1. Introduction
Soda lakes of the East African Rift Valley are unique habitats characterized by stable, high pH in the alkaline range, and high salinity. Alkalinity is a result of high carbonate-bicarbonate concentration, which also provides an unlimited carbon dioxide reserve for photosynthesis. In addition, because of their tropical location, these lakes get unlimited light all year round, and experience little temperature fluctuation. As a result, these lakes are highly productive, with the highest primary production reported for any natural habitat [
1,
2]. High primary production also implies an abundance of organic matter released due to cell death or excess photosynthesis, which, in turn, supports a diverse group of heterotrophic microorganisms [
3].
Lake Chitu is one of the soda lakes found in the central Rift Valley of Ethiopia. It is a crater lake with a stable pH of around 10.5 and salinity of about 6% [
4,
5]. The blue-green algae,
Arthrospira fusiformis (formerly known as
Spirulina platensis or commonly as Spirulina), is the most dominant primary producer in the lake [
4]. Oxygen measurement showed that, depending on the biomass concentration on the surface and the mixing rate, below a depth of 1–3 m (and sometimes below a depth of half a meter) from the surface, the lake is totally anoxic. Therefore, throughout the year, a significant portion of the lake’s depth is anoxic.
A study using culture-independent molecular techniques revealed the presence of high microbial diversity in Lake Chitu. Furthermore, both microbial diversity and abundance increased with increasing depth [
5], which is probably a result of an increased availability of organic matter released from dead algal cells sinking downward. This could also indicate that the available organic matter is utilized by a diverse group of microorganisms distributed along the entire depth of the lake. Such microbial degradation of organic matter involves hydrolysis of macromolecules, such as proteins, DNA, RNA, lipids, and carbohydrates to their respective monomeric units, such as amino acids, fatty acids, and simple sugars. Further oxidation of these monomeric units under aerobic and anaerobic conditions could lead to their conversion to CO
2, nitrate, and sulfate.
The main primary producer in Lake Chitu,
Arthrospira fusiformis, is well known for its high protein content [
6]. During its active growth cycle, the entire lake appears covered by a thick green biomass, indicating active growth. Dense algal growth requires an unlimited supply of all the required nutrients and an availability of light energy. Since Lake Chitu has a high carbonate bicarbonate concentration and is found in the tropics, the supply of CO
2 and access to energy (sun light) could not be a limiting factor on the rate of photosynthesis. However, in addition to the above inputs, an accumulation of a large quantity of biomass also requires an adequate supply of nitrogen sources for the synthesis of amino acids, nucleotides, and other nitrogen-containing organic compounds.
The lake is essentially a closed basin with no surface inflow and outflow. The only known inflows are a few hot springs that spring from the shore of the lake, and drain directly into the lake. Therefore, the lake does not get any external nitrogen supply, be it in the form of organic or inorganic nitrogen. One assumption that could be put forward is a closed nitrogen cycle within the lake, where ammonia is released upon the degradation of organic matter (for example, proteins, DNA, and RNA), and is released as ammonia and nitrate, which will then be taken up by the primary producer and other heterotrophic microorganisms in the lake.
However, the assumption of a closed nitrogen cycle within the lake could only be true in the absence of denitrification. A recent study in our laboratory on Lake Chitu demonstrated the presence of a diverse group of denitrifying bacteria, many of them exhibiting a high efficiency of denitrification [
7]. Since the bulk of the water column (up to 95%) is devoid of oxygen, the presence of a diverse group of denitrifying prokaryotes may indicate that the nitrate and nitrite released during the degradation process of nitrogen-containing organic compounds could serve as a final electron acceptor, leading to its release as molecular nitrogen, N
2. If nitrate and nitrite of the lake are lost as N
2 through the process of denitrification, the question regarding the source of nitrogen to support prolific cyanobacterial growth in such a closed environment remains unanswered.
Therefore, if nitrate is lost due to denitrification, the other potential source of nitrogen to support algal growth in this closed environment could be nitrogen fixation. Biological nitrogen fixation involves the reduction of N
2 into ammonia (NH
3), the only inorganic nitrogen that can be assimilated by living organisms. Only a few groups of prokaryotes belonging to the domains, archaea and bacteria, can fix atmospheric nitrogen. These diazotrophic microbes possess the enzyme, nitrogenase, a complex biocatalyst consisting of several subunits that are involved in nitrogen fixation. Nitrogen-fixing microorganisms could exist as free-living organisms in terrestrial and aquatic habitats, or form symbiotic associations with other organisms with varying degrees of complexity, and are a vital source of nitrogen in any ecosystem [
8]. Therefore, the nitrogen economy of Lake Chitu might depend on biological nitrogen fixation.
Most known nitrogen-fixing prokaryotes, whether free-living or symbiotic, grow under neutral pH and in the presence of low or moderate concentrations of salt. On the other hand, the environment in Lake Chitu is alkaline and saline. Therefore, prokaryotes that exist in this lake are subjected to double extreme conditions. The aim of this study was, therefore, to investigate the presence and diversity of culturable nitrogen-fixing alkaliphiles and their potential role in the nitrogen economy of the soda lake habitat.
2. Materials and Methods
2.1. Sampling Site and Sample Collection
Lake Chitu is located in the central Rift Valley of Ethiopia at 7°24′0″ N and 38°25′0.02″ E. It is a small crater lake with a surface area of approximately 800 m2, a maximum depth of 17 m, and salinity of about 6%. A few hot springs emerge from the shore and drain into the lake. Other than these hot springs and direct precipitation, the lake is a closed basin with no outflow and inflow.
Water and sediment samples were collected in triplicate from different depths, including the surface (0 m) and the sediment from a depth of 17 m. At the time of sampling, the transition between aerobic and anaerobic zones was determined to be at 2.9 m, and this was selected as one of the sampling depths. The other sampling depths include surface sample (0 m), which represents the oxic zone, 5, 10, 14, and 17 m, all representing the anoxic zone.
Water and sediment samples were collected using a bottle sampler and an Ekman grab, respectively. Samples were immediately transferred to sterile bottles and tubes, and transported to the laboratory in an icebox kept at 4 °C. Physicochemical parameters, including pH, salinity, conductivity, and temperature, were measured on site at the time of sampling.
2.2. Enrichment Cultures
To isolate nitrogen-fixing bacteria, a nitrogen-free media was prepared following the method of Jensen [
9] with slight modification. The medium was composed of (g/L): glucose, 5; NaCl, 5; MgSO
4·7H
2O, 0.2; K
2HPO
4, 0.1; CaCl
2·2H
2O, 0.26; FeSO
4, 0.1; Na
2CO
3, 10; and trace mineral solution, 10 mL/L. Glucose, Na
2CO
3, and trace mineral salt solution were sterilized separately and added to the rest of the medium after cooling to around 50 to 60 °C. The stock trace metal salt solution was composed of (g/L): H
3BO
3, 0.03; NaMO
4·2H
2O, 0.003; MnCl
2·4H
2O, 0.003; ZnSO
4·7H
2O, 0.01; CoCl
2·6H
2O; 0.01; NiCl·6H
2O, 0.002; and CaCl
2·2H
2O, 0.001. After adjusting the pH to 7.6 using 1M NaOH, the medium was autoclaved at 121 °C for 15 min. For solid media, agar was added to the above media to a final concentration of 20 g/L.
Enrichment was carried out by inoculating 100 mL sterile liquid media with 1 mL of a well-mixed water. For sediment samples, about 1 g was first suspended in 100 mL sterile diluent, and 1 mL of the suspension was used to inoculate 100 mL medium. The culture was incubated at 30 °C for up to seven days under aerobic and anaerobic conditions.
2.3. Isolation of Nitrogen-Fixing Bacterial Strains
The enrichment culture was serially diluted in the range of 10−1 to 10−5 using filter-sterilized saline solution, pH adjusted to 10. A 100 µL sample was spread onto agar plates containing nitrogen-free medium. Plates were incubated at 30 °C under aerobic and anaerobic conditions until distinct colonies emerged. A total of 180 colonies were randomly picked and purified through repeated streaking. Isolates were kept on nitrogen-rich agar slants at 4 °C for further characterization.
2.4. Co-Culture of Nitrogen-Fixing Bacterial Isolates with Non-Nitrogen-Fixing Isolate
The release of reduced nitrogen in the form of ammonia by the nitrogen-fixing alkaliphiles was measured indirectly through co-culturing with a known non-nitrogen-fixing alkaliphile, Alkalibacterium sp. 3.5R*1. When it is alone, Alkalibacterium sp. 3.5R*1 was unable to grow in the nitrogen-free medium. However, when it is cocultured with nitrogen-fixing strains, the organism grows well in the nitrogen-free medium, indicating an uptake of reduced nitrogen released in the medium. Another advantage of using Alkalibacterium sp. 3.5R*1 was that it has a marked color difference from the cocultured nitrogen-fixing isolates, making it easier to distinguish and enumerate the two when grown mixed on agar plates.
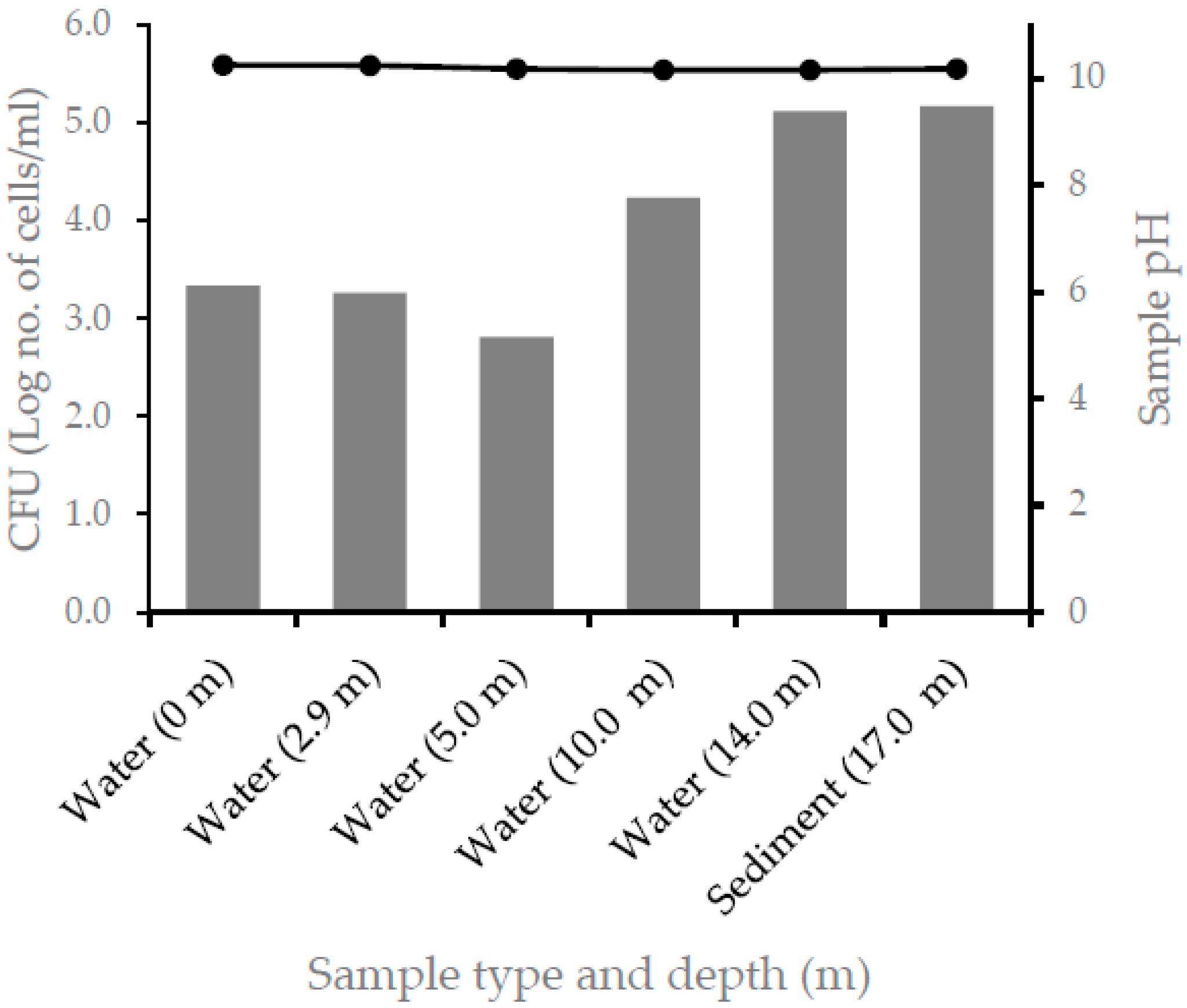
For the coculturing experiment, one of the alkaliphilic nitrogen-fixing isolate, NF10m6, was used. Thus, the nitrogen-fixing isolate, NF10m6, and the non-nitrogen-fixing strain, Alkalibacterium sp. 3.5R*1 (JX434738), were inoculated in a liquid nitrogen-free medium both independently (single culture) and the two together (co-culture), and incubated at 30 °C for ten days under anaerobic conditions. A sample of each culture was taken and serially diluted and spread on a nitrogen-rich solid medium and incubated as described above. In each plate, colonies of the nitrogen-fixing and non-nitrogen-fixing strains were enumerated based on their distinct colony color, and expressed as CFU/mL, and the result is expressed as log number of cells/mL.
2.5. Genomic DNA Extraction
Genomic DNA from 118 pure culture isolates was extracted following the modified freeze-thaw DNA extraction method [
10]. Few colonies were picked from an agar plate using a sterile loop and re-suspended in 50 μL of TE buffer, pH 8.0 in 1.5 mL Eppendorf tubes. The tubes were in a boiling water bath for 5 min, followed by immediate freezing at −20 °C for 15 min. The boiling and freezing steps were repeated once again. The cell lysate was stored at −20 °C and used for PCR amplification.
2.6. PCR Amplification and Amplified Ribosomal DNA Restriction Analysis (ARDRA)
PCR amplification of the 16S rRNA gene and endonuclease restriction digestion of the amplified 16S rRNA gene were carried out as described previously [
11]. For PCR amplification of the 16S rRNA universal bacterial primer sets, B338f (5′-ACTCCTACGGGAGGCAGCAG-3′) and H1542r (5′-TGCGGCTGGATCACCTCCTT-3′) primers [
12] were used. Restriction digestion was carried out using the restriction enzyme, TaqI (5′…T^CGA…3′).
2.7. Sequencing of Selected Isolates
Twenty representative isolates were selected from each OTU group and refreshed on nitrogen-free media for the extraction of DNA. PCR amplification and sequencing of the 760 bp 16S rRNA gene fragment were carried as described previously [
11] using primer sets, A8f and H1542r [
12].
2.8. PCR Detection and Cloning of nifH Gene
PCR detection of the
nifH gene was carried on the nineteen representative isolates selected randomly from each OTU group. PCR amplification of the
nifH gene was carried by a nested PCR approach as described previously [
13]. The first round of the nested PCR was carried out using
nifH primer sets,
nifH4 (
Azotobacter vinelandii nucleotide positions 546 to 562; 5′-TTY TAY GGN AAR GGN GG-3′) and
nifH3 (
A. vinelandii nucleotide positions 1018 to 1002; 5′-ATR TTR TTN GCN GCR TA-3′). The second round of the nested PCR was performed using primer sets, nifH1 (
A. vinelandii nucleotide positions 639 to 655; 5′-TGYGAYCCNAAR GCN GA-3′) and
nifH2 (
A.
vinelandii nucleotide positions 1000 to 984; 5′-ANDGCCTCATYTCNCC-3′). The presence and amplification of
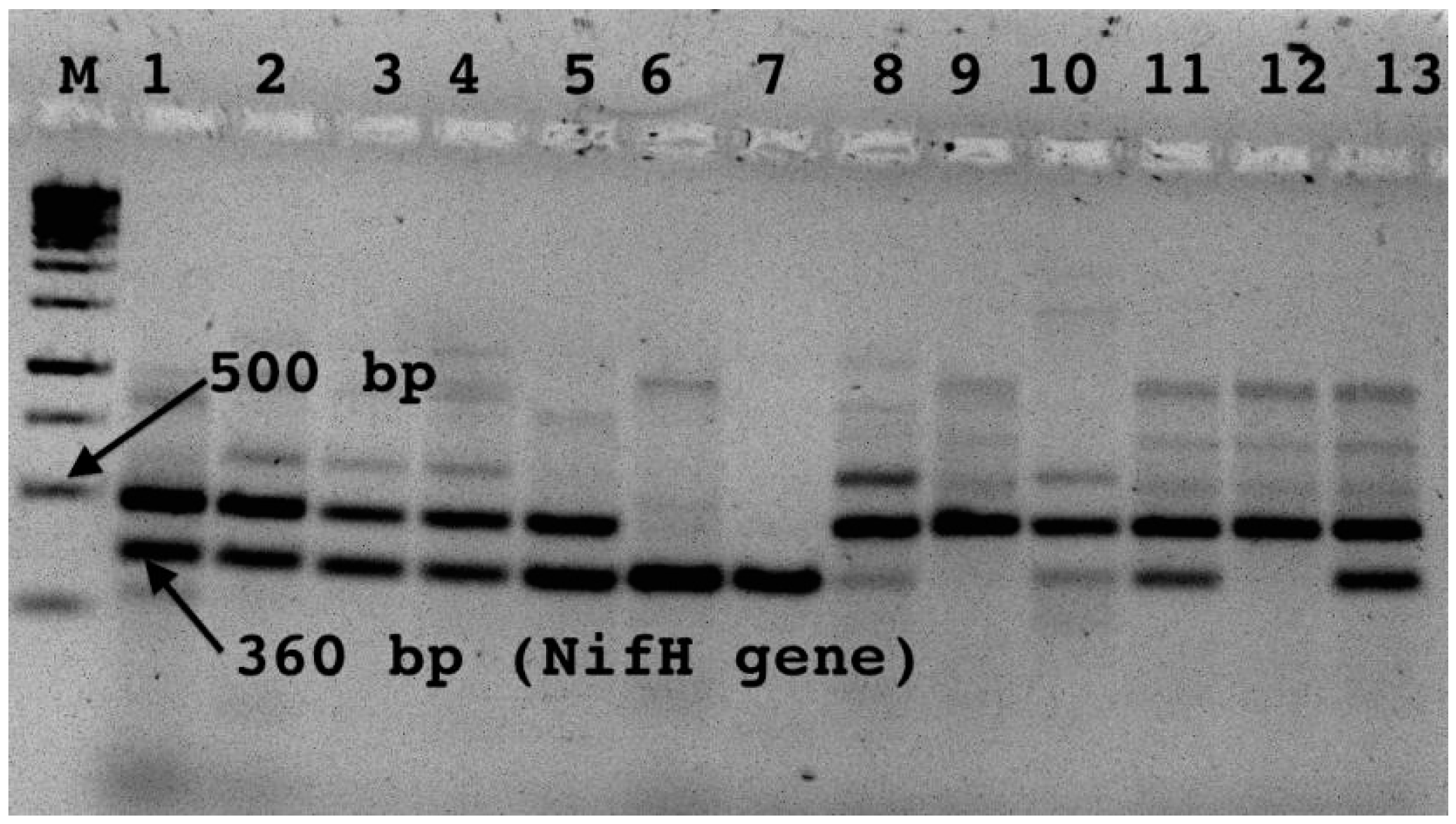
nifH of the gene was detected by running the amplicons on 2.5% agarose gel electrophoresis. Bands, which correspond to the
nifH gene at around 360 bp, were excised and gel-purified by using a gel Nucleospine purification kit according to the manufacturer’s instructions (Nucleospine extraction kit 2004). The elute was preserved at 4 °C for cloning and further analysis.
2.9. Cloning of nifH Gene, Reamplification, and Sequencing of Cloned nifH Gene Products
Depending on the band intensity, 3–4 µL of the gel-purified fragment of the PCR-amplified nifH gene (approximately 360 bp) was ligated into the plasmid vector (pCR® 2.1-TOPO® vector, 3.9 kb) using a TOPO® TA Cloning Kit (Invitrogen, Waltham, MA, USA, 2006) according to the manufacturer’s instruction. The heat shock method of transformation on One Shot® Chemically Competent E. coli cells (TOPO10) and the screening of transformed cells were carried out as described in TOPO TA Cloning Kit (Invitrogen, 2006).
Cloned PCR products with the right insert were screened by PCR amplification using M13 primers directly from overnight cultured transformed E. coli cells, and by running the amplified PCR products on agarose gel (3% w/v) electrophoresis. Clones with the right insert were sequenced using primer sets, nifH1 and nifH2, from both directions.
2.10. Phylogenetic Analysis
Sequences were edited manually using CLUSTALW [
14]. The program, Bellerophon on Detection, of chimeric sequences in multiple alignments was applied to the studied sequences in order to detect possible chimeric artifacts [
15]. Reference 16S rRNA sequences were retrieved from the NCBI GenBank database available at
http://www.ncbi.nlm.nih.gov (accessed on 1 April 2022) using BLAST (Basic Local Alignment Search Tool) analysis to provide the identity of the sequence. The phylogenetic relationship of sequences to the closest matches in the public database based on 16S RNA gene sequences was constructed by using the maximum likelihood method using distances calculated with the Kimura 2-parameter model [
16] in MEGA 10.0 software [
17]. The stability and reliability of the relationships of lineages on the inferred trees was tested by bootstrap analysis for 1000 replicates [
18]. Sequences with sufficient length were included in the alignment; the sequence alignments were then corrected manually, and sequences with ambiguous alignment positions were removed from the analysis.
2.11. Sequence Deposition
The 16S rRNA and nifH genes’ nucleotide sequences were submitted to the NCBI nucleotide database under accession numbers, OL444760-OL444777 (16S rRNA gene) and OL606585-OL606586 (nifH gene).
4. Discussion
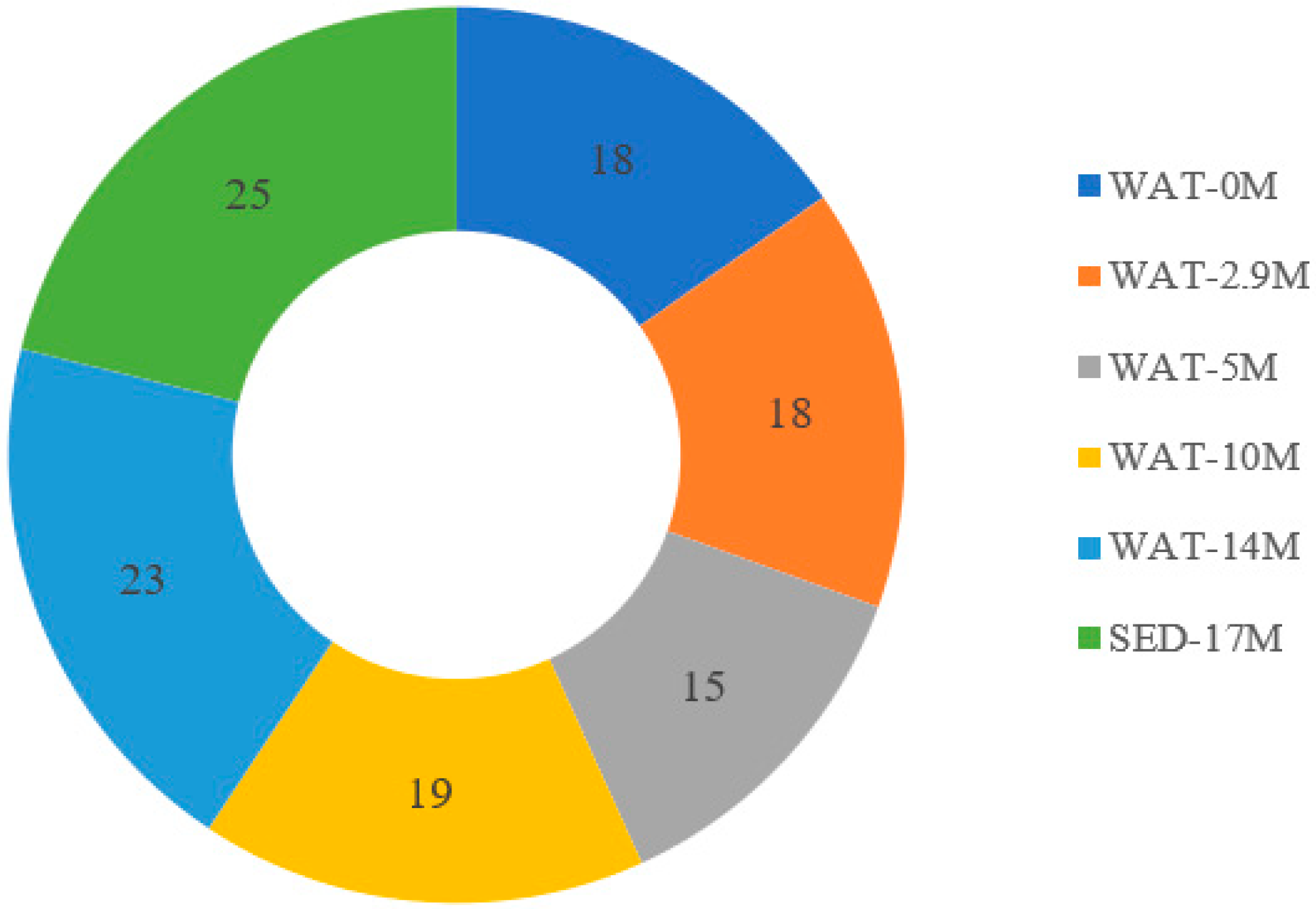
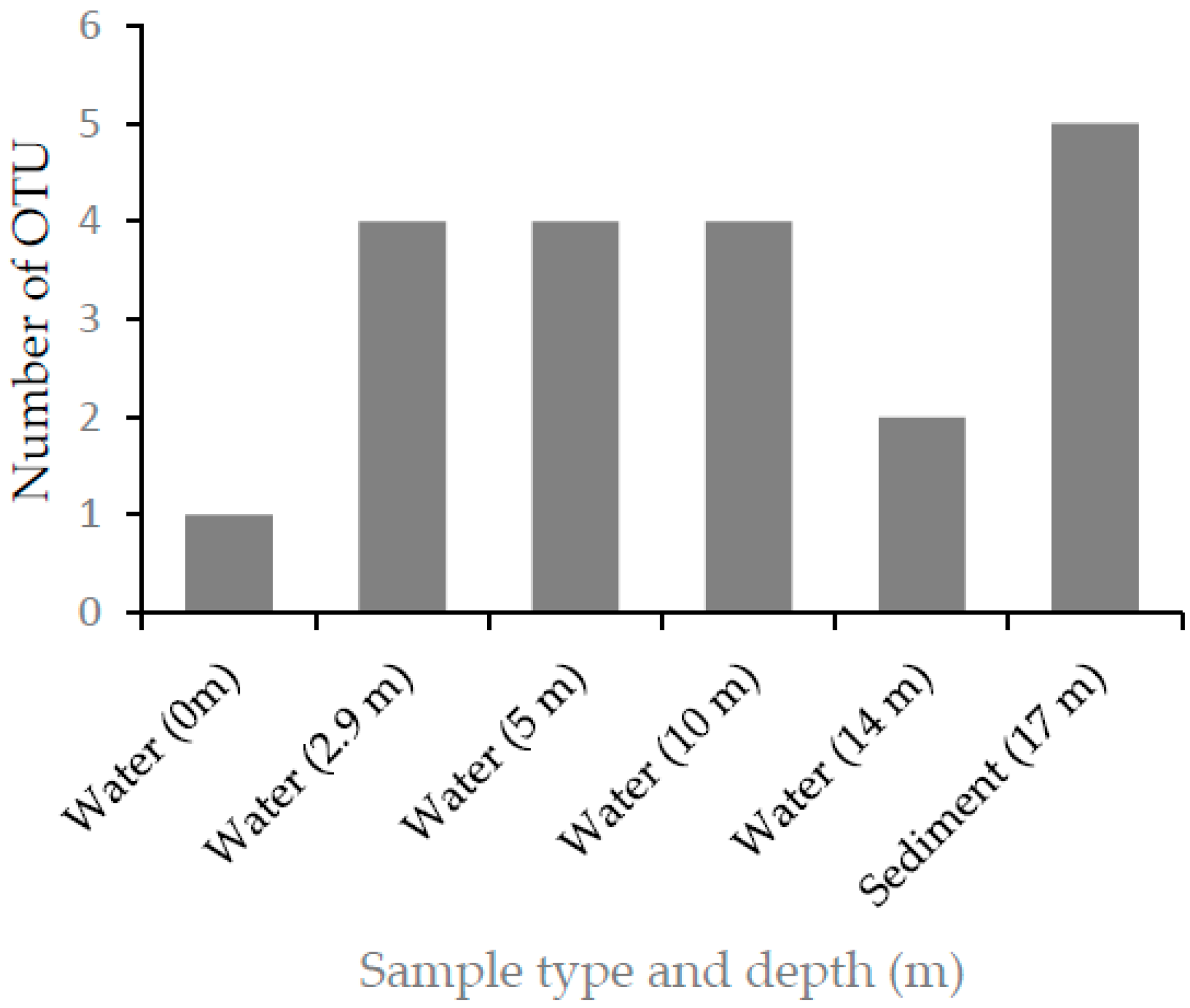
In this study, alkaliphilic bacterial isolates that grew on a nitrogen-free medium were isolated from samples collected from different depths of Lake Chitu, and were grouped into nine different OTUs. This shows that nitrogen-fixing prokaryotes probably have a significant contribution in the nitrogen economy of the lake.
Lake Chitu is a crater lake with no inflow and outflow other than a few hot springs that emerge from the shore and enter the lake, and the source of nitrogen to support the high biomass of the primary producer and a diverse group of heterotrophic microorganisms remains unclear. An abundance of denitrifiers, as revealed in a recent study [
7], indicates a potential loss of nitrate available in the lake. This is especially important when one considers the fact that a significant portion of the lake (below about 0.5–3 m out of a maximum depth of 17) is devoid of oxygen. Hence, an abundance of denitrifying microorganisms show that nitrate released from the degradation of organic nitrogen could be released as molecular nitrogen. Therefore, to support such prolific growth observed in Lake Chitu, nitrogen fixation by different alkaliphiles might play an important role in supplying the required nitrogen.
In addition to their ability to grow on nitrogen-free medium, the alkaliphilic isolates from Lake Chitu possess the
nifH gene that code for the nitrogenase enzyme involved in nitrogen fixation. This shows that the alkaliphilic isolates have the genetic potential to fix nitrogen. However, the possession of a nitrogenase gene may not necessarily show the existence of a functional nitrogenase enzyme capable of fixing nitrogen under the alkaline and saline conditions of the lake. In this study, the ability to fix nitrogen was indirectly tested by co-culturing with a known non-nitrogen-fixing bacterial strain which was unable to grow on a nitrogen-free medium. Expression of the nitrogenase gene of nitrogen-fixing prokaryotes is regulated by the availability of nitrogen in the surrounding environment [
22]. When the gene is turned on and the cell fixes the atmospheric nitrogen, it is not only fixing enough for itself, but also releases excess nitrogen in the form of ammonia used by other organisms in its vicinity [
23], which is also the basis for the symbiotic association between the nitrogen-fixing rhizobia and leguminous plants [
24].
The strain used for co-culturing forms a distinct color, different from the nitrogen-fixing isolate, making the enumeration of each organism on the same plate easy. When the non-nitrogen-fixing strain was grown separately on a nitrogen-free medium, no colony was detected. However, upon coculturing with one of the nitrogen-fixing isolates of this study, the strain showed an abundant growth, indicating a supply of fixed nitrogen for the non-nitrogen-fixing strain. This, therefore, indirectly confirms that the alkaliphilic isolate from Lake Chitu expressed the nifH gene, and fixed enough atmospheric nitrogen, making reduced nitrogen available for the growth of the cocultured non-nitrogen-fixing organism. This probably indicates that nitrogen fixation by alkaliphilic microbial species may have an important contribution on the nitrogen economy of soda lakes.
The enzyme, nitrogenase, is highly sensitive to oxygen [
25]. Thus, nitrogen-fixing prokaryotes either grow anaerobically or develop mechanisms to protect their nitrogenase from exposure to oxygen through different mechanisms. Out of the nine-nitrogen-fixing alkaliphilic OTUs isolated from Lake Chitu, only one OTU was isolated from surface water, whereas eight isolates, accounting nearly 90%, were isolated from the anaerobic zone, below a depth of 2.9 m. Of these, three OTUs were isolated only from the sediment sample at a depth of 17 m. The distribution of the nitrogen-fixing OTUs to the anaerobic part of the lake indicate the adaptation of these organisms to anaerobic life, and this helps to protect the nitrogenase from inactivation by oxygen.
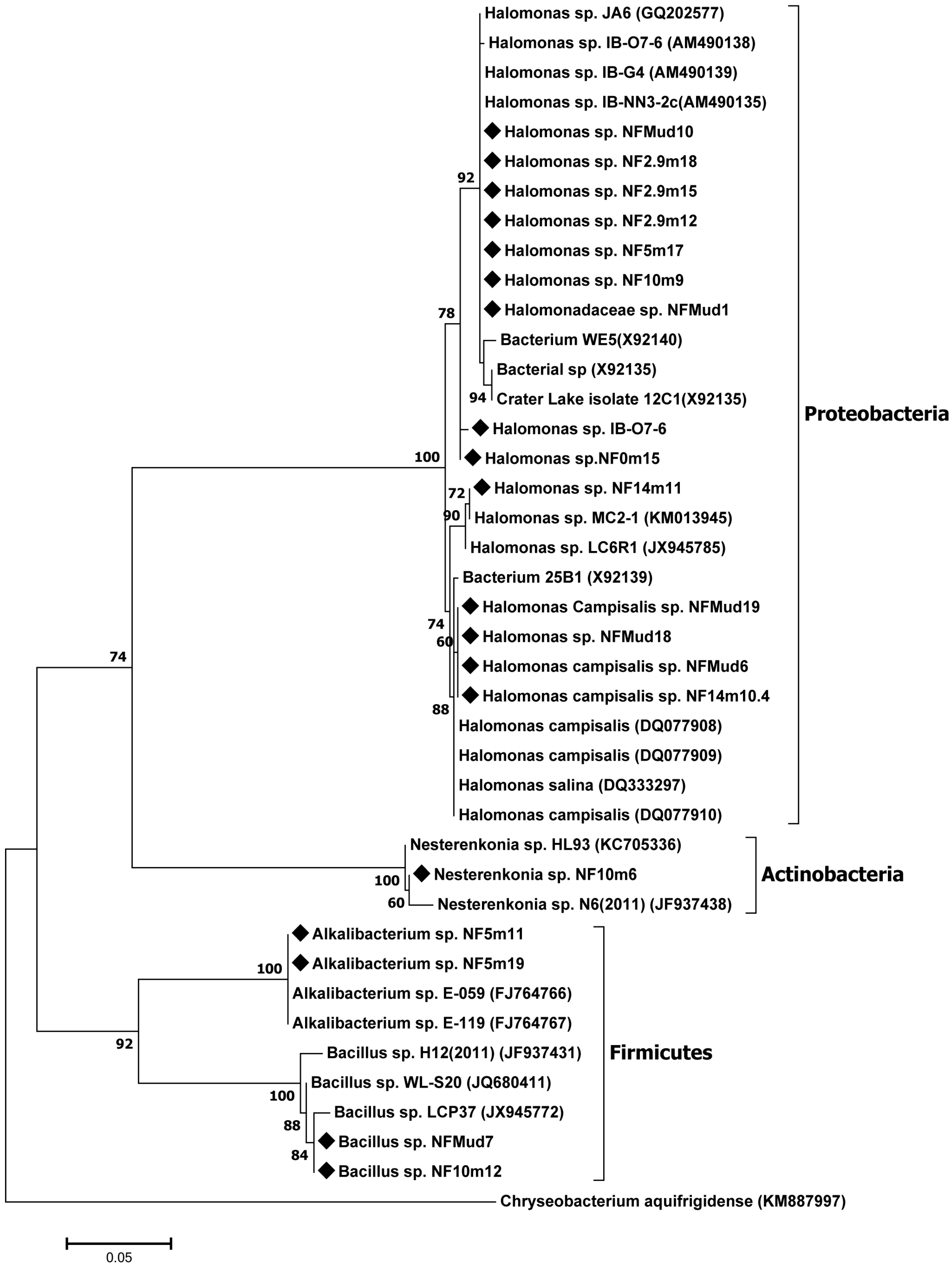
Most of the nitrogen-fixing isolates of the present study are closely related to bacterial strains isolated from various soda lakes in different parts of the world, with 99–100% 16S rRNA gene sequence similarity [
21,
26,
27,
28], where members of the genus,
Halomonas, were the most abundant. Previous studies reported that the genus,
Halomonas, was the most abundant culturable Gram-negative bacteria in soda lakes and other hypersaline environments [
28], able to grow on nutrient-poor culture media. Some members of the genus were also reported to be diazotrophs [
29].
The other group of isolates (represented by isolate NF10m6) that grow in nitrogen-free media and possess the nifH gene belong to the actinobacterial genus, Nesterenkonia. To the best of our knowledge, no member of the phylum, Actinobacteria, has been reported to fix nitrogen in soda lake habitats.
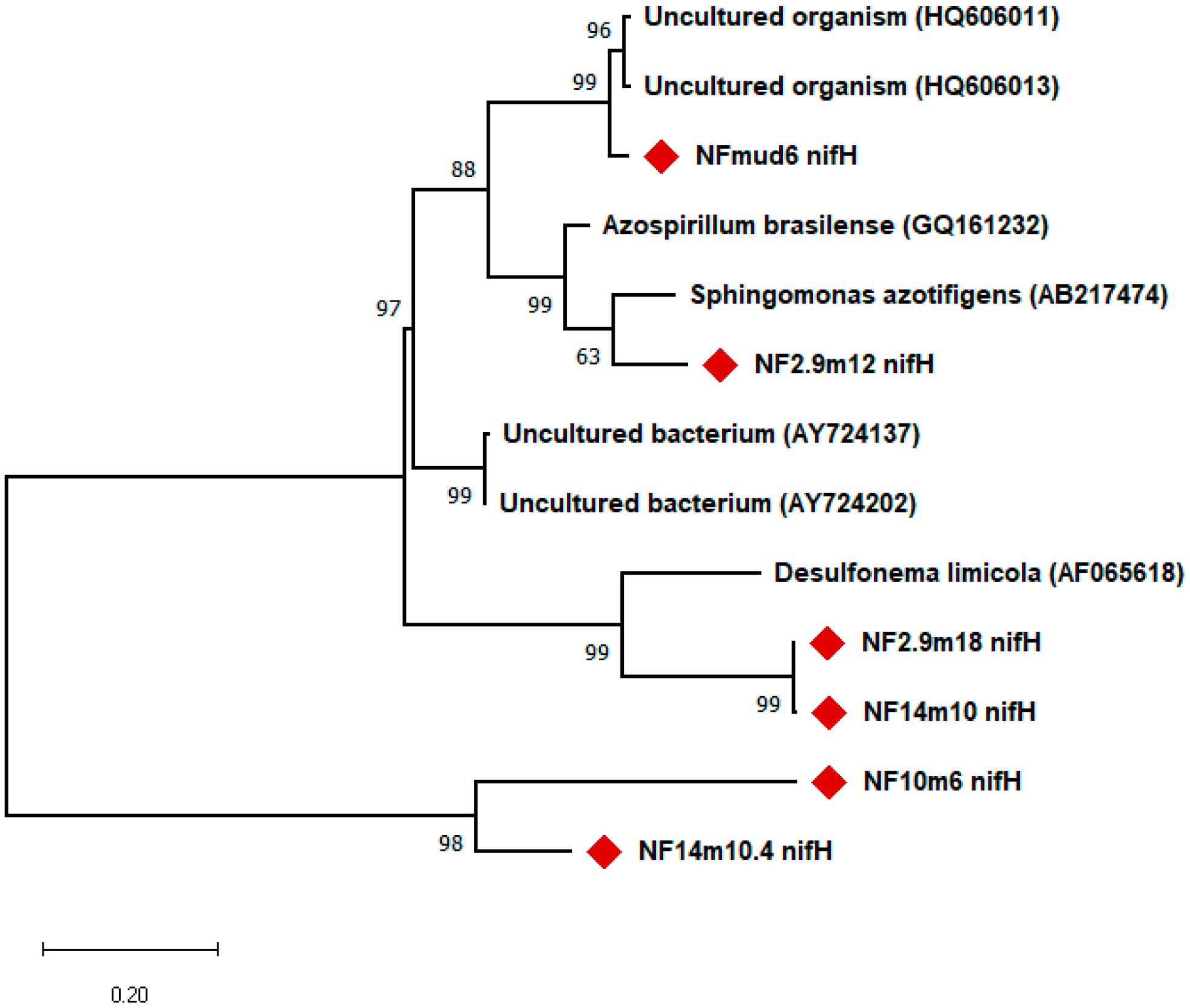
The
nifH gene fragment was amplified from 11 of the 19 isolates tested. Phylogenetic analysis based on
nifH gene fragment sequences showed that their closest matches were diazotrophic bacterial strains belonging to the phylum,
Proteobacteria, and uncultured microorganisms isolated from different environments. For example, the
nifH gene fragment sequenced from clone, Mud6, showed greater than 96% similarity to the
nifH gene sequence of an uncultured organism clone obtained from the Mediterranean Sea [
30]. The
nifH gene fragments of NF10m6, NF2.9m8, and NF14m10 showed 100%, with a putative nitrogenase reductase gene obtained from the uncultured microorganism clone, B10-31, obtained from a marine habitat [
31], whereas the
nifH gene fragment of isolate, 14m10, showed more than 92% sequence identity with the B21 dinitrogenase reductase (
nifH) gene of uncultured bacterium clone, DUN1_nifH_+, and uncultured bacterium clone, DUNnif131 (+B26) [
32].
The presence of a diverse group of nitrogen-fixing alkaliphiles in soda lakes may indicate the important role these organisms play as sources of nitrogen, supporting the high productivity of soda lakes. Spirulina is the main primary producer in Lake Chitu, a cyanobacterium with a wide range of industrial applications [
33]. At present, Spirulina is cultivated commercially using an artificial medium containing an inorganic nitrogen source, mostly in the form of nitrate [
34]. In its natural setting at Lake Chitu, Spirulina grows together with nitrogen-fixing prokaryote. This could indicate the interesting possibility of designing a mixed culture system consisting of nitrogen-fixing prokaryotes and the cyanobacteria. In addition to providing a cheap and sustainable way of producing food, animal feed, and other industrial products from the Spirulina biomass [
33], such a system could provide an efficient mechanism of fixing CO
2 released by different industries [
34].









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}