
Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida auris and Klebsiella pneumoniae
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains and Culture Conditions
2.3. Determination of Minimum Inhibitory Concentration (MIC)
2.4. Time–Kill Curves
2.5. Biofilm Formation and Characterization
2.6. Minimum Biofilm Inhibitory Concentration (MBIC)
2.7. qRT-PCR Analysis
2.8. Measurement of Intracellular ROS Levels and Mitochondrial Specific ROS Accumulation
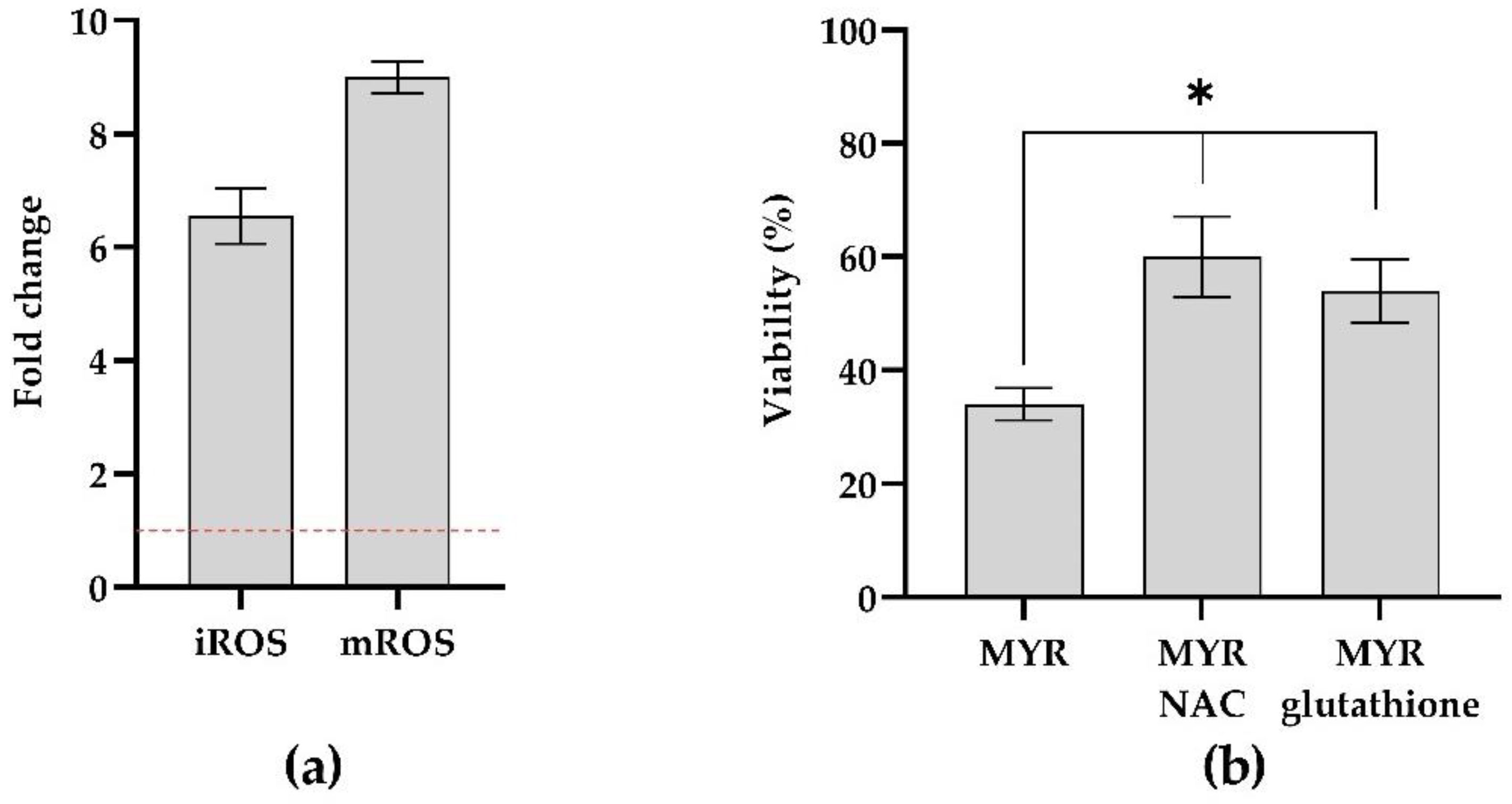
2.9. Cell Rescue Assay Using ROS Scavengers
2.10. Checkerboard Microdilution Assay
2.11. MTT Assay
2.12. Galleria Mellonella Assays: Toxicity, Infection Rescue Assay
2.13. Statistical Analysis
3. Results
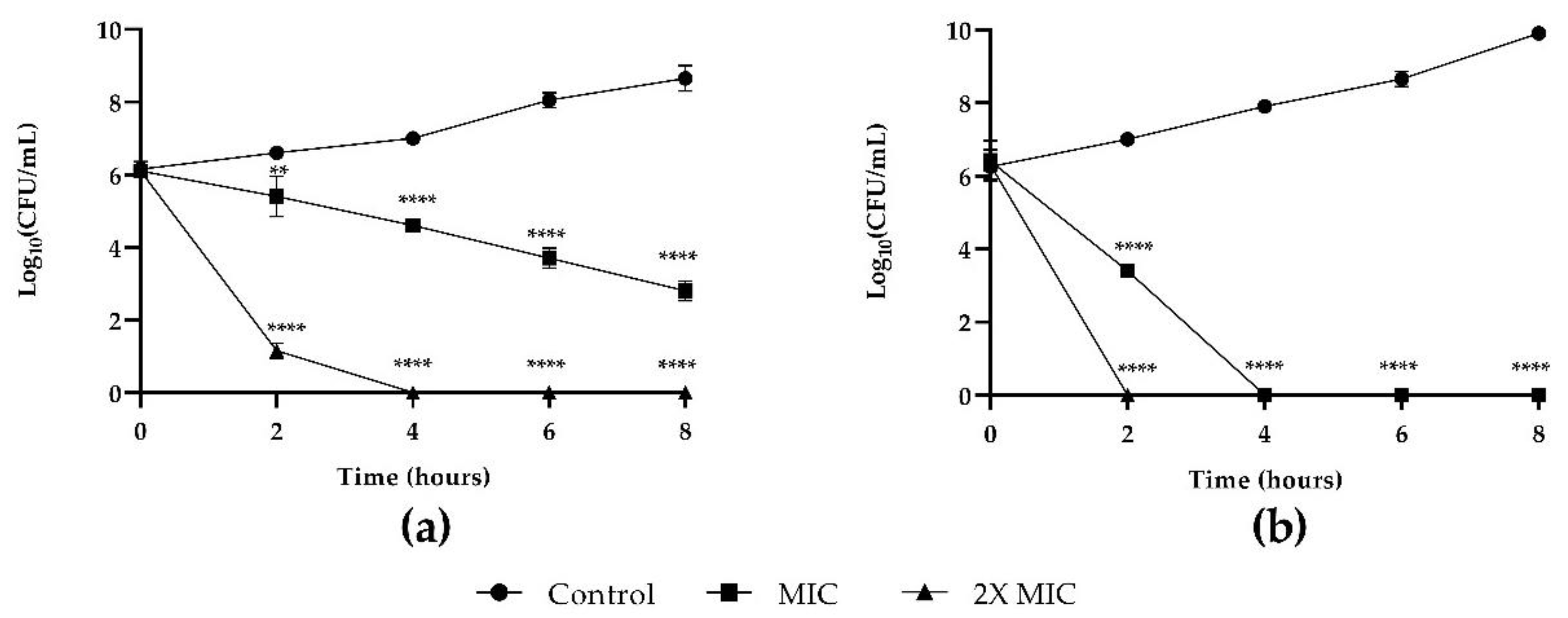
3.1. Susceptibility Testing of Planktonic Populations of C. auris and K. pneumoniae
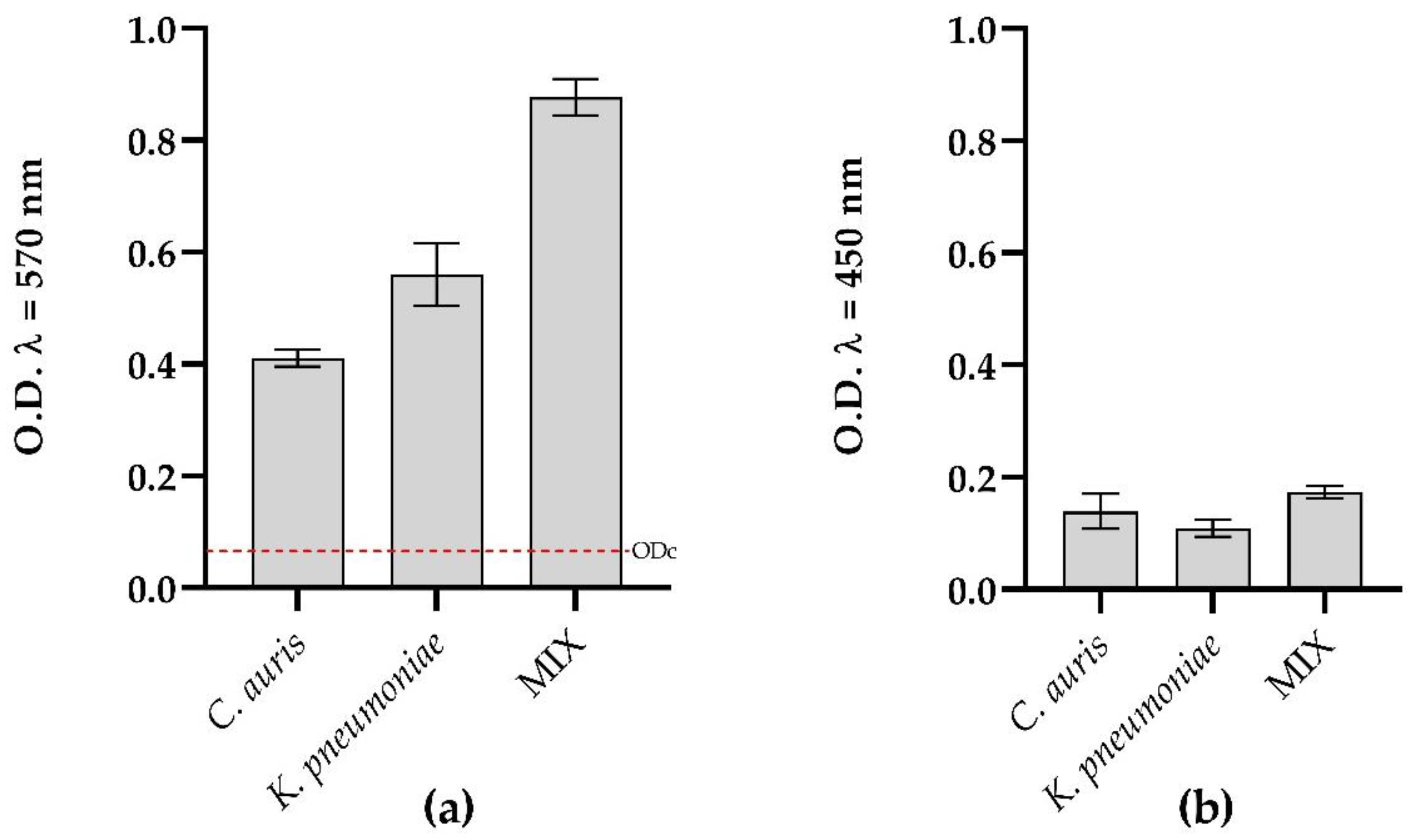
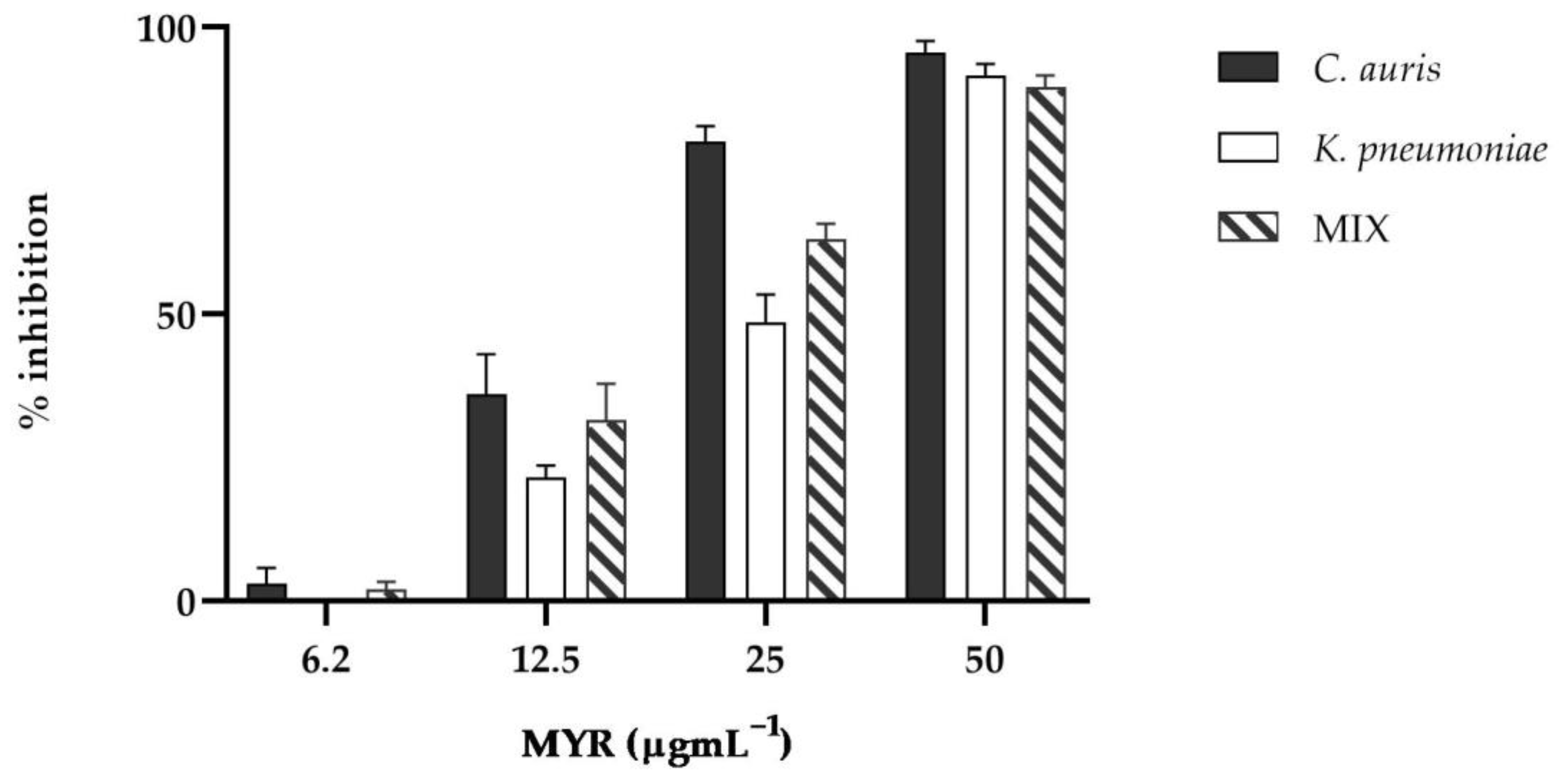
3.2. Effect of Myrtenol on C. auris and K. pneumoniae Biofilms
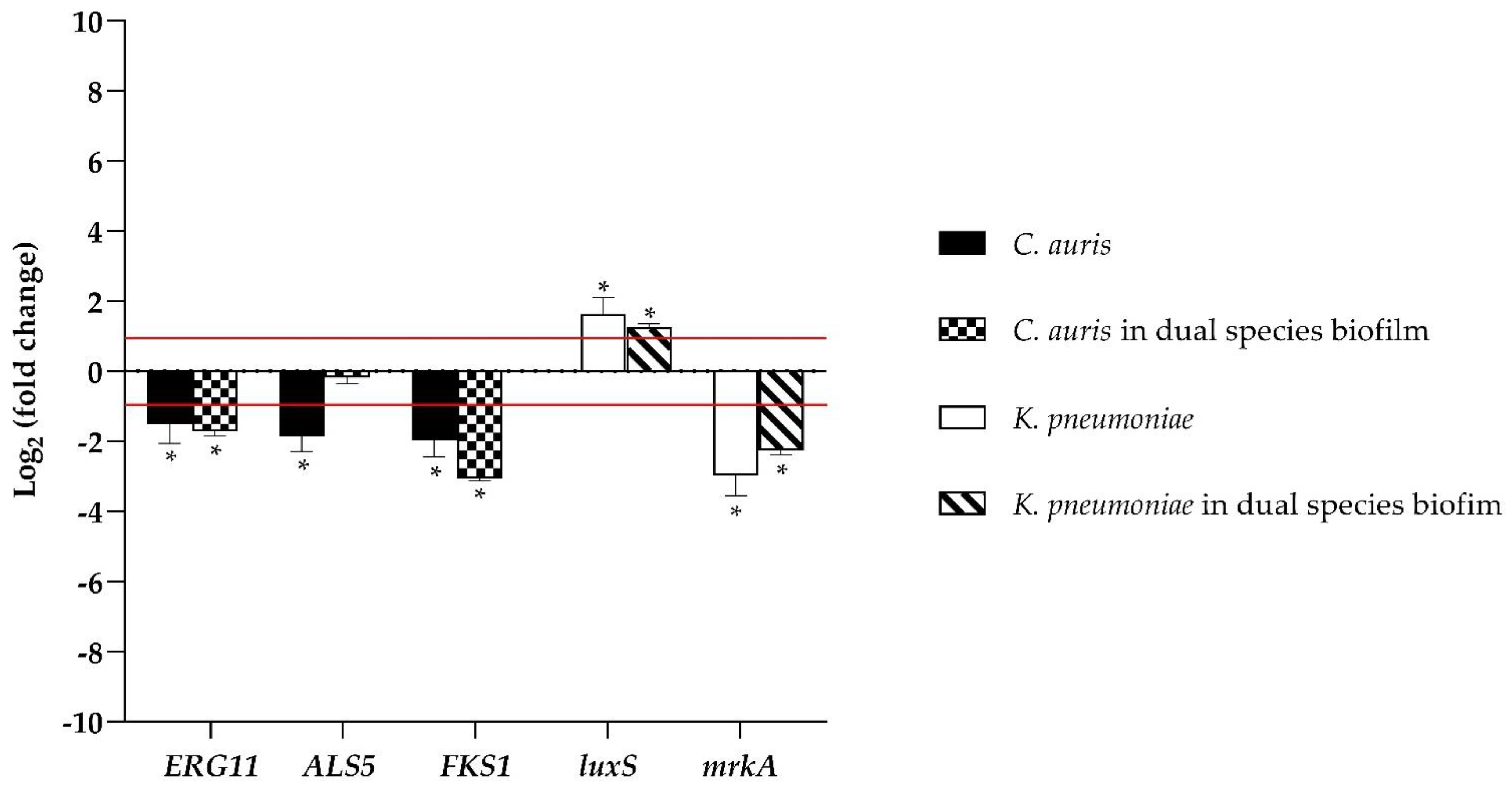
3.3. Myrtenol Inhibited the Expression of Biofilm Formation Related Genes
3.4. Measuring ROS Production in Mixed Biofilm
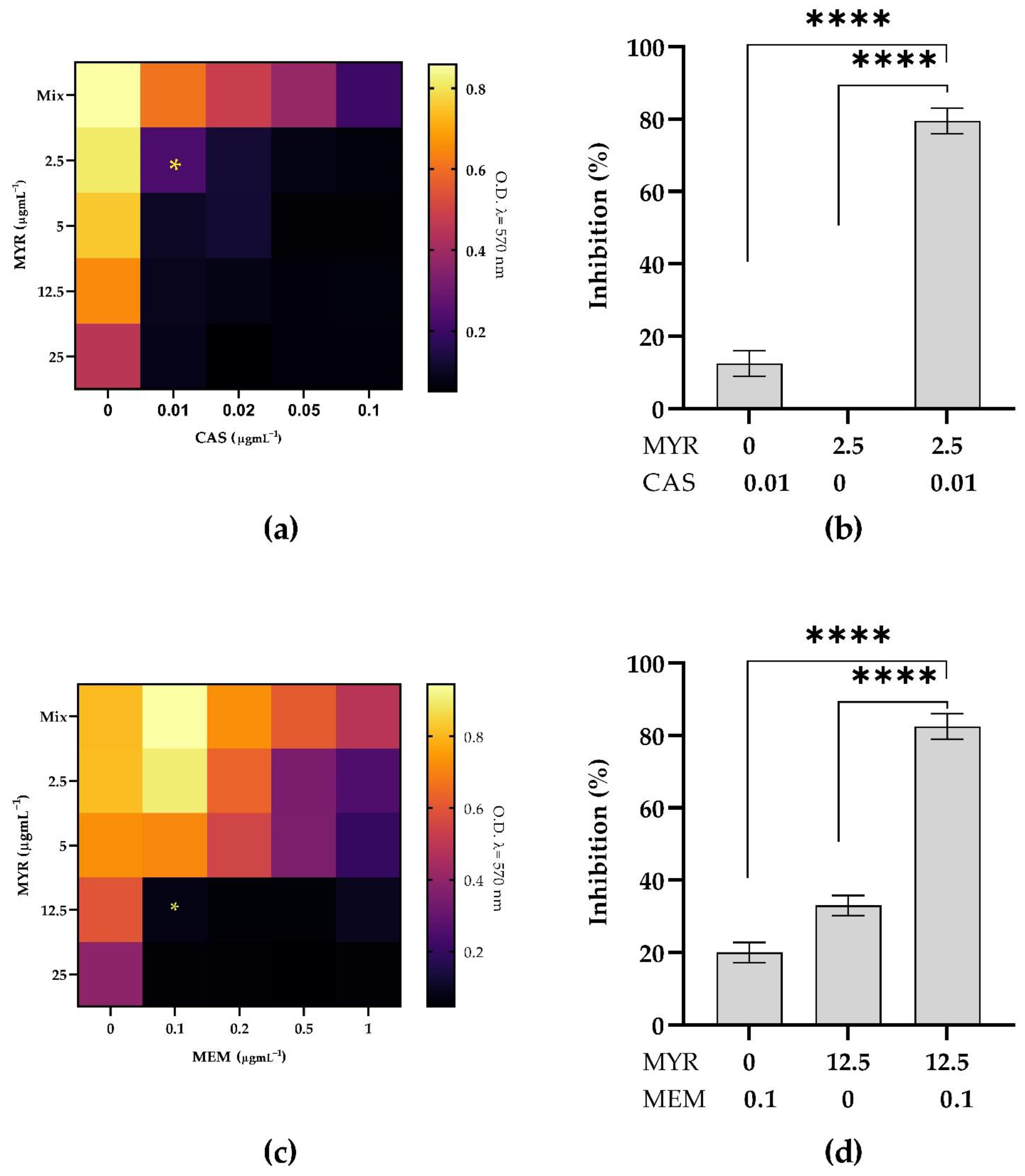
3.5. Combined Antimicrobial Effect on Inhibition of Mixed Biofilms
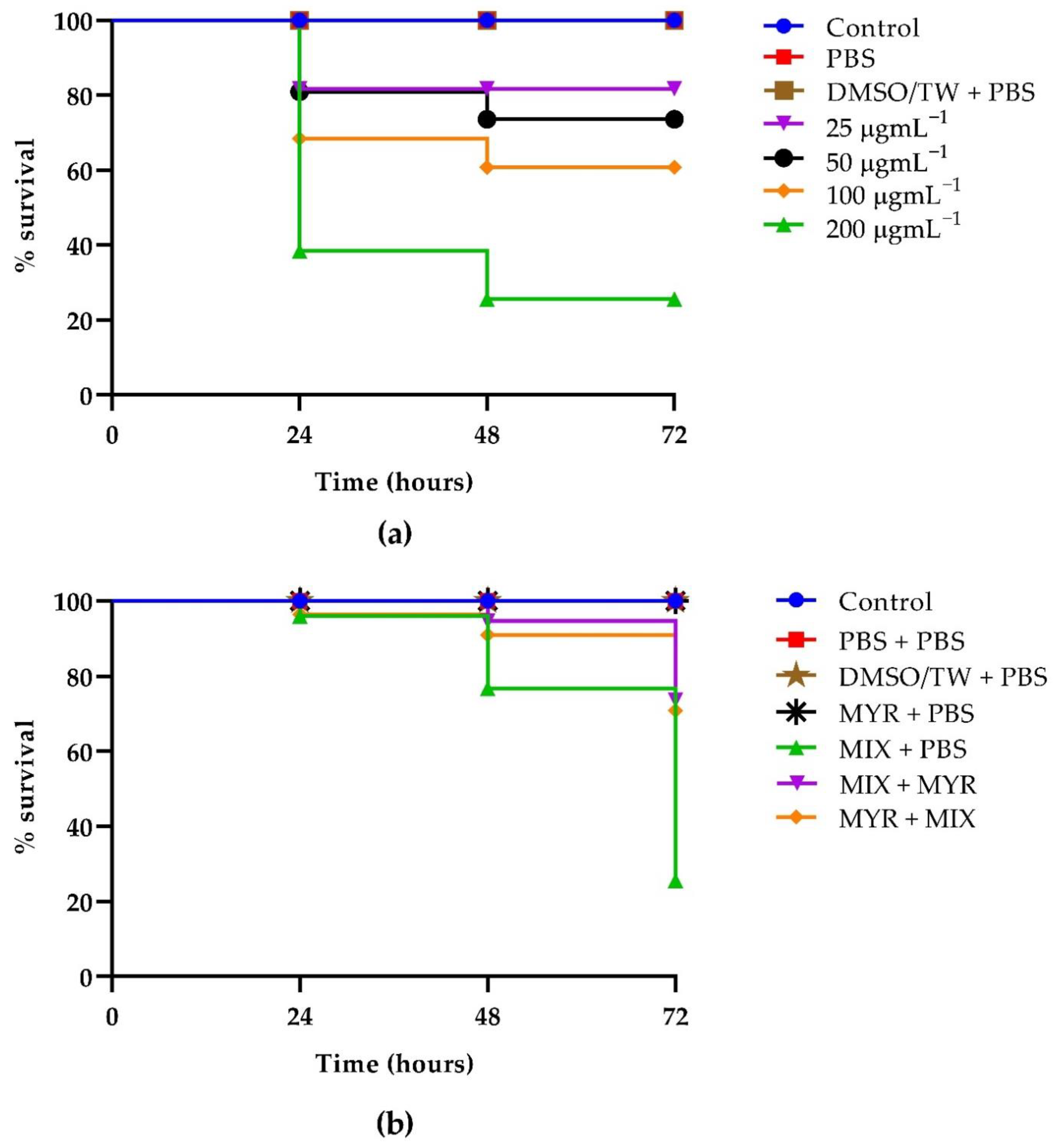
3.6. Kaplan–Meier Survival Curves
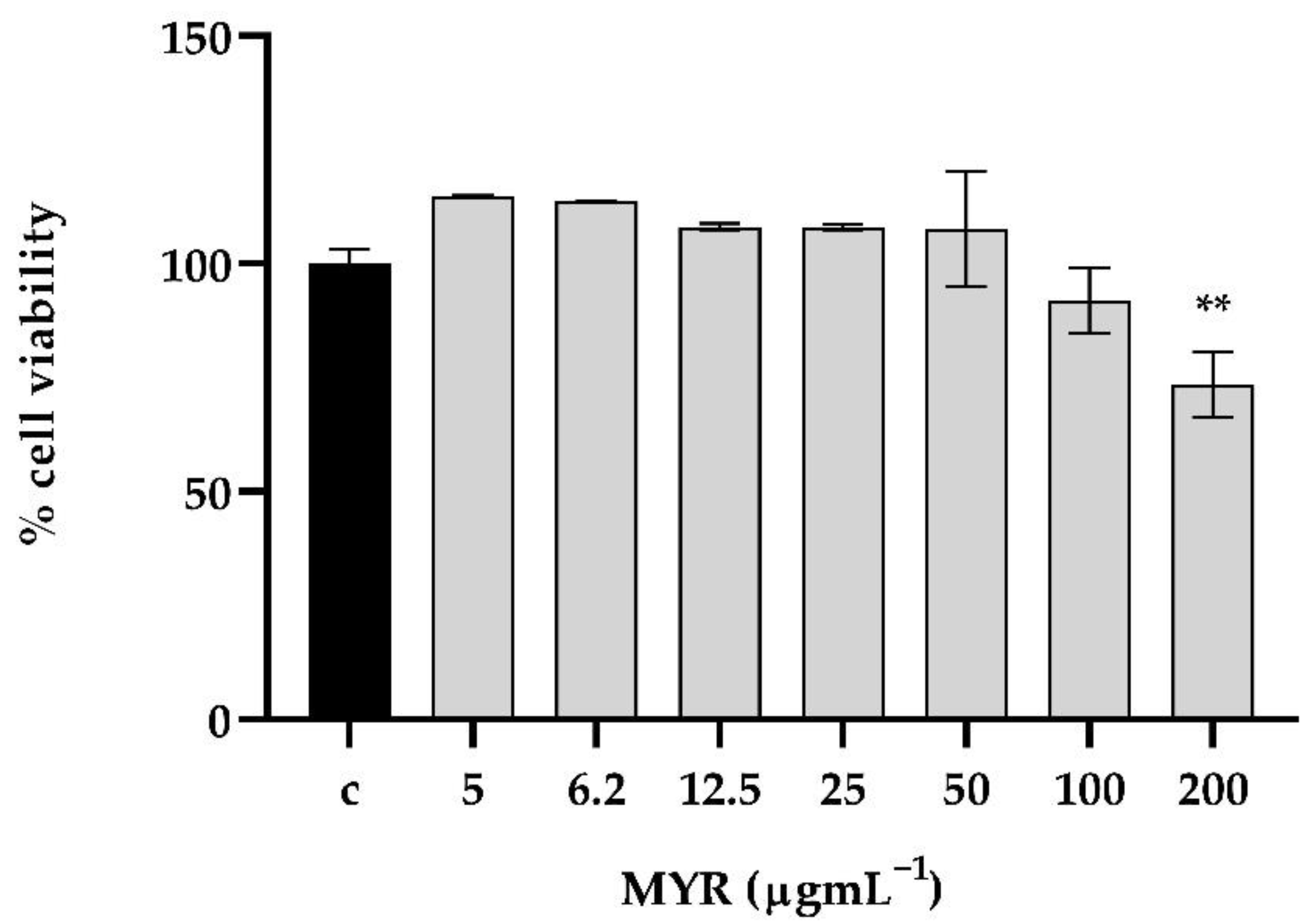
3.7. The Tested Concentrations of Myrtenol Exhibited No Considerable Cytotoxicity on In Vitro Human Keratinocyte Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dos Santos Pedroso, R.; Balbino, B.L.; Andrade, G.; Dias, M.C.P.S.; Alvarenga, T.A.; Pedroso, R.C.N.; Pimenta, L.P.; Lucarini, R.; Pauletti, P.M.; Januário, A.H.; et al. In vitro and in vivo anti-Candida spp. activity of plant-derived products. Plants 2019, 8, 494. [Google Scholar] [CrossRef] [PubMed]
- Serra, E.; Hidalgo-Bastida, L.A.; Verran, J.; Williams, D.; Malic, S. Antifungal Activity of Commercial Essential Oils and Biocides against Candida Albicans. Pathogens 2018, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, R.; Milho, C.; Liberal, Â.; Silva, J.; Fonseca, C.; Barbosa, A.; Ferreira, I.C.F.R.; Alves, M.J.; Barros, L. Antibiofilm Potential of Medicinal Plants against Candida spp. Oral Biofilms: A Review. Antibiotics 2021, 10, 1142. [Google Scholar] [CrossRef]
- Baharvand-Ahmadi, B.; Bahmani, M.; Naghdi, N.; Saki, K.; Baharvand-Ahmadi, S.; Rafieian-Kopaei, M. Review on phytochemistry, therapeutic and pharmacological effects of myrtus (Myrtus communis). Der Pharm. Lett. 2015, 7, 160–165. [Google Scholar]
- Gomes, B.S.; Neto, B.P.; Lopes, E.M.; Cunha, F.V.; Araújo, A.R.; Wanderley, C.W.; Wong, D.V.; Júnior, R.C.P.; Ribeiro, R.A.; Sousa, D.P.; et al. Anti-inflammatory effect of the monoterpene myrtenol is dependent on the direct modulation of neutrophil migration and oxidative stress. Chem.-Biol. Interact. 2017, 273, 73–81. [Google Scholar] [CrossRef]
- Paknejadi, M.; Foroohi, F.; Yousefzadi, M. Antimicrobial activities of the essential oils of five Salvia species from Tehran province, Iran. Arch. Adv. Biosci. 2012, 3, 12–18. [Google Scholar]
- Coté, H.; Boucher, M.-A.; Pichette, A.; Legault, J. Anti-Inflammatory, Antioxidant, Antibiotic, and Cytotoxic Activities of Tanacetum vulgare L. Essential Oil and Its Constituents. Medicines 2017, 4, 34. [Google Scholar] [CrossRef]
- Galdiero, E.; Di Onofrio, V.; Maione, A.; Gambino, E.; Gesuele, R.; Menale, B.; Ciaravolo, M.; Carraturo, F.; Guida, M. Allium ursinum and Allium oschaninii against Klebsiella pneumoniae and Candida albicans mono-and polymicrobic biofilms in in vitro static and dynamic models. Microorganisms 2020, 8, 336. [Google Scholar] [CrossRef]
- Gambino, E.; Maione, A.; Guida, M.; Albarano, L.; Carraturo, F.; Galdiero, E.; Di Onofrio, V. Evaluation of the Pathogenic-Mixed Biofilm Formation of Pseudomonas aeruginosa/Staphylococcus aureus and Treatment with Limonene on Three Different Materials by a Dynamic Model. Int. J. Environ. Res. Public Health 2022, 19, 3741. [Google Scholar] [CrossRef]
- De Alteriis, E.; Maione, A.; Falanga, A.; Bellavita, R.; Galdiero, S.; Albarano, L.; Salvatore, M.M.; Galdiero, E.; Guida, M. Activity of Free and Liposome-Encapsulated Essential Oil from Lavandula angustifolia against Persister-Derived Biofilm of Candida auris. Antibiotics 2022, 11, 26. [Google Scholar] [CrossRef]
- Rossatto, F.C.P.; Tharmalingam, N.; Escobar, I.E.; d’Azevedo, P.A.; Zimmer, K.R.; Mylonakis, E. Antifungal Activity of the Phenolic Compounds Ellagic Acid (EA) and Caffeic Acid Phenethyl Ester (CAPE) against Drug-Resistant Candida auris. J. Fungi 2021, 7, 763. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.J.; Eix, E.F.; Lam, B.C.; Wartman, K.M.; Meudt, J.J.; Shanmuganayagam, D.; Nett, J.E. Augmenting the Activity of Chlorhexidine for Decolonization of Candida auris from Porcine skin. J. Fungi 2021, 7, 804. [Google Scholar] [CrossRef] [PubMed]
- Kordalewska, M.; Perlin, D.S. Identification of drug resistant Candida auris. Front. Microbiol. 2019, 10, 1918. [Google Scholar] [CrossRef] [PubMed]
- Kubiczek, D.; Raber, H.; Gonzalez-García, M.; Morales-Vicente, F.; Staendker, L.; Otero-Gonzalez, A.J.; Rosenau, F. Derivates of the antifungal peptide Cm-p5 inhibit development of Candida auris biofilms in vitro. Antibiotics 2020, 9, 363. [Google Scholar] [CrossRef]
- Da Silva, M.A.; Baronetti, J.L.; Páez, P.L.; Paraje, M.G. Oxidative imbalance in Candida tropicalis biofilms and its relation with persister cells. Front. Microbiol. 2021, 11, 598834. [Google Scholar] [CrossRef]
- Tutelyan, A.V.; Shlykova, D.S.; Voskanyan, S.L.; Gaponov, A.M.; Pisarev, V.M. Molecular Epidemiology of Hypervirulent K. pneumoniae and Problems of Health-Care Associated Infections. Bull. Exp. Biol. Med. 2022, 172, 507–522. [Google Scholar] [CrossRef]
- Guerra, M.E.S.; Destro, G.; Vieira, B.; Lima, A.S.; Ferraz, L.F.C.; Hakansson, A.P.; Darrieux, M.; Converso, T.R. Klebsiella pneumoniae Biofilms and Their Role in Disease Pathogenesis. Front. Cell. Infect. Microbiol. 2022, 12, 877995. [Google Scholar] [CrossRef]
- Wikler, M.A. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically: Approved standard. CLSI 2006, 26, M7-A7. [Google Scholar]
- Fothergill, A.W. Antifungal susceptibility testing: Clinical laboratory and standards institute (CLSI) methods. In Interactions of Yeasts, Moulds, and Antifungal Agents; Springer: Berlin/Heidelberg, Germany, 2012; pp. 65–74. [Google Scholar]
- Maione, A.; de Alteriis, E.; Carraturo, F.; Galdiero, S.; Falanga, A.; Guida, M.; Di Cosmo, A.; Maselli, V.; Galdiero, E. The Membranotropic Peptide gH625 to Combat Mixed Candida albicans/Klebsiella pneumoniae Biofilm: Correlation between In Vitro Anti-Biofilm Activity and In Vivo Antimicrobial Protection. J. Fungi 2021, 7, 26. [Google Scholar] [CrossRef]
- Flamm, R.K.; Farrell, D.J.; Rhomberg, P.R.; Scangarella-Oman, N.E.; Sader, H.S. Gepotidacin (GSK2140944) in vitro activity against Gram-positive and Gram-negative bacteria. Antimicrob. Agents Chemother. 2017, 61, e00468-17. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Maione, A.; Bellavita, R.; de Alteriis, E.; Galdiero, S.; Albarano, L.; La Pietra, A.; Guida, M.; Parrilli, E.; D’Angelo, C.; Galdiero, E.; et al. WMR Peptide as Antifungal and Antibiofilm against Albicans and Non-Albicans Candida Species: Shreds of Evidence on the Mechanism of Action. Int. J. Mol. Sci. 2022, 23, 2151. [Google Scholar] [CrossRef] [PubMed]
- Falanga, A.; Maione, A.; La Pietra, A.; de Alteriis, E.; Vitale, S.; Bellavita, R.; Carotenuto, R.; Turrà, D.; Galdiero, S.; Galdiero, E.; et al. Competitiveness during Dual-Species Biofilm Formation of Fusarium oxysporum and Candida albicans and a Novel Treatment Strategy. Pharmaceutics 2022, 14, 1167. [Google Scholar] [CrossRef]
- Galdiero, E.; Ricciardelli, A.; D’Angelo, C.; de Alteriis, E.; Maione, A.; Albarano, L.; Casillo, A.; Corsaro, M.M.; Tutino, M.; Parrilli, E. Pentadecanoic acid against Candida albicans-Klebsiella pneumoniae biofilm: Towards the development of an anti-biofilm coating to prevent polymicrobial infections. Res. Microbiol. 2021, 172, 103880. [Google Scholar] [CrossRef]
- Di Onofrio, V.; Gesuele, R.; Maione, A.; Liguori, G.; Liguori, R.; Guida, M.; Nigro, R.; Galdiero, E. Prevention of Pseudomonas aeruginosa Biofilm Formation on Soft Contact Lenses by Allium sativum Fermented Extract (BGE) and Cannabinol Oil Extract (CBD). Antibiotics 2019, 8, 258. [Google Scholar] [CrossRef]
- Ivanov, M.; Ćirić, A.; Stojković, D. Emerging Antifungal Targets and Strategies. Int. J. Mol. Sci. 2022, 23, 2756. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Chang, C.-K.; Kao, M.-C.; Lan, C.-Y. Antimicrobial activity of the peptide LfcinB15 against Candida albicans. J. Fungi 2021, 7, 519. [Google Scholar] [CrossRef]
- Tan, J.; Jiang, S.; Tan, L.; Shi, H.; Yang, L.; Sun, Y.; Wang, X. Antifungal activity of minocycline and azoles against fluconazole-resistant Candida species. Front. Microbiol. 2021, 12, 649026. [Google Scholar] [CrossRef] [PubMed]
- Bellavita, R.; Maione, A.; Merlino, F.; Siciliano, A.; Dardano, P.; De Stefano, L.; Galdiero, S.; Galdiero, E.; Grieco, P.; Falanga, A. Antifungal and Antibiofilm Activity of Cyclic Temporin L Peptide Analogues against Albicans and Non-Albicans Candida Species. Pharmaceutics 2022, 14, 454. [Google Scholar] [CrossRef] [PubMed]
- Billamboz, M.; Fatima, Z.; Hameed, S.; Jawhara, S. Promising Drug Candidates and New Strategies for Fighting against the Emerging Superbug Candida auris. Microorganisms 2021, 9, 634. [Google Scholar] [CrossRef]
- Khan, Z.; Ahmad, S.; Al-Sweih, N.; Joseph, L.; Alfouzan, W.; Asadzadeh, M. Increasing prevalence, molecular characterization and antifungal drug susceptibility of serial Candida auris isolates in Kuwait. PLoS ONE 2018, 13, e0195743. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Herrera, N.; Lopez-Ribot, J.L. Repositionable compounds with antifungal activity against multidrug resistant Candida auris identified in the medicines for malaria venture’s pathogen box. J. Fungi 2019, 5, 92. [Google Scholar] [CrossRef]
- Hassoun-Kheir, N.; Snitser, O.; Hussein, K.; Rabino, G.; Eluk, O.; Warman, S.; Aboalhega, W.; Geffen, Y.; Mendelson, S.; Kishony, R. Concordance between epidemiological evaluation of probability of transmission and whole genome sequence relatedness among hospitalized patients acquiring Klebsiella pneumoniae carbapenemase-producing Klebsiella pneumoniae. Clin. Microbiol. Infect. 2021, 27, 468.e1–468.e7. [Google Scholar] [CrossRef]
- Sisay, M.; Gashaw, T. Ethnobotanical, ethnopharmacological, and phytochemical studies of Myrtus communis Linn: A popular herb in Unani system of medicine. J. Evid.-Based Complement. Altern. Med. 2017, 22, 1035–1043. [Google Scholar] [CrossRef]
- Asllani, U. Chemical composition of Albanian myrtle oil (Myrtus communis L.). J. Essent. Oil Res. 2000, 12, 140–142. [Google Scholar] [CrossRef]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials—A review. Plants 2017, 6, 16. [Google Scholar] [CrossRef]
- Selvaraj, A.; Jayasree, T.; Valliammai, A.; Pandian, S.K. Myrtenol attenuates MRSA biofilm and virulence by suppressing sarA expression dynamism. Front. Microbiol. 2019, 10, 2027. [Google Scholar] [CrossRef]
- Dwivedi, S.; Wahab, R.; Khan, F.; Mishra, Y.K.; Musarrat, J.; Al-Khedhairy, A.A. Reactive oxygen species mediated bacterial biofilm inhibition via zinc oxide nanoparticles and their statistical determination. PLoS ONE 2014, 9, e111289. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Acronym | Primer Name | Sequence (5′→3′) |
|---|---|---|---|
| S-ribosylhomocysteine lyase | luxS | K.pneumoniae_luxS_F | ATCGACATTTCGCCAATGGG |
| K.pneumoniae_luxS_R | ACTGGTAGACGTTGAGCTCC | ||
| Type 3 fimbrial shaft | mrkA | K.pneumoniae_mrkA_F | ACGTCTCTAACTGCCAGGC |
| K.pneumoniae_mrkA_R | TAGCCCTGTTGTTTGCTGGT | ||
| 16S ribosomial RNA | 16S rRNA | K.pneumoniae_16S_F | AGCACAGAGAGCTTG |
| K.pneumoniae_16S_R | ACTTTGGTCTTGCGAC | ||
| 1,3-beta-glucan synthase | FKS1 | C.auris_ FKS1_F | GCAAACTTTCATGTTGGTGTTA |
| C.auris_ FKS1_R | TGTGAACAAGGAGTTTGAGTAA | ||
| Ergosterol Biosynthesis | ERG11 | C.auris_ERG11_F | GTGCCCATCGTCTACAACCT |
| C.auris_ERG11_R | TCTCCCACTCGATTTCTGCT | ||
| Hyphal-specific genes | ALS5 | C.auris_ALS5_F | CCTTCTGGATCGGACACAGT |
| C.auris_ALS5_R | AGTTGTGGTGGAGGAACCAG | ||
| Actin | actin | C. auris_actin_F | GAAGGAGATCACTGCTTTAGCC |
| C.auris_actin_R | GAGCCACCAATCCACACAG |
| Strains | CFU/Well ± SD | % Composition Dual-Species Biofilm | |
|---|---|---|---|
| Single-Species Biofilm | Dual-Species Biofilm | ||
| C. auris | (3.0 ± 0.05) × 106 | (6.3 ± 0.12) × 105 | 6 |
| K. pneumoniae | (6.0 ± 0.15) × 106 | (1.0 ± 0.03) × 107 | 94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maione, A.; La Pietra, A.; de Alteriis, E.; Mileo, A.; De Falco, M.; Guida, M.; Galdiero, E. Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida auris and Klebsiella pneumoniae. Microorganisms 2022, 10, 1773. https://doi.org/10.3390/microorganisms10091773
Maione A, La Pietra A, de Alteriis E, Mileo A, De Falco M, Guida M, Galdiero E. Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida auris and Klebsiella pneumoniae. Microorganisms. 2022; 10(9):1773. https://doi.org/10.3390/microorganisms10091773
Chicago/Turabian StyleMaione, Angela, Alessandra La Pietra, Elisabetta de Alteriis, Aldo Mileo, Maria De Falco, Marco Guida, and Emilia Galdiero. 2022. "Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida auris and Klebsiella pneumoniae" Microorganisms 10, no. 9: 1773. https://doi.org/10.3390/microorganisms10091773