prot. 2020, 127, 625–631. [Google Scholar] [CrossRef]
- Aiuchi, D.; Moyo, D.; Ishikura, S.; Tani, M.; Kinoshita, R.; Rakotondrafara, A.M.; Koike, M. Virulence of Lecanicillium spp. (Hypocreales: Cordycipitaceae) hybrid strains against various biological stages of the greenhouse whitefly, Trialeurodes vaporariorum (Hemiptera: Aleyroidae). Biocontrol Sci. Technol. 2020, 30, 1006–1017. [Google Scholar] [CrossRef]
- Ben Fekih, I.; Jensen, A.B.; Boukhris-Bouhachem, S.; Pozsgai, G.; Rezgui, S.; Rensing, C.; Eilenberg, J. Virulence of two entomophthoralean fungi, Pandora neoaphidis and Entomophthora planchoniana, to Their Conspecific (Sitobion avenae) and Heterospecific (Rhopalosiphum padi) Aphid Hosts. Insects 2019, 10, 54. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen Source | Colony Growth Rate (mm/d) | Spore Production (spore/mL) | Colonial Morphology | Mycelial Growth Vigor |
|---|---|---|---|---|
| Yeast extract | 4.63 ± 0.03 a | 1.15 × 106 d | The obverse side of the colony was white, and the reverse side was yellow with a white edge. | +++ |
| Silkworm pupa powder | 4.61 ± 0.19 a | 1.85 × 106 cd | The obverse side of the colony was white, and the reverse side was pale yellow with a white edge. | +++ |
| Soybean power | 4.03 ± 0.20 b | 2.40 × 106 bc | The obverse side of the colony was white, and the reverse side was pale yellow with a white edge. | +++ |
| Peptone | 3.33 ± 0.56 c | 2.02 × 106 cd | The obverse side of the colony was white, and the reverse side was pale yellow with a white edge. | ++ |
| NaNO3 | 3.00 ± 0.06 d | 1.40 × 106 cd | The obverse side of the colony was white, and the reverse side was pale yellow with a white edge. | ++ |
| NH4NO3 | 2.24 ± 0.06 e | 2.74 × 106 b | Both sides of the colony were golden yellow with white aerial mycelia. | + |
| NH4Cl | 2.22 ± 0.20 e | 4.08 × 106 a | The obverse of the colony was white in the middle with white aerial mycelia and yellow around it. The reverse side was yellow. | + |
| − | 0.52 ± 0.18 f | − | The colony was no obvious mycelium on both sides | + |
| Carbon Source | Colony Growth Rate (mm/d) | Spore Production (spore/mL) | Colonial Morphology | Mycelial Growth Vigor |
|---|---|---|---|---|
| Maltose | 2.65 ± 0.08 a | 7.17 × 105 b | The obverse of the colony was white. The middle of the reverse was light yellow and the edge was white. | +++ |
| Fructose | 2.64 ± 0.06 a | 7.17 × 105 b | The obverse of the colony was white. The middle of the reverse was milky colored with white edges. | ++ |
| D-mannitol | 2.56 ± 0.25 ab | 5.33 × 105 b | The middle of the obverse of the colony was yellow and the edge was white. The middle of the reverse was white, and the edge was pale yellow with white aerial mycelia. | ++ |
| Lactose | 2.51 ± 0.11 ab | 9.33 × 105 b | The obverse of the colony was white, and there were dense aerial mycelia around the disk. The reverse was brown in the middle and white around the edges. | + |
| Glucose | 2.45 ± 0.24 ab | 7.59 × 105 b | The obverse of the colony was white, and the middle of the reverse was milky colored with white edges. | +++ |
| Sucrose | 2.35 ± 0.11 b | 4.83 × 105 b | The obverse of the colony was white, and the middle of the reverse was light yellow with white edges. | +++ |
| D-sorbitol | 2.01 ± 0.10 c | 1.63 × 106 a | The obverse of the colony was orange, with white aerial mycelia above. The reverse was orange. | + |
| − | 0.31 ± 0.13 d | − | Colonies were white on both sides. | + |
| Pests | LC50 (spore/mL) | 95% Confidence Interval | LC90 (spore/mL) | 95% Confidence Interval |
|---|---|---|---|---|
| Henosepilachna vigintioctopunctata | 4.83 × 105 | 7.811–3169.280 | 7.12 × 1010 | 3.51–∞ |
| Spodoptera exigua | 1.32 × 105 | 2.001–970.003 | 1.39 × 109 | 1.492–∞ |
| Plutella xylostella | 4.83 × 105 | 7.811–3169.280 | 7.12 × 1010 | 3.51–∞ |
| Spodoptera frugiperda | 4.83 × 105 | 7.811–3169.280 | 7.12 × 1010 | 3.51–∞ |
| Sitobion avenae | 6.30 × 103 | 0.215–1.713 | 2.36 × 108 | 0.187–247.71 |
| Hyalopterus perikonus | 1.49 × 104 | 0.826–2.260 | 1.88 × 109 | 12.215–34.82 |
| Aphis citricola | 6.40 × 103 | 0.042–1.923 | 2.11 × 107 | 0.0340–28.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, Y.; Wellabada Hewage Don, P.I.D.; Wang, D. A New Strain of Lecanicillium uredinophilum Isolated from Tibetan Plateau and Its Insecticidal Activity. Microorganisms 2022, 10, 1832. https://doi.org/10.3390/microorganisms10091832
Meng Y, Wellabada Hewage Don PID, Wang D. A New Strain of Lecanicillium uredinophilum Isolated from Tibetan Plateau and Its Insecticidal Activity. Microorganisms. 2022; 10(9):1832. https://doi.org/10.3390/microorganisms10091832
Chicago/Turabian StyleMeng, Yun, P. I. Damithri Wellabada Hewage Don, and Dun Wang. 2022. "A New Strain of Lecanicillium uredinophilum Isolated from Tibetan Plateau and Its Insecticidal Activity" Microorganisms 10, no. 9: 1832. https://doi.org/10.3390/microorganisms10091832