
Laboratory Risk Assessment of Three Entomopathogenic Fungi Used for Pest Control toward Social Bee Pollinators
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Stingless Bees
2.2. Honey Bees and Bumble Bees
2.3. Fungal Material
2.4. Fungal Exposure Bioassay
2.5. Statistical Analysis
3. Results
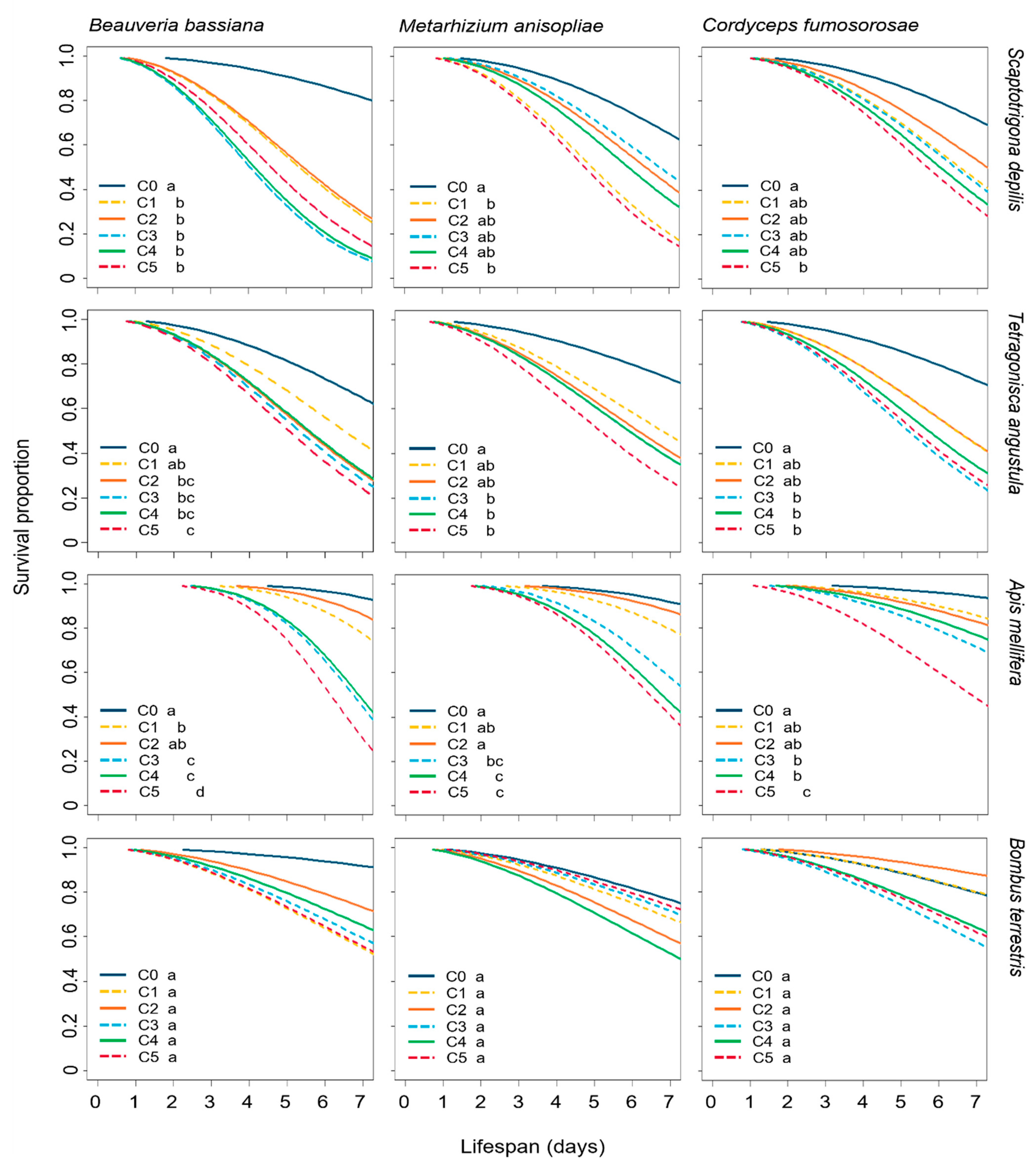
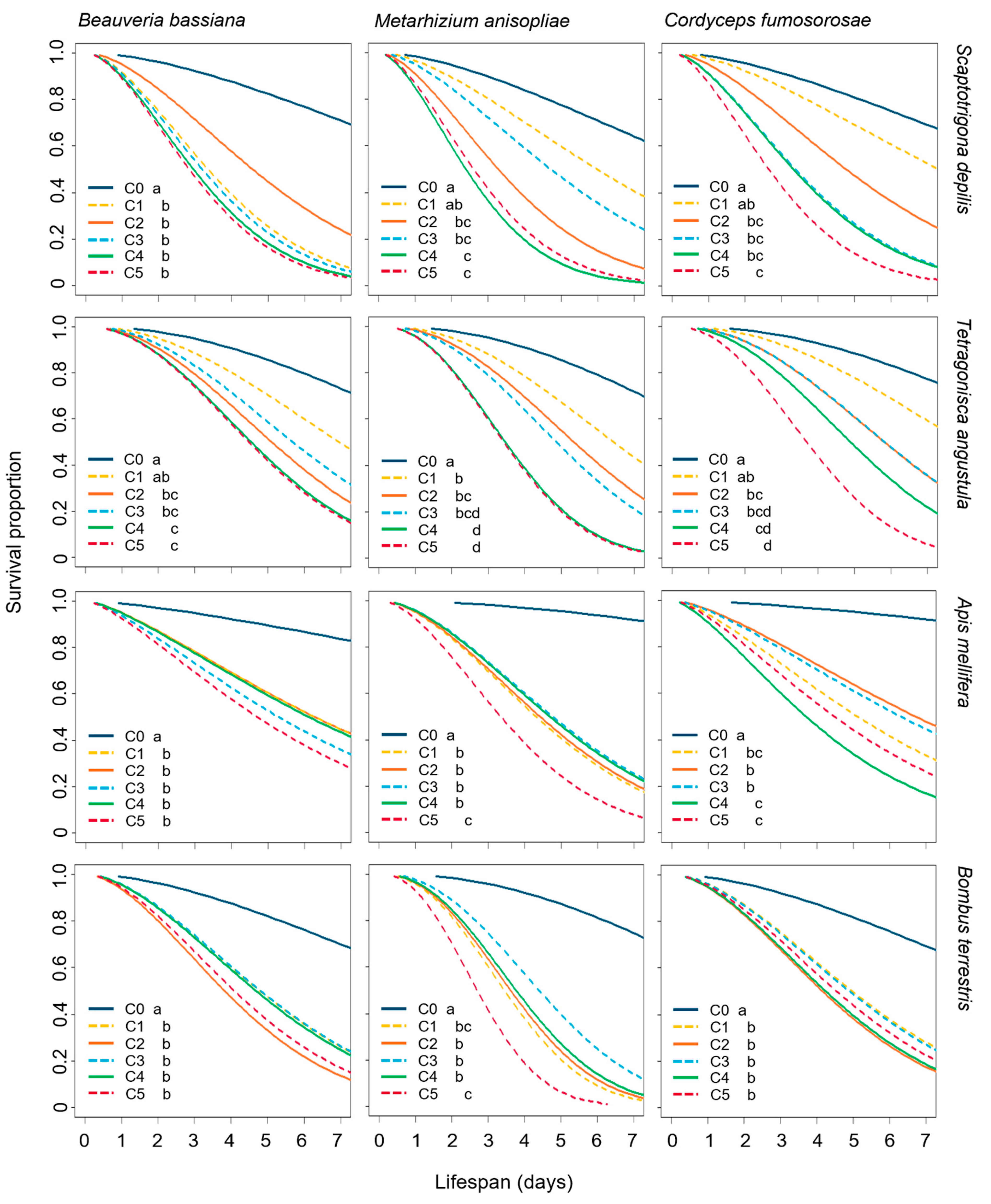
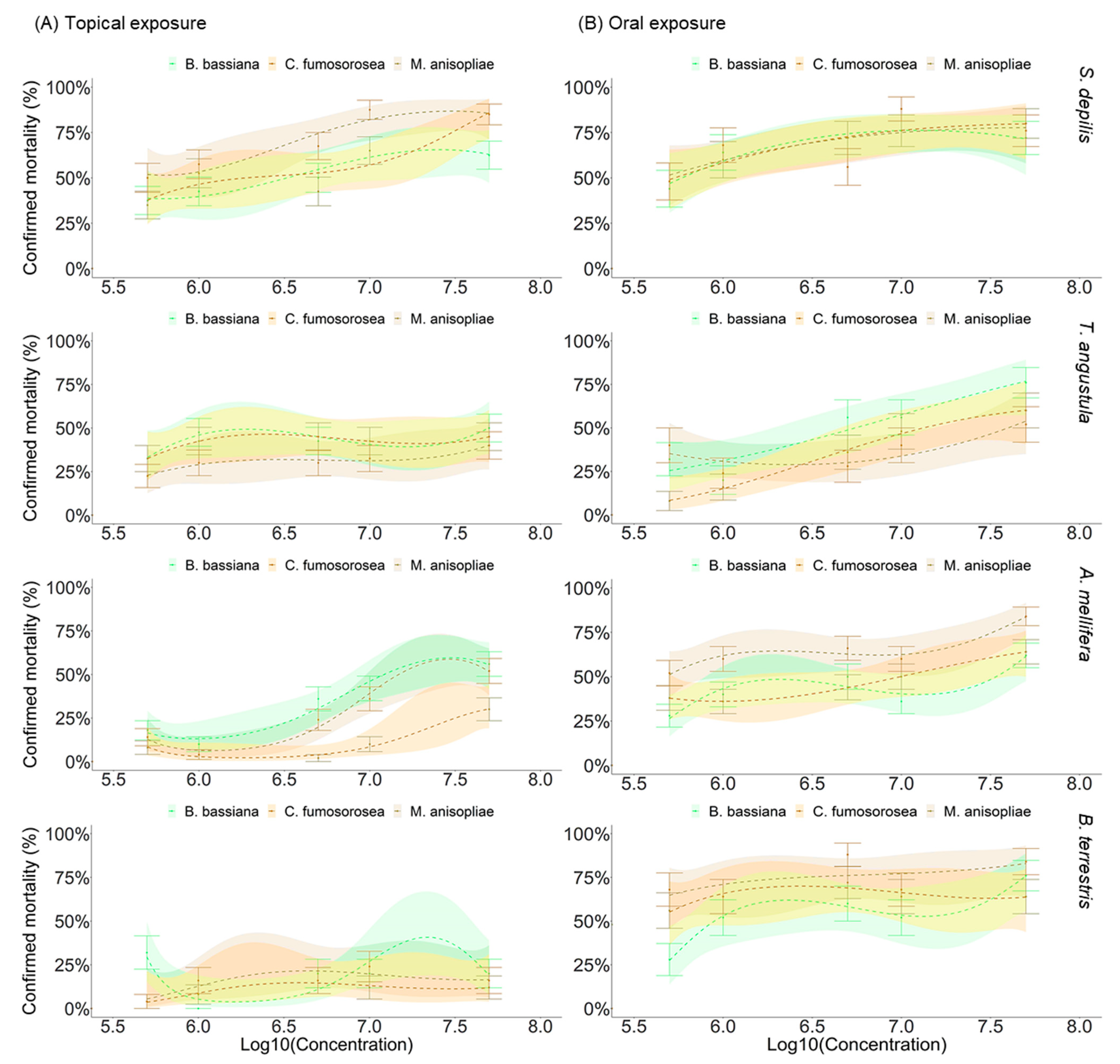
3.1. Effect of EF on the Survival of Bees
3.2. Sporulation of Entomopathgenic Fungi on Dead Bees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaronski, S.T.; Mascarin, G.M. Mass production of fungal entomopathogens. In Microbial Control of Insects and Mite Pests, 1st ed.; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 141–155. [Google Scholar]
- Van Lenteren, J.C.; Bolckmans, K.; Kohl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Arthurs, S.; Dara, S.K. Microbial biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2018, 165, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Mascarin, G.M.; Lopes, G.B.; Jr, I.D.; Fernandes, E.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef]
- Li, Z.; Alves, S.B.; Roberts, D.W.; Fan, M.; Jr, I.D.; Tang, J.; Lopes, R.B.; Faria, M.; Rangel, D.E. Biological control of insects in Brazil and China: History, current programs and reasons for their successes using entomopathogenic fungi. Bioc. Sci. Techn. 2010, 2, 117–136. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Wraight, S.P.; Galaini-Wraight, S.; Howes, R.L.; Castrillo, L.A.; Griggs, M.H.; Carruthers, R.I.; Smith, R.H.; Matsumoto, T.K.; Keith, L.M. Efficacy of Beauveria bassiana strain GHA spray applications against coffee berry borer Hypothenemus hampei on Hawai‘i Island. Biol. Control 2021, 161, 104587. [Google Scholar] [CrossRef]
- Ausique, J.J.S.; D’Alessandro, C.P.; Conceschi, M.R.; Mascarin, G.M.; Delalibera Jr, I. Efficacy of entomopathogenic fungi against adult Diaphorina citri from laboratory to field applications. J. Pest Sci. 2017, 90, 947–960. [Google Scholar] [CrossRef]
- Castro, T.; Mayerhofer, J.; Enkerli, J.; Eilenberg, J.; Meyling, N.V.; de Andrade Moral, R.; Demetrio, D.G.B.; Jr, I.D. Persistence of Brazilian isolates of the entomopathogenic fungi Metarhizium anisopliae and M. robertsii in strawberry crop soil after soil drench application. Agric. Ecosyst. Environ. 2016, 233, 361–369. [Google Scholar] [CrossRef]
- Kapongo, J.P.; Shipp, L.; Kevan, P.; Sutton, J.C. Co-vectoring of Beauveria bassiana and Clonostachys rosea by bumble bees (Bombus impatiens) for control of insect pests and suppression of grey mould in greenhouse tomato and sweet pepper. Biol. Control 2008, 46, 508–514. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Egan, P.A.; Dicks, L.V.; Hokkanen, H.M.; Stenberg, J.A. Delivering integrated pest and pollinator management (IPPM). Trends Plant Sci. 2020, 25, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Salinas, A.; Chain-Guadarrama, A.; Aristizábal, N.; Vilchez-Mendoza, S.; Cerda, R.; Ricketts, T.H. Interacting pest control and pollination services in coffee systems. Proc. Natl. Acad. Sci. USA 2022, 119, e2119959119. [Google Scholar] [CrossRef] [PubMed]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microb. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef]
- Vestergaard, S.; Cherry, A.; Keller, S.; Goettel, M. Safety of hyphomycete fungi as microbial control agents. In Environmental Impacts of Microbial Insecticides, 1st ed.; Hokkanen, H.M.T., Hajek, A.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 35–62. [Google Scholar]
- Garrido-Jurado, I.; Ruano, F.; Campos, M.; Quesada-Moraga, E. Effects of soil treatments with entomopathogenic fungi on soil dwelling non-target arthropods at a commercial olive orchard. Biol. Control 2011, 59, 239–244. [Google Scholar] [CrossRef]
- Shaw, D.E. The incidental collection of fungal spores by bees and the collection of spores in lieu of pollen. Bee World 1990, 71, 158–176. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.L.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef]
- Osterman, J.; Aizen, M.A.; Biesmeijer, J.C.; Bosch, J.; Howlett, B.G.; Inouye, D.W.; Jung, C.; Martins, D.J.; Medel, R.; Pauw, A.; et al. Global trends in the number and diversity of managed pollinator species. Agric. Ecosyst. Environ. 2021, 322, 107653. [Google Scholar] [CrossRef]
- Klatt, B.K.; Holzschuh, A.; Westphal, C.; Clough, Y.; Smit, I.; Pawelzik, E.; Tscharntke, T. Bee pollination improves crop quality, shelf life and commercial value. Proc. R. Soc. B 2014, 281, 20132440. [Google Scholar] [CrossRef] [PubMed]
- Nowell, D.; Maynard, G.V. International guidelines for the export, shipment, import, and release of biological control agents and other beneficial organisms (international standard for phytosanitary measures no. 3). In Proceedings of the Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12–16 September 2005. [Google Scholar]
- Mazra’awi, M.S.A.; Shipp, J.L.; Broadbent, A.B.; Kevan, P.G. Dissemination of Beauveria bassiana by honey bees (Hymenoptera: Apidae) for control of tarnished plant bug (Hemiptera: Miridae) on canola. Environ. Entomol. 2006, 35, 1569–1577. [Google Scholar] [CrossRef]
- Butt, T.M.; Carreck, N.L.; Ibrahim, L.; Williams, I.H. Honey bee mediated infection of pollen beetle (Meligethes spp.) by the insect-pathogenic fungus, Metarhizium Anisopliae. Biocontrol Sci. Technol. 2010, 8, 533–538. [Google Scholar] [CrossRef]
- Carlesso, D.; Smargiassi, S.; Sassoli, L.; Cappa, F.; Cervo, R.; Baracchi, D. Exposure to a biopesticide interferes with sucrose responsiveness and learning in honey bees. Sci. Rep. 2020, 10, 19929. [Google Scholar] [CrossRef] [PubMed]
- Colombo, F.C.; Maciel, R.M.A.; Abati, R.; Raulino-Domanski, F.; Longhi, S.J.; Costa-Maia, F.M.; Vismara, E.S.; Lozano, E.R.; Potrich, M. Do Beauveria bassiana and Metarhizium anisopliae affect worker survival and the production of Africanized Apis mellifera queens? J. Apic. Res. 2021, 60, 260–269. [Google Scholar] [CrossRef]
- Peng, G.; Tong, S.; Zeng, D.; Xiaa, Y.; Feng, M. Colony heating protects honey bee populations from a risk of contact with wide-spectrum Beauveria bassiana insecticides applied in the field. Pest Manag. Sci. 2020, 76, 2627–2634. [Google Scholar] [CrossRef]
- Sinia, A.; Guzman-Novoa, E. Evaluation of the entomopathogenic fungi Beauveria bassiana GHA and Metarhizium anisopliae UAMH 9198 alone or in combination with thymol for the control of Varroa destructor in honey bee (Apis mellifera) colonies. J. Apic. Res. 2018, 57, 308–316. [Google Scholar] [CrossRef]
- Shipp, J.L.; Kapongo, J.P.; Park, H.; Kevan, P. Effect of bee-vectored Beauveria bassiana on greenhouse beneficials under greenhouse cage conditions. Biol. Control 2012, 63, 135–142. [Google Scholar] [CrossRef]
- Toledo-Hernandez, R.A.; Ruiz-Toledo, J.; Toledo, J.; Sanchez, D. Effect of three entomopathogenic fungi on three species of stingless bees (Hymenoptera: Apidae) under laboratory conditions. J. Econ. Entomol. 2016, 109, 1015–1019. [Google Scholar] [CrossRef]
- Conceição, P.d.J.; Neves, C.M.L.; Sodré, G.S.; Carvalho, C.A.L.; Souza, A.V.; Ribeiro, G.S.; Pereira, R.C. Susceptibility of Melipona scutellaris Latreille, 1811 (Hymenoptera: Apidae) to Beauveria bassiana (Bals.) Vuill. Sociobiology 2014, 61, 184–188. [Google Scholar] [CrossRef]
- James, R.R.; McGuire, M.R.; Leland, J.E. Susceptibility of adult alfalfa leafcutting bees1 and honey bees2 to a microbial control agent, Beauveria bassiana. Southwest. Entomol. 2012, 37, 13–21. [Google Scholar] [CrossRef]
- Erler, S.; Eckert, J.H.; Steinert, M.; Alkassab, A.T. Impact of microorganisms and entomopathogenic nematodes used for plant protection on solitary and social bee pollinators: Host range, specificity, pathogenicity, toxicity, and effects of experimental parameters. Environ. Pollut. 2022, 302, 119051. [Google Scholar] [CrossRef]
- Cappa, F.; Baracchi, D.; Cervo, R. Biopesticides and insect pollinators: Detrimental effects, outdated guidelines, and future directions. Sci. Total Environ. 2022, 837, 155714. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.R.O.; Soares, H.M.; Carvalho, S.M.; Nocelli, R.C.F.; Malaspina, O. Acute toxicity of fipronil to the stingless bee Scaptotrigona postica Latreille. Bul. Environ. Contam. Toxic. 2013, 90, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Del Sarto, M.C.L.; Oliveira, E.E.; Guedes, R.N.C.; Campos, L.A.O. Differential insecticide susceptibility of the Neotropical stingless bee Melipona quadrifasciata and the honey bee Apis mellifera. Apidologie 2014, 45, 626–636. [Google Scholar] [CrossRef]
- Alves, S.B. Entomopathogenic fungi. In Controle Microbiano de Insetos, 2nd ed.; Alves, S.B., Ed.; Fundação de Estudos Agrários Luiz de Queiroz (FEALQ): Piracicaba, Brazil, 1998; pp. 289–370. [Google Scholar]
- Kassambara, A.; Kosinski, M.; Biecek, P.; Fabian, S. survminer: R Package Version 0.4, 8, 556, Drawing Survival Curves using “ggplot2.”, 2020. Available online: https://CRAN.R-project.org/package=survminer (accessed on 21 April 2021).
- Therneau, T. R package version 3.2-3, A Package for Survival Analysis in R., [Computer software], Mayo Clinic, Rochester, MN, USA, 2020. Available online: https://CRAN.R-project.Org/package=survival (accessed on 21 April 2021).
- Takakura, K.I. Bayesian estimation for the effectiveness of pesticides and repellents. J. Econ. Entomol. 2012, 105, 1856–1862. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S.; Hothorn, M.T. Package ‘Multcomp’, Simultaneous Inference in General Parametric Models; Project for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-normal plots and overdispersed models in R: The hnp package. J. Stat. Softw. 2017, 81, 1–23. [Google Scholar] [CrossRef]
- Omuse, E.R.; Niassy, S.; Kiatoko, N.; Lattorff, H.M.G.; Wagacha, J.M.; Dubois, T. A fungal-based pesticide does not harm pollination service provided by the African stingless bee Meliponula ferruginea on cucumber (Cucumis sativus). Apidologie 2022, 53, 28. [Google Scholar] [CrossRef]
- Potrich, M.; Silva, R.T.L.; Maia, F.M.C.; Lozano, E.R.; Rossi, R.M.; Colombo, F.C.; Tedesco, F.G.; Gouvea, A. Effect of entomopathogens on Africanized Apis mellifera L. (Hymenoptera: Apidae). Revta. Bras. Entomol. 2018, 62, 23–28. [Google Scholar] [CrossRef]
- Espinosa-Ortiz, G.E.; Lara-Reyna, J.; Otero-Colina, G.; Alatorre-Rosas, R.; Valdez-Carrasco, J. Susceptibility of larval, pupal and adult honey bees to isolates of Beauveria bassiana (Bals.) Vuill., Metarhizium anisopliae (Sorokin) and Paecilomyces fumosoroseus (Wize). Interciencia 2011, 36, 148–152. [Google Scholar]
- Demirozer, O.; Uzun, A.; Gosterit, A. Lethal and sublethal effects of different biopesticides on Bombus terrestris (Hymenoptera: Apidae). Apidologie 2022, 53, 24. [Google Scholar] [CrossRef]
- Hokkanen, H.M.T.; Zeng, Q.Q.; Menzler-Hokkanen, I. Assessing the impacts of Metarhizium and Beauveria on bumble bees. In Environmental Impacts of Microbial Insecticides, 1st ed.; Hokkanen, H.M.T., Hajek, A.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 63–72. [Google Scholar]
- Karise, R.; Muljar, R.; Smagghe, G.; Kaart, T.; Kuusik, A.; Dreyersdorff, G.; Williams, I.H.; Mand, M. Sublethal effects of kaolin and the biopesticides Prestop-Mix and BotaniGard on metabolic rate, water loss and longevity in bumble bees (Bombus terrestris). J. Pest Sci. 2016, 89, 171–178. [Google Scholar] [CrossRef]
- Mommaerts, V.; Sterk, G.; Hoffmann, L.; Smagghe, G. A laboratory evaluation to determine the compatibility of microbiological control agents with the pollinator Bombus terrestris. Pest Manag. Sci. 2009, 65, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Dunlap, C.A.; Jaronski, S.T. Ecological considerations in producing and formulating fungal entomopathogens for use in insect biocontrol. BioControl 2010, 55, 129–145. [Google Scholar] [CrossRef]
- Boucias, D.G.; Pendland, J.C.; Latge, J.P. Nonspecific factors involved in attachment of entomopathogenic Deuteromycetes to host insect cuticle. Appl. Environ. Microbiol. 1988, 54, 1795–1805. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; The John Hopkins University Press: Baltimore, MD, USA, 2000; p. 972. [Google Scholar]
- Vaknin, Y.; Gan-Mor, S.; Bechar, A.; Ronen, B.; Eisikowitch, D. The role of electrostatic forces in pollination. Pollen Pollinat. 2000, 222, 133–142. [Google Scholar]
- Maistrou, S.; Natsopoulou, M.E.; Jensen, A.B.; Meyling, N.V. Virulence traits within a community of the fungal entomopathogen Beauveria: Associations with abundance and distribution. Fungal Ecol. 2020, 48, 100992. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef]
- Roberts, D.W.; Leger, R.J.S. Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 2004, 54, 1–70. [Google Scholar]
- Wang, C.; Leger, R.J.S. The MAD1 adhesin of Metarhizium anisopliae links adhesion with blastospore production and virulence to insects, and the MAD2 adhesin enables attachment to plants. Eukaryot. Cell 2007, 6, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Leemon, D.M.; Jonsson, N.N. Comparative studies on the invasion of cattle ticks (Rhipicephalus (Boophilus) microplus) and sheep blowflies (Lucilia cuprina) by Metarhizium anisopliae (Sorokin). J. Invertebr. Pathol. 2012, 109, 248–259. [Google Scholar] [CrossRef]
- Schabel, S. Oral infection of Hylobius pales by Metarhizium anisopliae. J. Invertebr. Pathol. 1976, 383, 377–383. [Google Scholar] [CrossRef]
- Dillon, R.J.; Charnley, A.K. Invasion of the pathogenic fungus Metarhizium anisopliae through the guts of germfree desert locusts. Schistocerca Gregaria. Mycopathol. 1986, 96, 59–66. [Google Scholar] [CrossRef]
- Holder, D.J.; Keyhani, N.O. Adhesion of the entomopathogenic fungus Beauveria (Cordyceps) bassiana to substrata. Appl. Environ. Microbiol. 2005, 71, 5260–5266. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.B.; Marchini, L.C.; Pereira, R.M.; Baumgratz, L.L. Effects of some insect pathogens on the Africanized honey bee, Apis mellifera L. (Hym., Apidae). J. Appl. Entomol. 1996, 120, 559–564. [Google Scholar] [CrossRef]
- Amnuaykanjanasin, A.; Jirakkakul, J.; Panyasiri, C.; Panyarakkit, P.; Nounurai, P.; Chantasingh, D.; Eurwilaichitr, L.; Cheevadhanarak, S.; Tanticharoen, M. Infection and colonization of tissues of the aphid Myzus persicae and cassava mealybug Phenacoccus manihoti by the fungus Beauveria bassiana. BioControl 2013, 58, 379–391. [Google Scholar] [CrossRef]
- Ugelvig, L.V.; Cremer, S. Social prophylaxis: Group interaction promotes collective immunity in ant colonies. Curr. Biol. 2007, 17, 1967–1971. [Google Scholar] [CrossRef]
- Brighenti, D.M.; Carvalho, C.F.; Carvalho, G.A.; Brighenti, C.R.G.; Carvalho, S.M. Bioatividade do Bacillus thuringiensis var. kurstaki (Berliner, 1915) para adultos de Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae). Ciênc. E Agrotecnol. 2007, 31, 279–289. [Google Scholar] [CrossRef]
- Blacquiere, T.; Smagghe, G.; Van Gestel, C.A.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxic. 2012, 21, 973–992. [Google Scholar] [CrossRef]
- Cremer, S.; Pull, C.D.; Fürst, M.A. Social Immunity: Emergence and evolution of colony-level disease protection. Annu. Rev. Entomol. 2018, 63, 105–123. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Tosi, S.; Nieh, J.C.; Sgolastra, F.; Cabbri, R.; Medrzycki, P. Neonicotinoid pesticides and nutritional stress synergistically reduce survival in honey bees. Proc. R. Soc. B 2017, 284, 20171711. [Google Scholar] [CrossRef] [Green Version]
- Meikle, M.G.; Mercadier, G.; Holst, N.; Nansen, C.; Girod, V. Impact of a treatment of Beauveria bassiana (Deuteromycota: Hyphomycetes) on honeybee (Apis mellifera) colony health and on Varroa destructor mites (Acari: Varroidae). Apidologie 2008, 39, 247–259. [Google Scholar] [CrossRef]
- Kanga, L.H.; Adamczyk, J.; Patt, J.; Gracia, C.; Cascino, J. Development of a user-friendly delivery method for the fungus Metarhizium anisopliae to control the ectoparasitic mite Varroa destructor in honey bee, Apis mellifera, colonies. Exp. Appl. Acarol. 2010, 52, 327–342. [Google Scholar] [CrossRef]
- Rajula, J.; Karthi, S.; Mumba, S.; Pittarate, S.; Thungrabeab, M.; Krutmuang, P. Entomopathogenic fungi. In Recent Advancement in Microbial Biotechnology: Agricultural and Industrial Approach, 2nd ed.; Mandal, S., Passari, A.K., Eds.; Elsevier: London, UK, 2021; pp. 72–75. [Google Scholar]
- Cremer, S.; Armitage, S.A.; Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, 693–702. [Google Scholar] [CrossRef]
- Almeida, F.C.R.; Magalhães, D.M.; Favaris, A.P.; Rodríguez, J.; Azevedo, K.E.X.; Bento, J.M.S.; Alves, D.A. Side effects of a fungus-based biopesticide on stingless bee guarding behaviour. Chemosphere 2022, 287, 132147. [Google Scholar] [CrossRef]
- Menegatti, C.; Da Paixão Melo, W.G.; Carrão, D.B.; Oliveira, A.R.M.; Nascimento, F.S.; Lopes, N.P.; Pupo, M.T. Paenibacillus polymyxa associated with the stingless bee Melipona scutellaris produces antimicrobial compounds against entomopathogens. J. Chem. Ecol. 2018, 44, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- OECD. Test No. 213: Honeybees, Acute Oral Toxicity Test. Available online: https://www.oecd-ilibrary.org/environment/test-no-213-honeybees-acute-oral-toxicity-test_9789264070165-en (accessed on 10 July 2022).
- OECD. Test No. 237: Honey bee (Apis mellifera) Larval Toxicity Test, Single EXPOSURE. Available online: https://www.oecd-ilibrary.org/environment/test-no-237-honey-bee-apis-mellifera-larval-toxicity-test-single-exposure_9789264203723-en (accessed on 10 July 2022).
- Botina, L.L.; Bernardes, R.C.; Barbosa, W.F.; Lima, M.A.P.; Guedes, R.N.C.; Martins, G.F. Toxicological assessments of agrochemical effects on stingless bees (Apidae, Meliponini). MethodsX 2020, 7, 100906. [Google Scholar] [CrossRef]
- Cham, K.O.; Nocelli, R.C.; Borges, L.O.; Viana-Silva, F.E.C.; Tonelli, C.A.M.; Malaspina, O.; Menezes, C.; Rosa-Fontana, A.S.; Blochtein, B.; Freitas, B.M.; et al. Pesticide exposure assessment paradigm for stingless bees. Environ. Entomol. 2019, 48, 36–48. [Google Scholar] [CrossRef]
- Cabrera, A.R.; Almanza, M.T.; Cutler, G.C.; Fischer, D.L.; Hinarejos, S.; Lewis, G.; Nigro, D.; Olmstead, A.; Overmyer, J.; Potter, D.A.; et al. Initial recommendations for higher-tier risk assessment protocols for bumble bees, Bombus spp. (Hymenoptera: Apidae). Integr. Environ. Assess. Manag. 2016, 12, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Klinger, E.G.; Camp, A.A.; Strange, J.P.; Cox-Foster, D.; Lehmann, D.M. Bombus (Hymenoptera: Apidae) microcolonies as a tool for biological understanding and pesticide risk assessment. Environ. Entomol. 2019, 48, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Reinbacher, L.; Bacher, S.; Praprotnik, E.; Grabenweger, G. Standard non-target tests for risk assessment of plant protection products are unsuitable for entomopathogenic fungi—A proposal for a new protocol. J. Soils Sediments 2021, 21, 2357–2368. [Google Scholar] [CrossRef]
- Köhl, J.; Booij, K.; Kolnaar, R.; Ravensberg, W.J. Ecological arguments to reconsider data requirements regarding the environmental fate of microbial biocontrol agents in the registration procedure in the European Union. BioControl 2019, 64, 469–487. [Google Scholar] [CrossRef]
- Machado, A.C.P.; Barônio, G.J.; Oliveira, F.F.; Garcia, C.T.; Rech, A.R. Does a coffee plantation host potential pollinators when it is not flowering? Bee distribution in an agricultural landscape with high biological diversity in the Brazilian Campo Rupestre. J. Sci. Food Agri. 2021, 101, 2345–2354. [Google Scholar] [CrossRef] [PubMed]
- Van Oystaeyen, A.; Klatt, B.K.; Petit, C.; Lenaerts, N.; Wäckers, F. Short-term lab assessments and microcolonies are insufficient for the risk assessment of insecticides for bees. Chemosphere 2021, 273, 128518. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Fungi | Bee Species | p-Value | |||
|---|---|---|---|---|---|---|
| S. depilis | T. angustula | A. mellifera | B. terrestris | |||
| topical | B. bassiana | 0.76 a | 0.48 b | 0.36 b | 0.34 b | <0.0001 |
| C. fumosorosea | 0.48 a | 0.48 a | 0.30 ab | 0.16 b | 0.0008 | |
| M. anisopliae | 0.38 b | 0.50 a | 0.22 bc | 0.14 c | 0.0003 | |
| oral | B. bassiana | 0.80 a | 0.58 ab | 0.50 b | 0.66 ab | 0.0189 |
| C. fumosorosea | 0.66 b | 0.70 b | 0.58 b | 0.92 a | 0.0004 | |
| M. anisopliae | 0.64 a | 0.58 a | 0.28 b | 0.64 a | 0.0003 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, M.O.G.; Alves, D.A.; Lecocq, A.; Malaquias, J.B.; Delalibera, I., Jr.; Jensen, A.B. Laboratory Risk Assessment of Three Entomopathogenic Fungi Used for Pest Control toward Social Bee Pollinators. Microorganisms 2022, 10, 1800. https://doi.org/10.3390/microorganisms10091800
Leite MOG, Alves DA, Lecocq A, Malaquias JB, Delalibera I Jr., Jensen AB. Laboratory Risk Assessment of Three Entomopathogenic Fungi Used for Pest Control toward Social Bee Pollinators. Microorganisms. 2022; 10(9):1800. https://doi.org/10.3390/microorganisms10091800
Chicago/Turabian StyleLeite, Mariana O. G., Denise A. Alves, Antoine Lecocq, José Bruno Malaquias, Italo Delalibera, Jr., and Annette B. Jensen. 2022. "Laboratory Risk Assessment of Three Entomopathogenic Fungi Used for Pest Control toward Social Bee Pollinators" Microorganisms 10, no. 9: 1800. https://doi.org/10.3390/microorganisms10091800