
Bile Acids: Major Regulator of the Gut Microbiome
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inhibition of Bacteria by Bile Acid
2.1.1. Strains and Growth Conditions
2.1.2. Bile Acid Susceptibility Test
2.2. Bile Acid Administration to the Mouse Model
2.2.1. Animals
2.2.2. Experimental Design
2.2.3. Stool Collection and Real-Time Polymerase Chain Reaction (qRT-PCR)
2.2.4. Statistical Analysis
3. Results
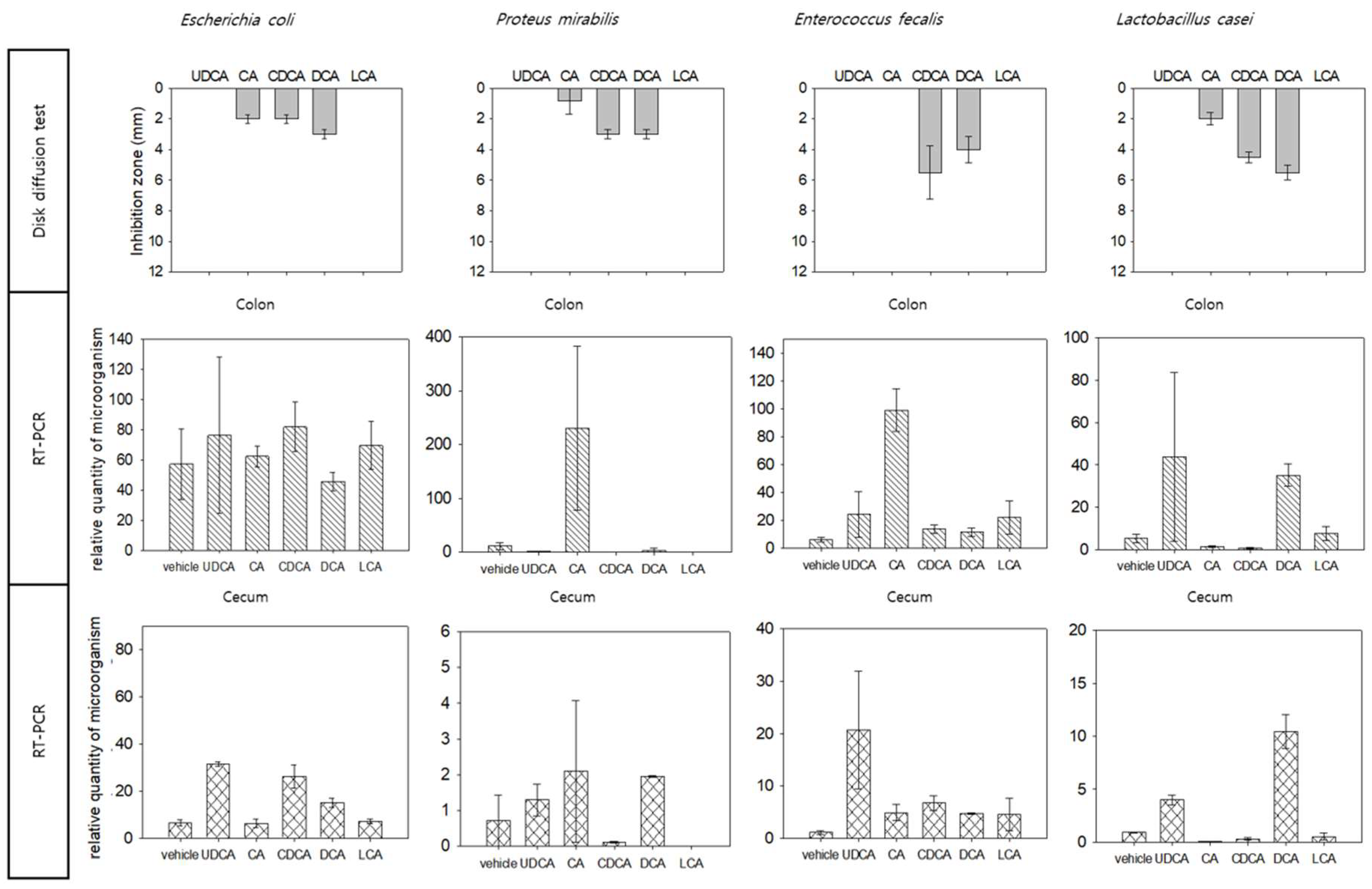
3.1. Inhibition of Bacteria by Bile Acid
3.2. Bile Acid Injection Model
3.3. Microbiota in Colon and Cecum
3.4. Comparative Analysis of In Vitro and In Vivo Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| UDCA | ursodeoxycholic acid |
| CA | cholic acid |
| CDCA | chenodeoxycholic acid |
| DCA | deoxycholic acid |
| LCA | lithocholic acid |
| qRT-PCR | real-time polymerase chain reaction |
| FXR | farnesoid X receptor |
| VDR | vitamin D receptor |
| IACUC | Institutional Animal Care and Use Committee |
| Cq | cycle quantification value |
| F/B ratio | Firmicutes/Bacteroidetes ratio |
| β-MCA | β-muricholic acid |
| HDCA | hyodeoxycholic acid |
| HHI | hydrophilic-hydrophobic index |
| β-HCA | β-hyocholic acid |
| α-MCA | α-muricholic acid |
| MDCA | murideoxycholic acid |
References
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef]
- Qi, Y.; Jiang, C.; Cheng, J.; Krausz, K.W.; Li, T.; Ferrell, J.M.; Gonzalez, F.J.; Chiang, J.Y. Bile acid signaling in lipid metabolism: Metabolomic and lipidomic analysis of lipid and bile acid markers linked to anti-obesity and anti-diabetes in mice. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 19–29. [Google Scholar] [CrossRef]
- Russell, D.W. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed]
- Leverrier, P.; Dimova, D.; Pichereau, V.; Auffray, Y.; Boyaval, P.; Jan, G. Susceptibility and adaptive response to bile salts in Propionibacterium freudenreichii: Physiological and proteomic analysis. Appl. Environ. Microbiol. 2003, 69, 3809–3818. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Mori, M. Influence of bile salts on β-glucuronidase activity of intestinal bacteria. Lett. Appl. Microbiol. 1996, 22, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Kandell, R.L.; Bernstein, C. Bile salt/acid induction of DNA damage in bacterial and mammalian cells: Implications for colon cancer. 1991. Nutr. Cancer 1991, 16, 227–238. [Google Scholar] [CrossRef]
- Bernstein, C.; Bernstein, H.; Payne, C.M.; Beard, S.E.; Schneider, J. Bile salt activation of stress response promoters in Escherichia coli. Curr. Microbiol. 1999, 39, 68–72. [Google Scholar] [CrossRef]
- Sanyal, A.J.; Hirsch, J.I.; Moore, E.W. Premicellar taurocholate enhances calcium uptake from all regions of rat small intestine. Gastroenterology 1994, 106, 866–874. [Google Scholar] [CrossRef]
- Rajagopalan, N.; Lindenbaum, S. The binding of Ca2+ to taurine-and glycine-conjugated bile salt micelles. Biochim. Et Biophys. Acta (BBA)-Lipids Lipid Metab. 1982, 711, 66–74. [Google Scholar] [CrossRef]
- Flahaut, S.; Frere, J.; Boutibonnes, P.; Auffray, Y. Comparison of the bile salts and sodium dodecyl sulfate stress responses in Enterococcus faecalis. Appl. Environ. Microbiol. 1996, 62, 2416–2420. [Google Scholar] [CrossRef] [PubMed]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a nuclear receptor for bile acids. Science 1999, 284, 1362–1365. [Google Scholar] [CrossRef] [PubMed]
- D’Aldebert, E.; Mve, M.J.B.B.; Mergey, M.; Wendum, D.; Firrincieli, D.; Coilly, A.; Fouassier, L.; Corpechot, C.; Poupon, R.; Housset, C. Bile salts control the antimicrobial peptide cathelicidin through nuclear receptors in the human biliary epithelium. Gastroenterology 2009, 136, 1435–1443. [Google Scholar] [CrossRef]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef]
- Hernández, S.B.; Cava, F.; Pucciarelli, M.G.; García-del Portillo, F.; de Pedro, M.A.; Casadesús, J. Bile-induced peptidoglycan remodelling in S almonella enterica. Environ. Microbiol. 2015, 17, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Thanassi, D.G.; Cheng, L.W.; Nikaido, H. Active efflux of bile salts by Escherichia coli. J. Bacteriol. 1997, 179, 2512–2518. [Google Scholar] [CrossRef]
- Pos, K.M. Drug transport mechanism of the AcrB efflux pump. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2009, 1794, 782–793. [Google Scholar] [CrossRef]
- Prieto, A.I.; Ramos-Morales, F.; Casadesus, J. Repair of DNA damage induced by bile salts in Salmonella enterica. Genetics 2006, 174, 575–584. [Google Scholar] [CrossRef]
- Hernández, S.B.; Cota, I.; Ducret, A.; Aussel, L.; Casadesús, J. Adaptation and preadaptation of Salmonella enterica to bile. PLoS Genet. 2012, 8, e1002459. [Google Scholar] [CrossRef]
- Kakiyama, G.; Pandak, W.M.; Gillevet, P.M.; Hylemon, P.B.; Heuman, D.M.; Daita, K.; Takei, H.; Muto, A.; Nittono, H.; Ridlon, J.M. Modulation of the fecal bile acid profile by gut microbiota in cirrhosis. J. Hepatol. 2013, 58, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Rost, D.; Rudolph, G.; Kloeters-Plachky, P.; Stiehl, A. Effect of high-dose ursodeoxycholic acid on its biliary enrichment in primary sclerosing cholangitis. Hepatology 2004, 40, 693–698. [Google Scholar] [CrossRef]
- Gray, H. Antomy of the human body. Ann. Surg. 1918, 68, 564–566. [Google Scholar] [CrossRef]
- Agur, A.; Lee, M. Grant’s Atlas of anatomy, 10th ed; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1999. [Google Scholar]
- Nguyen, T.L.A.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Models Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef]
- Hugenholtz, F.; de Vos, W.M. Mouse models for human intestinal microbiota research: A critical evaluation. Cell. Mol. Life Sci. 2018, 75, 149–160. [Google Scholar] [CrossRef]
- Wells, C.; Erlandsen, S.L. Localization of translocating Escherichia coli, Proteus mirabilis, and Enterococcus faecalis within cecal and colonic tissues of monoassociated mice. Infect. Immun. 1991, 59, 4693–4697. [Google Scholar] [CrossRef]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- Inagaki, T.; Moschetta, A.; Lee, Y.-K.; Peng, L.; Zhao, G.; Downes, M.; Yu, R.T.; Shelton, J.M.; Richardson, J.A.; Repa, J.J. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 3920–3925. [Google Scholar] [CrossRef]
- Bansal, S.; Singh, M.; Kidwai, S.; Bhargava, P.; Singh, A.; Sreekanth, V.; Singh, R.; Bajaj, A. Bile acid amphiphiles with tunable head groups as highly selective antitubercular agents. MedChemComm 2014, 5, 1761–1768. [Google Scholar] [CrossRef]
- Melzer, M.; Toner, R.; Lacey, S.; Bettany, E.; Rait, G. Biliary tract infection and bacteraemia: Presentation, structural abnormalities, causative organisms and clinical outcomes. Postgrad. Med. J. 2007, 83, 773–776. [Google Scholar] [CrossRef]
- Gu, S.; Chen, D.; Zhang, J.-N.; Lv, X.; Wang, K.; Duan, L.-P.; Nie, Y.; Wu, X.-L. Bacterial community mapping of the mouse gastrointestinal tract. PLoS ONE 2013, 8, e74957. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 15064–15069. [Google Scholar] [CrossRef]
- Berry, D.; Stecher, B.; Schintlmeister, A.; Reichert, J.; Brugiroux, S.; Wild, B.; Wanek, W.; Richter, A.; Rauch, I.; Decker, T. Host-compound foraging by intestinal microbiota revealed by single-cell stable isotope probing. Proc. Natl. Acad. Sci. USA 2013, 110, 4720–4725. [Google Scholar] [CrossRef]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063.e8. [Google Scholar] [CrossRef]
- De Aguiar Vallim, T.Q.; Tarling, E.J.; Edwards, P.A. Pleiotropic roles of bile acids in metabolism. Cell Metab. 2013, 17, 657–669. [Google Scholar] [CrossRef]
- Xie, G.; Zhong, W.; Li, H.; Li, Q.; Qiu, Y.; Zheng, X.; Chen, H.; Zhao, X.; Zhang, S.; Zhou, Z. Alteration of bile acid metabolism in the rat induced by chronic ethanol consumption. FASEB J. 2013, 27, 3583–3593. [Google Scholar] [CrossRef]
- Heuman, D.; Hylemon, P.; Vlahcevic, Z. Regulation of bile acid synthesis. III. Correlation between biliary bile salt hydrophobicity index and the activities of enzymes regulating cholesterol and bile acid synthesis in the rat. J. Lipid Res. 1989, 30, 1161–1171. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vehicle | CA | LCA | DCA | CDCA | UDCA | |
|---|---|---|---|---|---|---|
| Glucose, mg/dL | 334.7 ± 16.2 | 262.3 ± 13.0 | 268.3 ± 46.9 | 212.3 ± 32.9 | 214.0 ± 28.4 | 245.7 ± 17.7 |
| Total cholesterol, mg/dL | 103.3 ± 6.2 | 114.7 ± 6.1 | 105.3 ± 13.0 | 107.5 ± 14.7 | 108.3 ± 11.1 | 119.3 ± 12.5 |
| GPT, U/L | 7.7 ± 0.3 | 8.0 ± 1.0 | 6.7 ± 0.9 | 6.0 ± 0.6 | 12.3 ± 4.1 | 6.0 ± 0.6 |
| T.bil, mg/dL | 1.9 ± 0.3 | 1.7 ± 0.5 | 1.7 ± 0.1 | 1.2 ± 0.2 | 3.2 ± 0.8 | 1.6 ± 0.0 |
| ALP, U/L | 124.7 ± 4.5 | 116.7 ± 15.9 | 164.3 ± 14.5 | 187.0 ± 1.7 | 116.7 ± 34.1 | 129.3 ± 33.1 |
| GGT, U/L | 6.7 ± 5.2 | 5.0 ± 2.6 | 1.0 ± 0.0 | 1.0 ± 0.2 | 31.3 ± 18.9 | 5.3 ± 4.3 |
| TG, mg/dL | 65.3 ± 16.2 | 58.0 ± 7.8 | 39.0 ± 3.0 | 35.0 ± 1.2 | 44.0 ± 8.0 | 33.7 ± 6.0 |
| Albumin, g/dL | 2.5 ± 0.1 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.3 ± 0.2 | 2.4 ± 0.2 | 2.1 ± 0.1 |
| HDLC, mg/dl | 89.5 ± 0.9 | 96.0 ± 2.5 | 96.7 ± 6.9 | 106.0 ± 2.3 | 104.0 ± 4.0 | 96.7 ± 6.8 |
| GOT, U/L | 312.5 ± 43.6 | 376.0 ± 100.7 | 250.3 ± 69.7 | 142.3 ± 1.8 | 146.0 ± 2.3 | 227.0 ± 1.2 |
| Total protein, g/dL | 5.0 ± 0.9 | 5.1 ± 0.1 | 4.9 ± 0.8 | 5.3 ± 0.4 | 5.4 ± 0.2 | 4.7 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, C.; Chon, H.; Ku, W.; Eom, S.; Seok, M.; Kim, S.; Lee, J.; Kim, D.; Lee, S.; Koo, H.; et al. Bile Acids: Major Regulator of the Gut Microbiome. Microorganisms 2022, 10, 1792. https://doi.org/10.3390/microorganisms10091792
An C, Chon H, Ku W, Eom S, Seok M, Kim S, Lee J, Kim D, Lee S, Koo H, et al. Bile Acids: Major Regulator of the Gut Microbiome. Microorganisms. 2022; 10(9):1792. https://doi.org/10.3390/microorganisms10091792
Chicago/Turabian StyleAn, Chihyeok, Hyeyeon Chon, Wanrim Ku, Sunho Eom, Mingyu Seok, Sangha Kim, Jaesun Lee, Daesung Kim, Sanghyuk Lee, Hoonsup Koo, and et al. 2022. "Bile Acids: Major Regulator of the Gut Microbiome" Microorganisms 10, no. 9: 1792. https://doi.org/10.3390/microorganisms10091792