
Sweet Potato Symptomless Virus 1: First Detection in Europe and Generation of an Infectious Clone


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Samples
2.2. DNA Extraction, PCR Amplification, and Cloning
2.3. Sequence Analysis
2.4. Development of an Agroinfectious Clone and Plant Agroinoculation
3. Results
3.1. Sweet Potato Symptomless Virus 1 Naturally Infects Sweet Potato in Continental Spain and the Canary Islands
3.2. Sweet Potato Symptomless Virus 1 Is Present in Pathogen-Tested In Vitro Plants Obtained from a Germplasm Collection
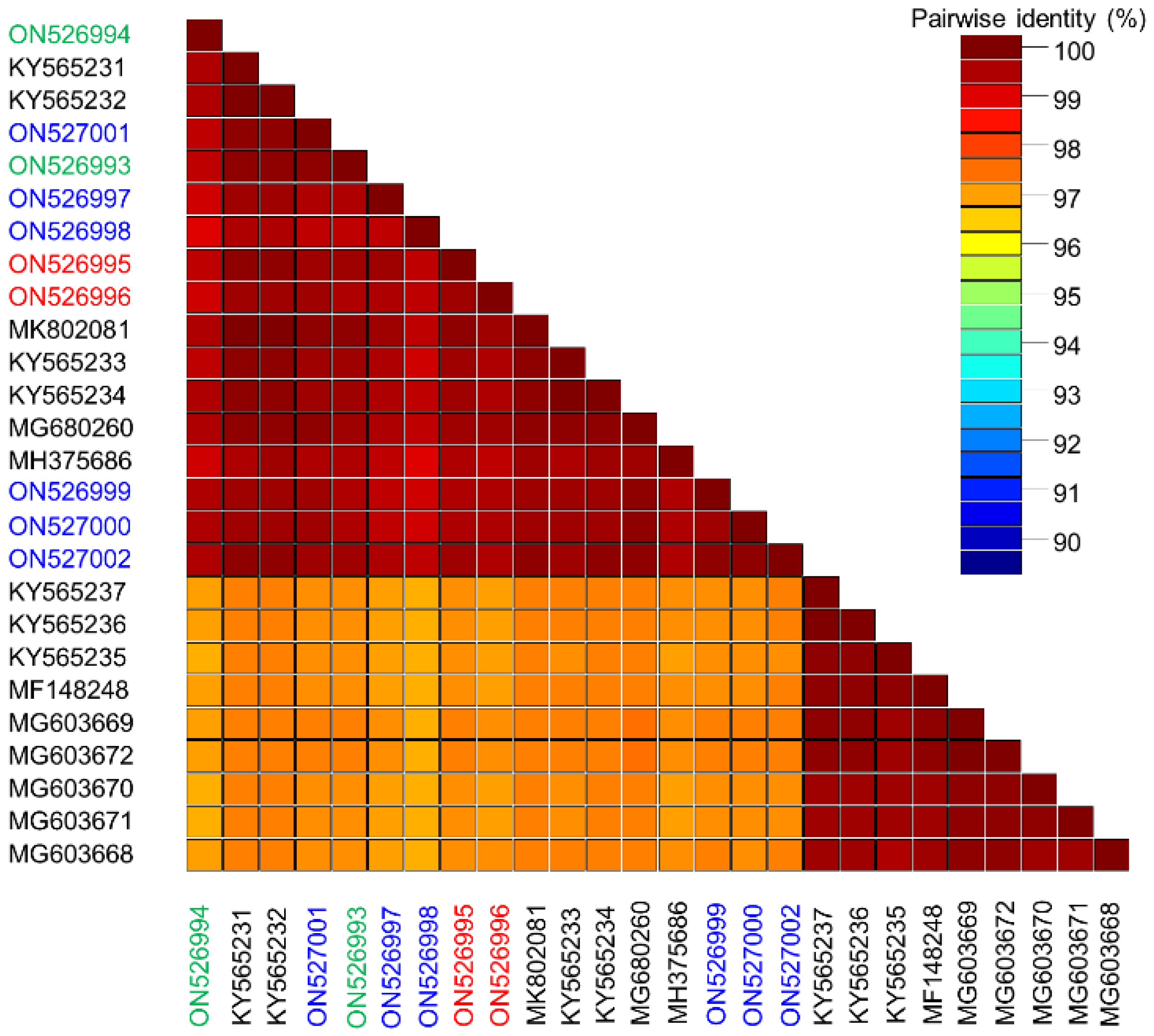
3.3. Sweet Potato Symptomless Virus 1 Isolates from the Canary Islands and Málaga Are Closely Related with Isolates from All over the World but Contain Novel Molecular Features
3.4. A Dimeric Sweet Potato Symptomless Virus 1 Clone Is Infectious in N. benthamiana, I. nil, I. setosa, and Sweet Potato
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valverde, R.A.; Sim, J.; Lotrakul, P. Whitefly transmission of sweet potato viruses. Virus Res. 2004, 100, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Valverde, R.A.; Lozano, G.; Navas-Castillo, J.; Ramos, A.; Valdés, F. First report of Sweet potato chlorotic stunt virus and Sweet potato feathery mottle virus infecting sweet potato in Spain. Plant Dis. 2004, 88, 428. [Google Scholar] [CrossRef]
- Trenado, H.P.; Lozano, G.; Valverde, R.A.; Navas-Castillo, J. First report of Sweet potato virus G and Sweet potato virus 2 infecting sweet potato in Spain. Plant Dis. 2007, 9, 1687. [Google Scholar] [CrossRef] [PubMed]
- Banks, G.K.; Bedford, I.D.; Beitia, F.J.; Rodriguez-Cerezo, E.; Markham, P.G. A novel geminivirus of Ipomoea indica (Convolvulacae) from Southern Spain. Plant Dis. 1999, 83, 486. [Google Scholar] [CrossRef]
- Lozano, G.; Trenado, H.P.; Valverde, R.A.; Navas-Castillo, J. Novel begomovirus species of recombinant nature in sweet potato (Ipomoea batatas) and Ipomoea indica: Taxonomic and phylogenetic implications. J. Gen. Virol. 2009, 90, 2550–2562. [Google Scholar] [CrossRef]
- Lozano, G.; Trenado, H.P.; Fiallo-Olivé, E.; Chirinos, D.; Geraud-Pouey, F.; Briddon, R.W.; Navas-Castillo, J. Characterization of non-coding DNA satellites associated with sweepoviruses (genus Begomovirus, Geminiviridae)—definition of a distinct class of begomovirus-associated satellites. Front. Microbiol. 2016, 7, 162. [Google Scholar] [CrossRef] [PubMed]
- Hassan, I.; Orílio, A.F.; Fiallo-Olivé, E.; Briddon, R.W.; Navas-Castillo, J. Infectivity, effects on helper viruses and whitefly transmission of the deltasatellites associated with sweepoviruses (genus Begomovirus, family Geminiviridae). Sci. Rep. 2016, 6, 30204. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Geminiviridae 2021. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef]
- Kreuze, J.F.; Perez, A.; Untiveros, M.; Quispe, D.; Fuentes, S.; Barker, I.; Simon, R. Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: A generic method for diagnosis, discovery and sequencing of viruses. Virology 2009, 388, 1–7. [Google Scholar] [CrossRef]
- Cao, M.; Lan, P.; Li, F.; Abad, J.; Zhou, C.; Li, R. Genome characterization of sweet potato symptomless virus 1: A mastrevirus with an unusual nonanucleotide sequence. Arch. Virol. 2017, 162, 2881–2884. [Google Scholar] [CrossRef]
- Kwak, H.-R.; Kim, M.-K.; Shin, J.-C.; Lee, Y.-J.; Seo, J.-K.; Lee, H.-U.; Jung, M.-N.; Kim, S.-H.; Choi, H.-S. The current incidence of viral disease in Korean sweet potatoes and development of multiplex RT-PCR assays for simultaneous detection of eight sweet potato viruses. Plant Pathol. J. 2014, 30, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Mbanzibwa, D.R.; Tugume, A.K.; Chiunga, E.; Mark, D.; Tairo, F.D. Small RNA deep sequencing-based detection and further evidence of DNA viruses infecting sweetpotato plants in Tanzania. Ann. Appl. Biol. 2014, 165, 329–339. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Zhang, D.-S.; Zhang, Z.-C.; Wang, S.; Qiao, Q.; Qin, Y.-H.; Tian, Y.-T. First report on sweetpotato symptomless virus 1 (genus Mastrevirus, family Geminiviridae) in sweetpotato in China. Plant Dis. 2015, 99, 1042. [Google Scholar] [CrossRef]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Muhire, B.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Ferreira, P.T.; Lemos, T.O.; Nagata, T.; Inoue-Nagata, A.K. One-step cloning approach for construction of agroinfectious begomovirus clones. J. Virol. Methods 2008, 147, 351–354. [Google Scholar] [CrossRef]
- Ferro, C.G.; Zerbini, F.M.; Navas-Castillo, J.; Fiallo-Olivé, E. Revealing the complexity of sweepovirus-deltasatellite-plant host interactions: Expanded natural and experimental helper virus range and effect dependence on virus-host combination. Microorganisms 2021, 9, 1018. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Tovar, R.; Navas-Castillo, J. Deciphering the biology of deltasatellites from the New World: Maintenance by New World begomoviruses and whitefly transmission. New Phytol. 2016, 212, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Untiveros, M.; Fuentes, S.; Salazar, L.F. Synergistic interaction of Sweet potato chlorotic stunt virus (Crinivirus) with carla-, cucumo-, ipomo-, and potyviruses infecting sweet potato. Plant Dis. 2007, 91, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Cuellar, W.J.; Galvez, M.; Fuentes, S.; Tugume, J.; Kreuze, J. Synergistic interactions of begomoviruses with Sweet potato chlorotic stunt virus (genus Crinivirus) in sweet potato (Ipomoea batatas L.). Mol. Plant Pathol. 2015, 16, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Perez, A.; Gargurevich, M.G.; Cuellar, W.J. Badnaviruses of sweet potato: Symptomless coinhabitants on a global scale. Front. Plant Sci. 2020, 11, 313. [Google Scholar] [CrossRef]





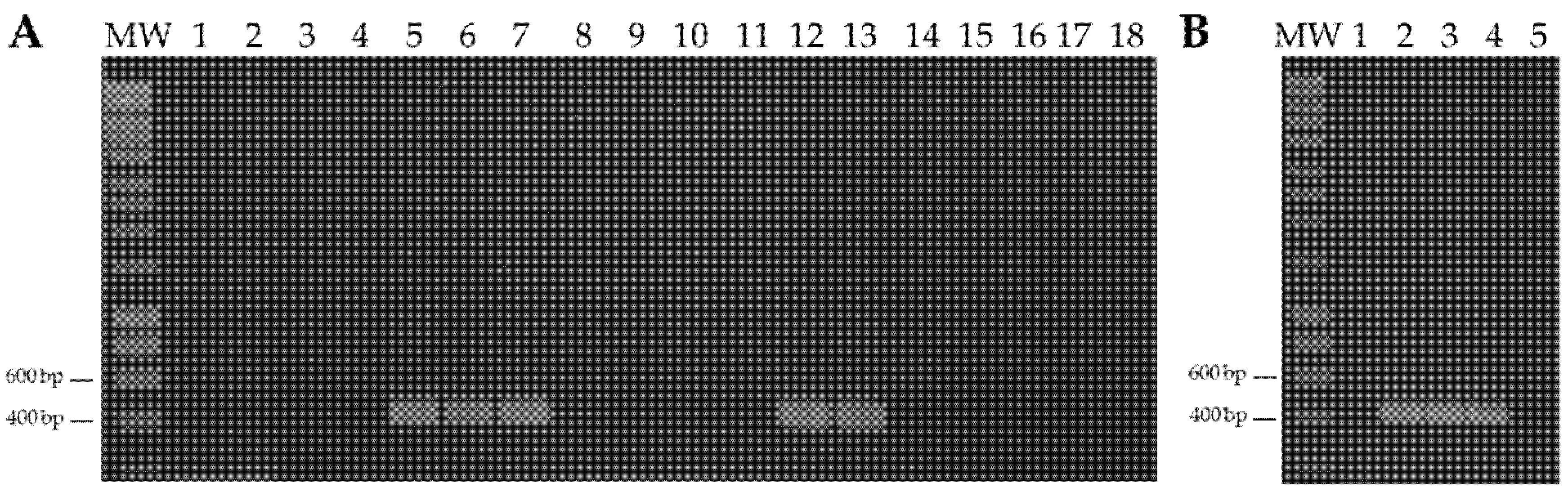{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | No. of Infected Sweet Potato Plants/Total No. of Sweet Potato Plants (Percentage) | No. of Infected I. indica Plants/Total No. of I. indica Plants |
|---|---|---|
| Tenerife (Canary Islands) | 16/34 (47.05%) | 0/12 |
| Gran Canaria (Canary Islands) | 14/22 (63.63%) | 0/12 |
| Málaga (southern continental Spain) | 6/15 (40%) | - |
| Total | 36/71 (50.70%) | 0/12 |
| No. of Infected Plants/No. of Agroinoculated Plants | |||
|---|---|---|---|
| Plant Species | Experiment 1 | Experiment 2 | Total (%) |
| Nicotiana benthamiana | 9/12 0/12 (mock) | 2/12 0/12 (mock) | 45.83 |
| Ipomoea nil | 3/24 0/12 (mock) | 3/24 0/12 (mock) | 12.5 |
| Ipomoea setosa | 6/24 0/12 (mock) | 9/24 0/12 (mock) | 31.25 |
| Sweet potato cv. ‘Tanzania’ | 2/18 0/5 (mock) | 4/10 0/3 (mock) | 21.42 |
| Sweet potato cv. ‘Camote Morado’ | 2/13 0/5 (mock) | 2/14 0/3 (mock) | 14.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiallo-Olivé, E.; García-Merenciano, A.C.; Navas-Castillo, J. Sweet Potato Symptomless Virus 1: First Detection in Europe and Generation of an Infectious Clone. Microorganisms 2022, 10, 1736. https://doi.org/10.3390/microorganisms10091736
Fiallo-Olivé E, García-Merenciano AC, Navas-Castillo J. Sweet Potato Symptomless Virus 1: First Detection in Europe and Generation of an Infectious Clone. Microorganisms. 2022; 10(9):1736. https://doi.org/10.3390/microorganisms10091736
Chicago/Turabian StyleFiallo-Olivé, Elvira, Ana Cristina García-Merenciano, and Jesús Navas-Castillo. 2022. "Sweet Potato Symptomless Virus 1: First Detection in Europe and Generation of an Infectious Clone" Microorganisms 10, no. 9: 1736. https://doi.org/10.3390/microorganisms10091736