
Mixed Fungal Biofilms: From Mycobiota to Devices, a New Challenge on Clinical Practice

Abstract
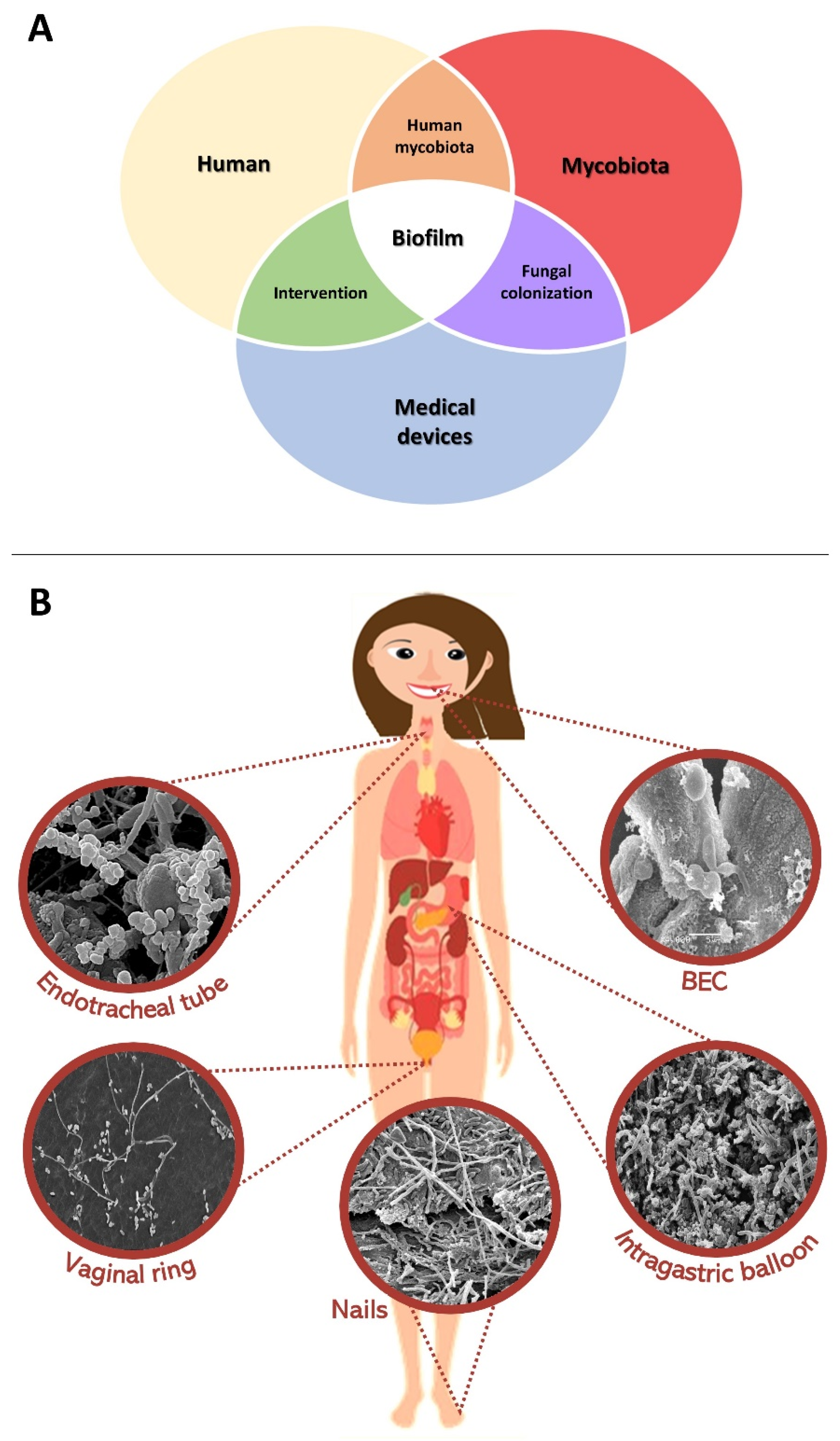
:1. Introduction
2. Microbiota of Human Anatomical Niches
3. What Are Mixed Fungal Biofilms and Where Are They Naturally Found?
4. Individual Fungal Attributes Relevant to Interaction and Communication in Mixed Fungal–Fungal Biofilms
5. Conclusions and Perspectives
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clancy, C.J.; Nguyen, M.H. Diagnosing Invasive Candidiasis. J. Clin. Microbiol. 2018, 56, e01909-17. [Google Scholar] [CrossRef]
- Ibáñez-Martínez, E.; Ruiz-Gaitán, A.; Pemán-García, J. Update on the diagnosis of invasive fungal infection. Rev. Esp. Quimioter. 2017, 30 (Suppl. S1), 16–21. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28882009 (accessed on 30 July 2022).
- Patterson, T.F.; Thompson, G.R., III; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef]
- Krüger, W.; Vielreicher, S.; Kapitan, M.; Jacobsen, I.D.; Niemiec, M.J. Fungal-Bacterial Interactions in Health and Disease. Pathogens 2019, 8, 70. [Google Scholar] [CrossRef]
- Singh, V.; Prasad, A.; Panda, P.K.; Totaganti, M.; Tyagi, A.K.; Thaduri, A.; Rao, S.; Bairwa, M.; Singh, A.K. Mixed invasive fungal infections among COVID-19 patients. Curr. Med. Mycol. 2021, 7, 19–27. [Google Scholar] [CrossRef]
- Gupta, A.K.; Taborda, V.B.A.; Taborda, P.R.O.; Shemer, A.; Summerbell, R.C.; Nakrieko, K.A. High prevalence of mixed infections in global onychomycosis. PLoS ONE 2020, 15, e0239648. [Google Scholar] [CrossRef]
- Gupta, A.K.; Nakrieko, K.A. Trichophyton rubrum DNA Strains in Patients with Onychomycosis with Persistent Mixed Infections Involving a Nondermatophyte Mold. J. Am. Podiatr. Med. Assoc. 2020, 110. [Google Scholar] [CrossRef]
- Gupta, A.K.; Nakrieko, K.A. Molecular determination of mixed infections of dermatophytes and nondermatophyte molds in individuals with onychomycosis. J. Am. Podiatr. Med. Assoc. 2014, 104, 330–336. [Google Scholar] [CrossRef]
- Tiew, P.Y.; Mac Aogain, M.; Ali, N.A.B.M.; Thng, K.X.; Goh, K.; Lau, K.J.X.; Chotirmall, S.H. The Mycobiome in Health and Disease: Emerging Concepts, Methodologies and Challenges. Mycopathologia 2020, 185, 207–231. [Google Scholar] [CrossRef]
- Thakur, R.; Goyal, R. Tinea capitis: Mixed or Consecutive Infection with White and Violet Strains of Trichophyton violaceum: A Diagnostic or Therapeutic Challenge. J. Clin. Diagn. Res. 2015, 9, WD03–WD04. [Google Scholar] [CrossRef]
- Huffnagle, G.B.; Noverr, M.C. The emerging world of the fungal microbiome. Trends Microbiol. 2013, 21, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Yao, Y.F.; Li, J.Y.; Li, Y. Human mycobiome and diseases. Hua Xi Kou Qiang Yi Xue Za Zhi 2019, 37, 314–319. [Google Scholar] [CrossRef]
- Ackerman, A.L.; Underhill, D.M. The mycobiome of the human urinary tract: Potential roles for fungi in urology. Ann. Transl. Med. 2017, 5, 31. [Google Scholar] [CrossRef]
- Cui, L.; Morris, A.; Ghedin, E. The human mycobiome in health and disease. Genome Med. 2013, 5, 63. [Google Scholar] [CrossRef]
- Delaney, C.; Kean, R.; Short, B.; Tumelty, M.; McLean, W.; Nile, C.J.; Ramage, G. Fungi at the scene of the crime: Innocent bystanders or accomplices in oral infections? Curr. Clin. Microbiol. Rep. 2018, 5, 190–200. [Google Scholar] [CrossRef]
- Jo, J.H.; Kennedy, E.A.; Kong, H.H. Topographical and physiological differences of the skin mycobiome in health and disease. Virulence 2017, 8, 324–333. [Google Scholar] [CrossRef]
- Prigitano, A.; Perrone, P.M.; Esposto, M.C.; Carnevali, D.; De Nard, F.; Grimoldi, L.; Principi, N.; Cogliati, M.; Castaldi, S.; Romanò, L. ICU environmental surfaces are a reservoir of fungi: Species distribution in northern Italy. J. Hosp. Infect. 2022, 123, 74–79. [Google Scholar] [CrossRef]
- da Silva, E.M.; Mansano, E.S.B.; Bonfim-Mendonça, P.D.S.; Olegário, R.; Tobaldini-Valério, F.; Fiorini, A.; Svidzinski, T.I. High colonization by Candida parapsilosis sensu stricto on hands and surfaces in an adult intensive care unit. J. Mycol. Med. 2021, 31, 101110. [Google Scholar] [CrossRef]
- Barantsevich, N.; Barantsevich, E. Diagnosis and Treatment of Invasive Candidiasis. Antibiotics 2022, 11, 718. [Google Scholar] [CrossRef]
- Adamczyk, K.; Garncarczyk, A.; Antończak, P.; Wcisło-Dziadecka, D. The foot microbiome. J. Cosmet. Dermatol. 2020, 19, 1039–1043. [Google Scholar] [CrossRef]
- Tipton, L.; Ghedin, E.; Morris, A. The lung mycobiome in the next-generation sequencing era. Virulence 2017, 8, 334–341. [Google Scholar] [CrossRef]
- Pathak, J.L.; Yan, Y.; Zhang, Q.; Wang, L.; Ge, L. The role of oral microbiome in respiratory health and diseases. Respir. Med. 2021, 185, 106475. [Google Scholar] [CrossRef]
- Azzam, S.Z.; Cayme, G.J.; Martinez, L.R. Polymicrobial interactions involving fungi and their importance for the environment and in human disease. Microb. Pathog. 2020, 140, 103942. [Google Scholar] [CrossRef]
- Carpagnano, G.E.; Susca, A.; Scioscia, G.; Lacedonia, D.; Cotugno, G.; Soccio, P.; Santamaria, S.; Resta, O.; Logrieco, G.; Barbaro, M.P.F. A survey of fungal microbiota in airways of healthy volunteer subjects from Puglia (Apulia), Italy. BMC Infect. Dis. 2019, 19, 78. [Google Scholar] [CrossRef]
- Huang, C.; Yu, Y.; Du, W.; Liu, Y.; Dai, R.; Tang, W.; Wang, P.; Zhang, C.; Shi, G. Fungal and bacterial microbiome dysbiosis and imbalance of trans-kingdom network in asthma. Clin. Transl. Allergy 2020, 10, 42. [Google Scholar] [CrossRef]
- Dickson, R.P.; Erb-Downward, J.R.; Martinez, F.J.; Huffnagle, G.B. The Microbiome and the Respiratory Tract. Annu. Rev. Physiol. 2016, 78, 481–504. [Google Scholar] [CrossRef]
- Bandara, H.M.H.N.; Panduwawala, C.P.; Samaranayake, L.P. Biodiversity of the human oral mycobiome in health and disease. Oral Dis. 2019, 25, 363–371. [Google Scholar] [CrossRef]
- Baraniya, D.; Chen, T.; Nahar, A.; Alakwaa, F.; Hill, J.; Tellez, M.; Ismail, A.; Puri, S.; Al-Hebshi, N.N. Supragingival mycobiome and inter-kingdom interactions in dental caries. J. Oral Microbiol. 2020, 12, 1729305. [Google Scholar] [CrossRef]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Kong, H.H.; et al. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef]
- Imabayashi, Y.; Moriyama, M.; Takeshita, T.; Ieda, S.; Hayashida, J.-N.; Tanaka, A.; Maehara, T.; Furukawa, S.; Ohta, M.; Kubota, K.; et al. Molecular analysis of fungal populations in patients with oral candidiasis using next-generation sequencing. Sci. Rep. 2016, 6, 28110. [Google Scholar] [CrossRef]
- Monteiro-da-Silva, F.; Araujo, R.; Sampaio-Maia, B. Interindividual variability and intraindividual stability of oral fungal microbiota over time. Med. Mycol. 2014, 52, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Shelburne, S.A.; Ajami, N.J.; Chibucos, M.; Beird, H.; Tarrand, J.J.; Galloway-Pena, J.; Albert, N.D.; Chemaly, R.F.; Ghantoji, S.S.; Marsh, L.; et al. Implementation of a Pan-Genomic Approach to Investigate Holobiont-Infecting Microbe Interaction: A Case Report of a Leukemic Patient with Invasive Mucormycosis. PLoS ONE 2015, 10, e0139851. [Google Scholar] [CrossRef]
- Schwartz, S.; Thiel, E. Images in clinical medicine. Palate destruction by Aspergillus. N. Engl. J. Med. 1997, 337, 241. [Google Scholar] [CrossRef]
- Iatta, R.; Napoli, C.; Borghi, E.; Montagna, M.T. Rare mycoses of the oral cavity: A literature epidemiologic review. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2009, 108, 647–655. [Google Scholar] [CrossRef]
- Pasqualotto, A.C.; Antunes, A.G.V.; Severo, L.C. Candida guilliermondii as the aetiology of candidosis. Rev. Inst. Med. Trop. Sao Paulo 2006, 48, 123–127. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Lu, X.L.; Mounmin, F.A. Diagnosis and Treatment of Esophageal Candidiasis: Current Updates. Can. J. Gastroenterol. Hepatol. 2019, 2019, 3585136. [Google Scholar] [CrossRef]
- Robertson, K.D.; Nagra, N.; Mehta, D. Esophageal Candidiasis. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30725953 (accessed on 25 June 2022).
- Balish, E. A URA3 null mutant of Candida albicans (CAI-4) causes oro-oesophageal and gastric candidiasis and is lethal for gnotobiotic, transgenic mice (Tgepsilon26) that are deficient in both natural killer and T cells. J. Med. Microbiol. 2009, 58 Pt 3, 290–295. [Google Scholar] [CrossRef]
- Hoeflinger, J.L.; Coleman, D.A.; Oh, S.H.; Miller, M.J.; Hoyer, L.L. A piglet model for studying Candida albicans colonization of the human oro-gastrointestinal tract. FEMS Microbiol. Lett. 2014, 357, 10–15. [Google Scholar] [CrossRef]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; Brown, J.; Becker, C.A.; Fleshner, P.R.; Dubinsky, M.; et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012, 336, 1314–1317. [Google Scholar] [CrossRef]
- Huseyin, C.E.; O’Toole, P.W.; Cotter, P.D.; Scanlan, P.D. Forgotten fungi-the gut mycobiome in human health and disease. FEMS Microbiol. Rev. 2017, 41, 479–511. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Shoemaker, R.; Kim, J. Urobiome: An outlook on the metagenome of urological diseases. Investig. Clin. Urol. 2021, 62, 611–622. [Google Scholar] [CrossRef]
- Rodríguez-Cerdeira, C.; Martínez-Herrera, E.; Carnero-Gregorio, M.; López-Barcenas, A.; Fabbrocini, G.; Fida, M.; El-Samahy, M.; González-Cespón, J.L. Pathogenesis and Clinical Relevance of Biofilms in Vulvovaginal Candidiasis. Front. Microbiol. 2020, 11, 544480. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef]
- Prado, A.; Brito, R.O.; Pereira, E.C.A.; Correa, J.L.; Neto, M.G.; Dayyeh, B.K.A.; Negri, M.; Svidzinski, T.I.E. First Study of Naturally Formed Fungal Biofilms on the Surface of Intragastric Balloons. Obes. Surg. 2021, 31, 5348–5357. [Google Scholar] [CrossRef]
- Pereira, R.; Dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida albicans: Formation, regulation and resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef]
- van Tilburg Bernardes, E.; Gutierrez, M.W.; Arrieta, M.C. The Fungal Microbiome and Asthma. Front. Cell Infect. Microbiol. 2020, 10, 583418. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef]
- Kumar, J.; Sharma, V.K.; Parmar, S.; Singh, P.; Singh, R.K. Biofilm: A microbial assemblage on the surface—A boon or bane? In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Biofilms; Elsevier: Amsterdam, The Netherlands, 2020; pp. 139–150. [Google Scholar] [CrossRef]
- Lagree, K.; Mitchell, A.P. Fungal Biofilms: Inside Out. Microbiol. Spectr. 2017, 5, 5-2. [Google Scholar] [CrossRef]
- Nett, J.E.; Pohl, C.H. Editorial: Fungal Biofilms in Infection and Disease. Front. Cell Infect. Microbiol. 2021, 11, 753650. [Google Scholar] [CrossRef]
- Eix, E.F.; Nett, J.E. How Biofilm Growth Affects -Host Interactions. Front. Microbiol. 2020, 11, 1437. [Google Scholar] [CrossRef]
- Oliveira, L.T.; Medina-Alarcón, K.P.; Singulani, J.D.L.; Fregonezi, N.F.; Pires, R.H.; Arthur, R.A.; Fusco-Almeida, A.M.; Giannini, M.J.S.M. Dynamics of Mono- and Dual-Species Biofilm Formation and Interactions Between and. Front. Microbiol. 2020, 11, 551256. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; De Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarcón, K.P.; Melo, W.C.M.A.; Marcelino, M.Y.; Braz, J.D.; et al. Fungal Biofilms and Polymicrobial Diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef]
- Jarros, I.C.; Barros, I.L.E.; Prado, A.; Corrêa, J.L.; Malacrida, A.M.; Negri, M.; Svidzinski, T.I.E. Rhodotorula sp. And Trichosporon sp. Are more Virulent After a Mixed Biofilm. Mycopathologia 2022, 187, 85–93. [Google Scholar] [CrossRef]
- Ponde, N.O.; Lortal, L.; Ramage, G.; Naglik, J.R.; Richardson, J.P. biofilms and polymicrobial interactions. Crit. Rev. Microbiol. 2021, 47, 91–111. [Google Scholar] [CrossRef]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef]
- Gawaz, A.; Weisel, G. Mixed infections are a critical factor in the treatment of superficial mycoses. Mycoses 2018, 61, 731–735. [Google Scholar] [CrossRef]
- Gupta, A.K.; Renaud, H.J.; Quinlan, E.M.; Shear, N.H.; Piguet, V. The Growing Problem of Antifungal Resistance in Onychomycosis and Other Superficial Mycoses. Am. J. Clin. Dermatol. 2021, 22, 149–157. [Google Scholar] [CrossRef]
- Bunyaratavej, S.; Leeyaphan, C.; Rujitharanawong, C.; Muanprasat, C.; Matthapan, L. Clinical and Laboratory Characteristics of a Tinea capitis Outbreak Among Novice Buddhist Monks. Pediatr. Dermatol. 2017, 34, 371–373. [Google Scholar] [CrossRef]
- Wiegand, C.; Mugisha, P.; Mulyowa, G.K.; Elsner, P.; Hipler, U.C.; Gräser, Y.; Uhrlaß, S.; Nenoff, P. Trichophyton violaceum: Main cause of Tinea capitis in children at Mbarara Regional Referral Hospital in Uganda. Hautarzt 2016, 67, 712–717. [Google Scholar] [CrossRef]
- Yang, X.; Shi, X.; Chen, W.; Zhou, Y.; Lionakis, M.S.; Kontoyiannis, D.P.; Liu, W. First report of kerion (Tinea capitis) caused by combined and. Med. Mycol. Case Rep. 2020, 29, 5–7. [Google Scholar] [CrossRef]
- De Costa, P.S.; Mendes, V.; Veiga, F.F.; Negri, M.; Svidzinski, T.I.E. Relevant insights into onychomycosis’ pathogenesis related to the effectiveness topical treatment. Microb. Pathog. 2022, 169, 105640. [Google Scholar] [CrossRef]
- Gupta, A.K.; Versteeg, S.G.; Shear, N.H. Onychomycosis in the 21st Century: An Update on Diagnosis, Epidemiology, and Treatment. J. Cutan. Med. Surg. 2017, 21, 525–539. [Google Scholar] [CrossRef]
- Gupta, A.K.; Daigle, D.; Carviel, J.L. The role of biofilms in onychomycosis. J. Am. Acad. Dermatol. 2016, 74, 1241–1246. [Google Scholar] [CrossRef]
- Salakshna, N.; Bunyaratavej, S.; Matthapan, L.; Lertrujiwanit, K.; Leeyaphan, C. A cohort study of risk factors, clinical presentations, and outcomes for dermatophyte, nondermatophyte, and mixed toenail infections. J. Am. Acad. Dermatol. 2018, 79, 1145–1146. [Google Scholar] [CrossRef]
- Idris, N.F.B.; Huang, G.; Jia, Q.; Yuan, L.; Li, Y.; Tu, Z. Mixed Infection of Toe Nail Caused by Trichosporon asahii and Rhodotorula mucilaginosa. Mycopathologia 2020, 185, 373–376. [Google Scholar] [CrossRef]
- Roudbary, M.; Vahedi-Shahandashti, R.; dos Santos, A.L.S.; Mohammadi, S.R.; Aslani, P.; Lass-Flörl, C.; Rodrigues, C.F. Biofilm formation in clinically relevant filamentous fungi: A therapeutic challenge. Crit. Rev. Microbiol. 2022, 48, 197–221. [Google Scholar] [CrossRef]
- Ferreira, E.G.; Yatsuda, F.; Pini, M.; Jarros, I.C.; Veiga, F.F.; De Oliveira, A.G.; Negri, M.; Svidzinski, T.I.E. Implications of the presence of yeasts in tracheobronchial secretions of critically ill intubated patients. EXCLI J. 2019, 18, 801–811. [Google Scholar] [CrossRef]
- Obručová, H.; Kotásková, I.; Tihelková, R.; Holá, V.; Růžička, F.; Freiberger, T. Fluorescent Capillary Electrophoresis Is Superior to Culture in Detecting Species from Samples of Urinary Catheters and Ureteral Stents with Mono-or Polyfungal Biofilm Growth. J. Clin. Microbiol. 2019, 57, e01861-18. [Google Scholar] [CrossRef]
- Bekkal Brikci Benhabib, O.; Boucherit-Otmani, Z.; Boucherit, K.; Djediat, C. Interaction in a dual-species biofilm of Candida albicans and Candida glabrata co-isolated from intravascular catheter. Microb. Pathog. 2021, 152, 104613. [Google Scholar] [CrossRef]
- Bayrak, O.; Basmaci, I.; Zer, Y.; Karabulut, E.K.; Sen, H.; Erturhan, S.; Seckiner, I. Colonizations on biofilm layers of D-J catheters under sterile urine conditions. Arch. Esp. Urol. 2019, 72, 75–79. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30741656 (accessed on 30 June 2022).
- Duarte, H.; Santos, C.; Capelas, M.L.; Fonseca, J. Peristomal infection after percutaneous endoscopic gastrostomy: A 7-year surveillance of 297 patients. Arq. Gastroenterol. 2012, 49, 255–258. [Google Scholar] [CrossRef]
- Lucendo, A.J.; Friginal-Ruiz, A.B. Percutaneous endoscopic gastrostomy: An update on its indications, management, complications, and care. Rev. Esp. Enferm. Dig. 2014, 106, 529–539. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25544410 (accessed on 30 June 2022).
- Martins, C.H.G.; Pires, R.H.; Cunha, A.O.; Pereira, C.A.M.; Singulani, J.D.L.; Abrão, F.; de Moraes, T.; Mendes-Giannini, M.J.S. Candida/Candida biofilms. First description of dual-species Candida albicans/C. rugosa biofilm. Fungal Biol. 2016, 120, 530–537. [Google Scholar] [CrossRef]
- Shao, J.; Lu, K.; Tian, G.; Cui, Y.; Yan, Y.; Wang, T.; Zhang, X.; Wang, C. Lab-scale preparations of Candida albicans and dual Candida albicans-Candida glabrata biofilms on the surface of medical-grade polyvinyl chloride (PVC) perfusion tube using a modified gravity-supported free-flow biofilm incubator (GS-FFBI). J. Microbiol. Methods 2015, 109, 41–48. [Google Scholar] [CrossRef]
- Thorarinsdottir, H.R.; Kander, T.; Holmberg, A.; Petronis, S.; Klarin, B. Biofilm formation on three different endotracheal tubes: A prospective clinical trial. Crit. Care 2020, 24, 382. [Google Scholar] [CrossRef]
- Meslemani, D.; Yaremchuk, K.; Rontal, M. Presence of biofilm on adult tracheostomy tubes. Ear Nose Throat J. 2010, 89, 496–504. Available online: https://www.ncbi.nlm.nih.gov/pubmed/20981663 (accessed on 2 July 2022).
- Zhang, L.; Yang, L.; Dong, W.; Liu, X.; Lei, X.; Zhang, L. Risk factors and clinical analysis of peripherally inserted central catheter-related fungal colonization in premature infants. Sci. Rep. 2021, 11, 20897. [Google Scholar] [CrossRef]
- Seghir, A.; Boucherit-Otmani, Z.; Belkherroubi-Sari, L.; Boucherit, K. Catheterization and fungal infection risk in the University Hospital of Tlemcen: Epidemiology and susceptibility to antifungals. J. Mycol. Med. 2014, 24, e179–e184. [Google Scholar] [CrossRef]
- Gross, C.E.; Della Valle, C.J.; Rex, J.C.; Traven, S.A.; Durante, E.C. Fungal Periprosthetic Joint Infection: A Review of Demographics and Management. J. Arthroplast. 2021, 36, 1758–1764. [Google Scholar] [CrossRef]
- Karczewski, D.; Khakzad, T.; Kriechling, P.; Akgün, D. Aspergillus PJI-A systematic analysis of all known cases and report of a new one. J. Mycol. Med. 2021, 31, 101141. [Google Scholar] [CrossRef]
- Gao, Z.; Li, X.; Du, Y.; Peng, Y.; Wu, W.; Zhou, Y. Success Rate of Fungal Peri-Prosthetic Joint Infection Treated by 2-Stage Revision and Potential Risk Factors of Treatment Failure: A Retrospective Study. Med. Sci. Monit. 2018, 24, 5549–5557. [Google Scholar] [CrossRef]
- Konne, N.M.; Collier, S.A.; Spangler, J.; Cope, J.R. Healthy Contact Lens Behaviors Communicated by Eye Care Providers and Recalled by Patients-United States, 2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 693–697. [Google Scholar] [CrossRef]
- Efron, N.; Morgan, P.B.; Woods, C.A.; International Contact Lens Prescribing Survey Consortium. Survey of contact lens prescribing to infants, children, and teenagers. Optom. Vis. Sci. 2011, 88, 461–468. [Google Scholar] [CrossRef]
- Thomas, P.A.; Kaliamurthy, J. Mycotic keratitis: Epidemiology, diagnosis and management. Clin. Microbiol. Infect. 2013, 19, 210–220. [Google Scholar] [CrossRef]
- De Andrade, A.J.M.; Vieira, L.A.; Höfling-Lima, A.L.; Yu, M.C.Z.; Gompertz, O.F.; De Freitas, D.; De Souza, L.B. Laboratorial analyses of fungal keratitis in a University Service. Arq. Bras. Oftalmol. 2000, 63, 59–63. [Google Scholar] [CrossRef]
- Cheung, N.N.; Cheng, Y.Y.Y.; van Duinen, S.G.; Houbraken, J.; Verweij, P.E.; Gooskens, J. Contact lens-related fungal keratitis. Lancet Infect. Dis. 2020, 20, 1100. [Google Scholar] [CrossRef]
- Watson, S.L.; Cabrera-Aguas, M.; Keay, L.; Khoo, P.; McCall, D.; Lahra, M.M. The clinical and microbiological features and outcomes of fungal keratitis over 9 years in Sydney, Australia. Mycoses 2020, 63, 43–51. [Google Scholar] [CrossRef]
- Tendo, C.; Shahin, B.; Welch, J. Unusual finding of two filamentous fungi in a contact lens-related keratitis. BMJ Case Rep. 2021, 14, e239085. [Google Scholar] [CrossRef]
- Mousa, M.A.; Lynch, E.; Kielbassa, A.M. Denture-related stomatitis in new complete denture wearers and its association with Candida species colonization: A prospective case-series. Quintessence Int. 2020, 51, 554–565. [Google Scholar] [CrossRef]
- Meirowitz, A.; Rahmanov, A.; Shlomo, E.; Zelikman, H.; Dolev, E.; Sterer, N. Effect of Denture Base Fabrication Technique on Adhesion In vitro. Materials 2021, 14, 221. [Google Scholar] [CrossRef]
- Stober, T.; Bermejo, J.L.; Rues, S.; Rammelsberg, P. Wear of resin denture teeth in partial removable dental prostheses. J. Prosthodont. Res. 2020, 64, 85–89. [Google Scholar] [CrossRef]
- Baba, N.Z.; Goodacre, B.J.; Goodacre, C.J.; Müller, F.; Wagner, S. CAD/CAM Complete Denture Systems and Physical Properties: A Review of the Literature. J. Prosthodont. 2021, 30, 113–124. [Google Scholar] [CrossRef]
- Kim, J.J. Revisiting the Removable Partial Denture. Dent. Clin. N. Am. 2019, 63, 263–278. [Google Scholar] [CrossRef]
- Li, J.; Hirota, K.; Goto, T.; Yumoto, H.; Miyake, Y.; Ichikawa, T. Biofilm formation of Candida albicans on implant overdenture materials and its removal. J. Dent. 2012, 40, 686–692. [Google Scholar] [CrossRef]
- Gupta, V.; Abhisheik, K.; Balasundari, S.; Devendra, N.K.; Shadab, K.; Anupama, M. Identification of using different culture media and its association in leukoplakia and oral squamous cell carcinoma. J. Oral Maxillofac. Pathol. 2019, 23, 28–35. [Google Scholar] [CrossRef]
- McCullough, M.; Jaber, M.; Barrett, A.W.; Bain, L.; Speight, P.M.; Porter, S.R. Oral yeast carriage correlates with presence of oral epithelial dysplasia. Oral Oncol. 2002, 38, 391–393. [Google Scholar] [CrossRef]
- Gauch, L.M.R.; Pedrosa, S.S.; Silveira-Gomes, F.; Esteves, R.A.; Marques-da-Silva, S.H. Isolation of Candida spp. from denture-related stomatitis in Pará, Brazil. Braz. J. Microbiol. 2018, 49, 148–151. [Google Scholar] [CrossRef]
- Yano, J.; Yu, A.; Fidel, P.L., Jr.; Noverr, M.C. Candida glabrata Has No Enhancing Role in the Pathogenesis of -Associated Denture Stomatitis in a Rat Model. mSphere 2019, 4, e00191-19. [Google Scholar] [CrossRef]
- Gleysteen, J.J. A history of intragastric balloons. Surg. Obes. Relat. Dis. 2016, 12, 430–435. [Google Scholar] [CrossRef]
- Schapiro, M.; Benjamin, S.; Blackburn, G.; Frank, B.; Heber, D.; Kozarek, R.; Randall, S.; Stern, W. Obesity and the gastric balloon: A comprehensive workshop. Gastrointest. Endosc. 1987, 33, 323–327. [Google Scholar] [CrossRef]
- Neto, M.G.; Silva, L.B.; Grecco, E.; de Quadros, L.G.; Teixeira, A.; Souza, T.; Scarparo, J.; Parada, A.A.; Dib, R.; Moon, R.; et al. Brazilian Intragastric Balloon Consensus Statement (BIBC): Practical guidelines based on experience of over 40,000 cases. Surg. Obes. Relat. Dis. 2018, 14, 151–159. [Google Scholar] [CrossRef]
- Vargas, E.J.; Pesta, C.M.; Bali, A.; Ibegbu, E.; Bazerbachi, F.; Moore, R.L.; Kumbhari, V.; Sharaiha, R.Z.; Curry, T.W.; DosSantos, G.; et al. Single Fluid-Filled Intragastric Balloon Safe and Effective for Inducing Weight Loss in a Real-World Population. Clin. Gastroenterol. Hepatol. 2018, 16, 1073–1080.e1. [Google Scholar] [CrossRef]
- Coskun, H.; Bozkurt, S. A case of asymptomatic fungal and bacterial colonization of an intragastric balloon. World J. Gastroenterol. 2009, 15, 5751–5753. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Vasilaki, O.; Stefanidou, C.; Grosomanidis, V. Candida albicans colonization on an intragastric balloon. Asian J. Endosc. Surg. 2013, 6, 214–216. [Google Scholar] [CrossRef]
- Şimşek, Z.; Gürbüz, O.A.; Çoban, Ş. Fungal colonization of intragastric balloons. Endoscopy 2014, 46 (Suppl. S1), E642–E643. [Google Scholar] [CrossRef]
- Marques, L.M.; De Souza, T.F.; Grecco, E.; Neto, M.G.; Ramos, F.M.; Vieira, F.M.; Garcia, V.G.; Freitas, C.E., Jr. Proposed treatment of adjustable intragastric balloon contaminated with Candida. Bariatr. Surg. Pract. Patient Care 2015, 10, 169–172. [Google Scholar] [CrossRef]
- Barola, S.; Agnihotri, A.; Chang Chiu, A.; Kalloo, A.N.; Kumbhari, V. Spontaneous Hyperinflation of an Intragastric Balloon 5 Months After Insertion. Am. J. Gastroenterol. 2017, 112, 412. [Google Scholar] [CrossRef]
- Usuy, E.; Silva, M.; Dos Passos Galvão Neto, M.; Grecco, E.; Ferreira de Souza, T.; de Quadros, L.G. Antibiotics to Prevent Relapse of Adjustable Gastric Balloon Hyperinflation: Feasible for Balloon Maintenance? GE Port. J. Gastroenterol. 2020, 28, 52–55. [Google Scholar] [CrossRef]
- Prado, A.; de Oliveira Brito, R.; da Cruz Alves Pereira, E.; Alexandrino Becker, T.C.; Negri, M.; Inez Estivalet Svidzinski, T. The fungus Candida glabrata in intragastric balloon. Int. J. Case Rep. Images 2021, 12, 1–7. [Google Scholar] [CrossRef]
- Hawa, F.; Vargas, E.J.; Acosta, A.; McRae, A.; Bazerbachi, F.; Abu Dayyeh, B.K. Contamination of single fluid-filled intragastric balloons with orogastric fluid is not associated with hyperinflation: An ex-vivo study and systematic review of literature. BMC Gastroenterol. 2021, 21, 286. [Google Scholar] [CrossRef]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections. Methods Mol. Biol. 2017, 1508, 17–65. [Google Scholar] [CrossRef]
- Pramodhini, S.; Srirangaraj, S.; Easow, J.M. Candiduria-Study of Virulence Factors and Its Antifungal Susceptibility Pattern in Tertiary Care Hospital. J. Lab. Physicians 2021, 13, 231–237. [Google Scholar] [CrossRef]
- Songur Dagli, S.; Demir, T. Comparison of cervico-vaginal colonization among sexually active women by intrauterine device use. J. Infect. Dev. Ctries 2015, 9, 930–935. [Google Scholar] [CrossRef]
- Borges, K.R.A.; Pimentel, I.V.; Lucena, L.C.L.D.S.; Da Silva, M.A.C.N.; Monteiro, S.G.; Monteiro, C.A.; Nascimento, M.D.D.S.B.; Bezerra, G.F.D.B. Adhesion and biofilm formation of Candida parapsilosis isolated from vaginal secretions to copper intrauterine devices. Rev. Inst. Med. Trop. Sao Paulo 2018, 60, e59. [Google Scholar] [CrossRef]
- Auler, M.E.; Morreira, D.; Rodrigues, F.F.; Ão, M.S.A.; Margarido, P.F.; Matsumoto, F.E.; Silva, E.G.; Silva, B.C.; Schneider, R.P.; Paula, C.R. Biofilm formation on intrauterine devices in patients with recurrent vulvovaginal candidiasis. Med. Mycol. 2010, 48, 211–216. [Google Scholar] [CrossRef]
- Calışkan, S.; Keçeli Özcan, S.; Cınar, S.; Corakçı, A.; Calışkan, E. In vitro biofilm formation and relationship with antifungal resistance of Candida spp. isolated from vaginal and intrauterine device string samples of women with vaginal complaints. Mikrobiyoloji Bul. 2011, 45, 697–706. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22090300 (accessed on 4 July 2022).
- Zahran, K.M.; Agban, M.N.; Ahmed, S.H.; Hassan, E.A.; Sabet, M.A. Patterns of Candida biofilm on intrauterine devices. J. Med. Microbiol. 2015, 64 Pt 4, 375–381. [Google Scholar] [CrossRef]
- Donmez, H.G.; Sahal, G.; Akgor, U.; Cagan, M.; Ozgul, N.; Beksac, M.S. The relationship between the presence of HPV infection and biofilm formation in cervicovaginal smears. Infection 2020, 48, 735–740. [Google Scholar] [CrossRef]
- Urban & Vogel. In den Hohlräumen sprosst munter die Candida. MMW Fortschr. Med. 2013, 156, 7. [Google Scholar] [CrossRef]
- Shemer, A.; Trau, H.; Davidovici, B.; Grunwald, M.H.; Amichai, B. Onycomycosis due to artificial nails. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 998–1000. [Google Scholar] [CrossRef]
- Machado, P.; Pereira, D.; Morais, P. An itchy rash on the fingertips: A case study. Aust. Fam. Physician 2014, 43, 36–37. [Google Scholar] [CrossRef]
- Toukabri, N.; Dhieb, C.; El Euch, D.; Rouissi, M.; Mokni, M.; Sadfi-Zouaoui, N. Prevalence, Etiology, and Risk Factors of Tinea pedis and Tinea unguium in Tunisia. Can. J. Infect. Dis. Med. Microbiol. 2017, 2017, 6835725. [Google Scholar] [CrossRef]
- Jazdarehee, A.; Malekafzali, L.; Lee, J.; Lewis, R.; Mukovozov, I. Transmission of Onychomycosis and Dermatophytosis between Household Members: A Scoping Review. J. Fungi 2022, 8, 60. [Google Scholar] [CrossRef]
- Gupta, A.K.; Versteeg, S.G. The Role of Shoe and Sock Sanitization in the Management of Superficial Fungal Infections of the Feet. J. Am. Podiatr. Med. Assoc. 2019, 109, 141–149. [Google Scholar] [CrossRef]
- Klafke, G.B.; Silva RA da Pellegrin KT de Xavier, M.O. Analysis of the role of nail polish in the transmission of onychomycosis. An. Bras. Dermatol. 2018, 93, 930–931. [Google Scholar] [CrossRef]
- Van Bael, S.A.; Estrada, C.; Wcislo, W.T. Fungal-fungal interactions in leaf-cutting ant agriculture. Psyche 2011, 2011, 617478. [Google Scholar] [CrossRef]
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial interactions: Impact on pathogenesis and human disease. Clin. Microbiol. Rev. 2012, 25, 193–213. [Google Scholar] [CrossRef]
- Ellepola, K.; Truong, T.; Liu, Y.; Lin, Q.; Lim, T.K.; Lee, Y.M.; Cao, T.; Koo, H.; Seneviratne, C.J. Multi-omics Analyses Reveal Synergistic Carbohydrate Metabolism in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Infect. Immun. 2019, 87, e00339-19. [Google Scholar] [CrossRef]
- Palková, Z.; Váchová, L. Spatially structured yeast communities: Understanding structure formation and regulation with omics tools. Comput. Struct. Biotechnol. J. 2021, 19, 5613–5621. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef]
- Rodríguez-Cerdeira, C.; Gregorio, M.C.; Molares-Vila, A.; López-Barcenas, A.; Fabbrocini, G.; Bardhi, B.; Sinani, A.; Sánchez-Blanco, E.; Arenas-Guzmán, R.; Hernandez-Castro, R. Biofilms and vulvovaginal candidiasis. Colloids Surf. B Biointerfaces 2019, 174, 110–125. [Google Scholar] [CrossRef]
- Li, Q.; Liu, J.; Shao, J.; Da, W.; Shi, G.; Wang, T.; Wu, D.; Wang, C. Decreasing Cell Population of Individual Species Does Not Impair the Virulence of and Mixed Biofilms. Front. Microbiol. 2019, 10, 1600. [Google Scholar] [CrossRef]
- Gelotar, P.; Vachhani, S.; Patel, B.; Makwana, N. The prevalence of fungi in fingernail onychomycosis. J. Clin. Diagn. Res. 2013, 7, 250–252. [Google Scholar] [CrossRef]
- Lagacé, J.; Cellier, E. A case report of a mixed Chaetomium globosum/Trichophyton mentagrophytes onychomycosis. Med. Mycol. Case Rep. 2012, 1, 76–78. [Google Scholar] [CrossRef]
- Rossoni, R.D.; Barbosa, J.O.; Vilela, S.F.G.; Santos, J.D.D.; de Barros, P.P.; Prata, M.C.D.A.; Anbinder, A.L.; Fuchs, B.B.; Jorge, A.O.C.; Mylonakis, E.; et al. Competitive Interactions between C. albicans, C. glabrata and C. krusei during Biofilm Formation and Development of Experimental Candidiasis. PLoS ONE 2015, 10, e0131700. [Google Scholar] [CrossRef]
- Taj-Aldeen, S.J.; El Shafie, S.; Alsoub, H.; Eldeeb, Y.; de Hoog, G.S. Isolation of Exophiala dermatitidis from endotracheal aspirate of a cancer patient. Mycoses 2006, 49, 504–509. [Google Scholar] [CrossRef]
- Sadamatsu, H.; Takahashi, K.; Tashiro, H.; Ogusu, S.; Haraguchi, T.; Nakashima, C.; Nakamura, T.; Sueoka-Aragane, N. A rare case of Trichosporon mycotoxinivorans and Cryptococcus neoformans co-infection in lung. J. Infect. Chemother. 2020, 26, 838–842. [Google Scholar] [CrossRef]
- Wu, C.Y.; Huang, H.K.; Wu, P.K.; Chen, W.M.; Lai, M.C.; Chung, L.H. Acremonium species combined with Penicillium species infection in hip hemiarthroplasty: A case report and literature review. Hip Int. 2014, 24, 656–659. [Google Scholar] [CrossRef]
- Falanga, A.; Maione, A.; La Pietra, A.; de Alteriis, E.; Vitale, S.; Bellavita, R.; Carotenuto, R.; Turrà, D.; Galdiero, S.; Galdiero, E.; et al. Competitiveness during Dual-Species Biofilm Formation of Fusarium oxysporum and Candida albicans and a Novel Treatment Strategy. Pharmaceutics 2022, 14, 1167. [Google Scholar] [CrossRef]
- Ruiz, A.; Herráez, M.; Costa-Gutierrez, S.B.; Molina-Henares, M.A.; Martínez, M.J.; Espinosa-Urgel, M.; Barriuso, J. The architecture of a mixed fungal-bacterial biofilm is modulated by quorum-sensing signals. Environ. Microbiol. 2021, 23, 2433–2447. [Google Scholar] [CrossRef]
- Zarnowski, R.; Sanchez, H.; Covelli, A.S.; Dominguez, E.; Jaromin, A.; Bernhardt, J.; Mitchell, K.F.; Heiss, C.; Azadi, P.; Mitchell, A.; et al. Candida albicans biofilm-induced vesicles confer drug resistance through matrix biogenesis. PLoS Biol. 2018, 16, e2006872. [Google Scholar] [CrossRef]
- Olson, M.L.; Jayaraman, A.; Kao, K.C. Relative Abundances of Candida albicans and Candida glabrata in Coculture Biofilms Impact Biofilm Structure and Formation. Appl. Environ. Microbiol. 2018, 84, e02769-17. [Google Scholar] [CrossRef]
- Dos Santos, J.D.; Piva, E.; Vilela, S.F.G.; Jorge, A.O.C.; Junqueira, J.C. Mixed biofilms formed by C. albicans and non-albicans species: A study of microbial interactions. Braz. Oral Res. 2016, 30. [Google Scholar] [CrossRef] [Green Version]
- d’Enfert, C.; Janbon, G. Biofilm formation in Candida glabrata: What have we learnt from functional genomics approaches? FEMS Yeast Res. 2016, 16, fov111. [Google Scholar] [CrossRef]
- Gonçalves, B.; Azevedo, N.; Osório, H.; Henriques, M.; Silva, S. Revealing Candida glabrata biofilm matrix proteome: Global characterization and pH response. Biochem. J. 2021, 478, 961–974. [Google Scholar] [CrossRef]
- Purohit, D.; Gajjar, D. Tec1 and Ste12 transcription factors play a role in adaptation to low pH stress and biofilm formation in the human opportunistic fungal pathogen Candida glabrata. Int. Microbiol. 2022, 1–14. [Google Scholar] [CrossRef]
- Capote-Bonato, F.; Sakita, K.M.; de Oliveira Junior, A.G.; de Souza Bonfim-Mendonça, P.; Crivellenti, L.Z.; Negri, M.; Svidzinski, T.I.E. In vitro interaction of Candida tropicalis biofilm formed on catheter with human cells. Microb. Pathog. 2018, 125, 177–182. [Google Scholar] [CrossRef]
- Capote-Bonato, F.; Bonato, D.V.; Ayer, I.M.; Magalhães, L.F.; Magalhães, G.M.; Barros, F.F.P.D.C.; Teixeira, P.P.M.; Crivellenti, L.Z.; Negri, M.; Svidzinski, T.I.E. Murine model for the evaluation of candiduria caused by Candida tropicalis from biofilm. Microb. Pathog. 2018, 117, 170–174. [Google Scholar] [CrossRef]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef]
- Kowalski, C.H.; Morelli, K.A.; Schultz, D.; Nadell, C.D.; Cramer, R.A. Fungal biofilm architecture produces hypoxic microenvironments that drive antifungal resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 22473–22483. [Google Scholar] [CrossRef]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal Drug Resistance: Molecular Mechanisms in and Beyond. Chem. Rev. 2021, 121, 3390–3411. [Google Scholar] [CrossRef]
- Truong, T.; Zeng, G.; Qingsong, L.; Kwang, L.T.; Tong, C.; Chan, F.Y.; Wang, Y.; Seneviratne, C.J. Comparative Ploidy Proteomics of Candida albicans Biofilms Unraveled the Role of the AHP1 Gene in the Biofilm Persistence Against Amphotericin B. Mol. Cell Proteom. 2016, 15, 3488–3500. [Google Scholar] [CrossRef]
- Lafleur, M.D.; Qi, Q.; Lewis, K. Patients with long-term oral carriage harbor high-persister mutants of Candida albicans. Antimicrob. Agents Chemother. 2010, 54, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Denega, I.; d’Enfert, C.; Bachellier-Bassi, S. Biofilms Are Generally Devoid of Persister Cells. Antimicrob. Agents Chemother. 2019, 63, e01979-18. [Google Scholar] [CrossRef]
- Al-Dhaheri, R.S.; Douglas, L.J. Absence of amphotericin B-tolerant persister cells in biofilms of some Candida species. Antimicrob. Agents Chemother. 2008, 52, 1884–1887. [Google Scholar] [CrossRef]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. The Extracellular Matrix of Fungal Biofilms. Adv. Exp. Med. Biol. 2016, 931, 21–35. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Pesee, S.; Angkananuwat, C.; Tancharoensukjit, S.; Muanmai, S.; Sirivan, P.; Bubphawas, M.; Tanarerkchai, N. In vitro activity of Caspofungin combined with Fluconazole on mixed Candida albicans and Candida glabrata biofilm. Med. Mycol. 2016, 54, 384–393. [Google Scholar] [CrossRef]
- Shanahan, F.; Ghosh, T.S.; O’Toole, P.W. The Healthy Microbiome-What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef]
- Veiga, F.F.; de Castro-Hoshino, L.V.; Sato, F.; Bombassaro, A.; A Vicente, V.; Mendes, V.; Baesso, M.L.; Negri, M.; Svidzinski, T.I. Fusarium oxysporum is an onychomycosis etiopathogenic agent. Future Microbiol. 2018, 13, 1745–1756. [Google Scholar] [CrossRef]
- Coad, B.R.; Kidd, S.E.; Ellis, D.H.; Griesser, H.J. Biomaterials surfaces capable of resisting fungal attachment and biofilm formation. Biotechnol. Adv. 2014, 32, 296–307. [Google Scholar] [CrossRef]
- Ghannoum, M.; Roilides, E.; Katragkou, A.; Petraitis, V.; Walsh, T.J. The Role of Echinocandins in Candida Biofilm-Related Vascular Catheter Infections: In vitro and In vivo Model Systems. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S618–S621. [Google Scholar] [CrossRef]
- Thieme, L.; Hartung, A.; Tramm, K.; Klinger-Strobel, M.; Jandt, K.D.; Makarewicz, O.; Pletz, M.W. MBEC versus MBIC: The lack of differentiation between biofilm reducing and inhibitory effects as a current problem in biofilm methodology. Biol. Proced. Online 2019, 21, 18. [Google Scholar] [CrossRef]
- Garaicoa, J.L.; Fischer, C.L.; Bates, A.M.; Holloway, J.; Avila-Ortiz, G.; Guthmiller, J.M.; Johnson, G.K.; Stanford, C.; Brogden, K.A. Promise of combining antifungal agents in denture adhesives to Fight Candida Species infections. J. Prosthodont. 2018, 27, 755–762. [Google Scholar] [CrossRef]
- Ionescu, A.C.; Brambilla, E.; Sighinolfi, M.C.; Mattina, R. A new urinary catheter design reduces in-vitro biofilm formation by influencing hydrodynamics. J. Hosp. Infect. 2021, 114, 153–162. [Google Scholar] [CrossRef]
- Bajunaid, S.O.; Baras, B.H.; Balhaddad, A.A.; Weir, M.D.; Xu, H.H.K. Antibiofilm and Protein-Repellent Polymethylmethacrylate Denture Base Acrylic Resin for Treatment of Denture Stomatitis. Materials 2021, 14, 1067. [Google Scholar] [CrossRef]
- D’Adamio, S.; Ziranu, A.; Cazzato, G.; Sanguinetti, M.; Manicone, P.; Rosa, M.; Maccauro, G. Antifungal properties of silver coating on tumour endoprostheses: An in vitro study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23 (Suppl. S2), 252–257. [Google Scholar] [CrossRef]
- Takamiya, A.S.; Monteiro, D.R.; Gorup, L.F.; Silva, E.A.; Camargo, E.R.; Gomes-Filho, J.E.; De Oliveira, S.H.P.; Barbosa, D.B. Biocompatible silver nanoparticles incorporated in acrylic resin for dental application inhibit Candida albicans biofilm. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 118, 111341. [Google Scholar] [CrossRef]


{kind=link}
| Surface | Type of Study | Research Objective | Fungi Involved | MFB Highlights | Ref |
|---|---|---|---|---|---|
| Skin | Clinical | Authors discuss the reports of antifungal resistance from around the world, present their experience with treatment-resistant infections, and examine alternative treatment strategies | Trichophyton rubrum, Aspergillus spp. and Scopulariopsis brevicaulis Dermatophyte and NDM | Therapy resistance probably accentuated due to the presence of mixed-infection-associated drugs, meaning a long treatment time and recurrence after the end of the therapy | [60] |
| Scalp | Clinical | An analysis of the clinical features and laboratory findings associated with a Tinea capitis infection outbreak in young novice Buddhist monks | Trichophyton mentagrophytes and Microsporum canis | T. mentagrophytes and M. canis were the predominantly isolated mixed dermatophyte pathogens and an extensive area of infection was significantly associated with mixed-type clinical presentation | [61] |
| Scalp | Clinical | A clinical case of white scaly alopecia on the scalp of prepubertal children | Trichophyton violaceum (violet) and Trichophyton violaceum (white) | Before treatment with griseofulvin, T. violaceum (white variant) was isolated. After treatment, T. violaceum (violet variant) was isolated, indicating the possibility of mixed infection with both variants of T. violaceum: white and violet | [10] |
| Scalp | Clinical | To describe clinical manifestations of Tinea capitis in children in southwestern Uganda and identify the main pathogen | Microsporum audouinii and Trichophyton violaceum Trichophyton violaceum and Scopulariopsis brevicaulis Trichophyton soudanense and Microsporum audouinii Trichophyton violaceum, Trichophyton soudanense and Trichophyton rubrum | Several fungal species known to be pathogenic were found in association, affecting a group of patients with a developing immune system. In addition, in this country, there is a difficulty in the treatment due to there being only a few medication options | [62] |
| Nails | Clinical | A case of co-habitation of fungus-fungus as causative agents of onychomycosis in a healthy male | Trichosporon asahii and Rhodotorula mucilaginosa | The case report revealed the presence of dermatophyte and non-dermatophyte in the toenail, highlighting the co-habitation of T. asahii and R. mucilaginosa in the causation of onychomycosis and to raise the awareness of this infection among dermatologists | [68] |
| Nails | Clinical | To investigate the clinical manifestations, risk factors, and treatment outcomes of mixed-infection onychomycosis | Trichophyton mentagrophytes and Neoscytalidium dimidiatum Trichophyton rubrum and Neoscytalidium dimidiatum | The time of oral treatment for the mixed-infection group was significantly longer than that for the dermatophytes group | [67] |
| Nails | Clinical | The authors aimed to evaluate the feasibility of introducing microbiological techniques in the diagnosis of nail diseases based only on clinical parameters | Candida albicans and Trichophyton rubrum Candida albicans and Trichophyton mentagrophytes | Not surprisingly, C. albicans was the most isolated species causing finger nail onychomycosis, but a fact that deserves importance is its association with dermatophytes, mainly due to a diagnostic that demands a lot of experience and care from the laboratory mycologist | [136] |
| Nails | Clinical | To prove a clinical case of a mixed onychomycosis infection of a toenail | Chaetomium globosum and Trichophyton mentagrophytes | This association was proved for the first time | [137] |
| Nasal cavity and paranasal sinuses | Clinical | To report 10 cases of mixed invasive fungal in COVID-19 patients and their outcomes | Rhizopus arrhizus and Aspergillus flavus Rhizopus arrhizus and Aspergillus fumigatus | Mixed fungal infection must be valued, correctly identified and treated, reducing comorbidities for the COVID-19 patient | [5] |
| Oral | In vivo and in vitro | To evaluate the interaction of MFB in vitro, in vivo with murine models of experimental candidiasis and Galleria mellonella larvae | Candida albicans and Candida krusei Candida albicans and Candida glabrata | Single infections by C. albicans were more harmful for animal models than mixed infections with NCAC species, suggesting that C. albicans establish competitive interactions with C. krusei and C. glabrata during biofilm formation | [138] |
| Endotra- cheal aspirates | Clinical | Reported a respiratory tract colonization of E. dermatitidis in a cancer patient suffering from C. krusei fungemia and pulmonary disorder | Exophiala dermatitidis and Candida krusei | The patient’s death was attributed to cancer associated with C. krusei fungemia, but probably E. dermatitidis also played a role in the morbidity of the case | [139] |
| Lung | Clinical | This study showed a case of pulmonary co-habitation of two fungal species, T. mycotoxinivorans and C. neoformans | Trichosporon mycotoxinivorans and Cryptococcus neoformans | Case of co-infection of the lung with T. mycotoxinivorans and C. neoformans. This is the first report of T. mycotoxinivorans respiratory infection in Japan | [140] |
| ETT | Clinical | Aimed to determine the frequency of yeast colonization in the tracheobronchial secretions of critically ill intubated patients and to assess the presence of these yeasts in the infra-cuff region of the ETT | Candida glabrata and Candida tropicalis Candida glabrata and Candida albicans | NCAC species were found in co-colonization conditions. More than one species was isolated from both tracheobronchial secretion and ETT in 25% of the patients colonized by Candida spp. | [70] |
| IGB | Clinical | Determine the frequency of biofilms naturally formed on the external surface of IGB, as well as some variables related to IGB types and patients features, species of fungi involved and biofilm evidence | Candida glabrata and Candida albicans Candida albicans and Candida krusei Candida glabrata and Candida tropicalis Candida glabrata, Candida tropicalis and Candida krusei | Several highly pathogenic fungal species were found, forming mixed biofilms highly adapted to a hostile environment | [46] |
| Urinary catheter, stent urinary, and urine from these devices | In situ and in vitro | To detect Candida spp. using molecular detection by capillary electrophoresis | Candida albicans and Candida parapsilosis Candida albicans and Candida robusta Candida albicans and Candida krusei | The f-ITS2-PCR-CE method was more sensitive and more specific than routine culture both in mono and poly species in the Candida colonization | [71] |
| Intravascular catheter | In situ and in vitro | To search mixed fungal biofilm formed on the intravascular catheter | Candida albicans and Candida glabrata | C. albicans and C. glabrata can competitively and symbiotically coexist in a mixed biofilm | [72] |
| Hip prosthesis | In vivo | To illustrate that Acremonium and Penicillium species are being increasingly recognized in periprosthetic joint infections | Acremonium spp. and Penicillium spp. | The authors gave due importance to the growth of commonly neglected fungi and were able to importantly report the positivity of intraoperative mixed fungal cultures | [141] |
| Polystyrene and Zophobas morio larvae | In vitro and in vivo | To evaluate the pathogenesis of a co-infection by two less common yeasts on Z. morio larvae | Rhodotorula mucilaginosa and Trichosporon asahii | Yeasts cells recovered from an in vitro biofilm provoked increased death rates of larvae infected for mixed suspensions | [56] |
| Polystyrene | In vitro | To investigate the interaction in a dual-species biofilm, considering variable formation conditions | Candida albicans and Fusarium oxysporum | The total biomass of the dual-species biofilm was significantly lower in comparison to the single biofilm of F. oxysporum but superior to that of the single C. albicans biofilm | [142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, P.d.S.; Prado, A.; Bagon, N.P.; Negri, M.; Svidzinski, T.I.E. Mixed Fungal Biofilms: From Mycobiota to Devices, a New Challenge on Clinical Practice. Microorganisms 2022, 10, 1721. https://doi.org/10.3390/microorganisms10091721
Costa PdS, Prado A, Bagon NP, Negri M, Svidzinski TIE. Mixed Fungal Biofilms: From Mycobiota to Devices, a New Challenge on Clinical Practice. Microorganisms. 2022; 10(9):1721. https://doi.org/10.3390/microorganisms10091721
Chicago/Turabian StyleCosta, Polyana de Souza, Andressa Prado, Natalia Pecin Bagon, Melyssa Negri, and Terezinha Inez Estivalet Svidzinski. 2022. "Mixed Fungal Biofilms: From Mycobiota to Devices, a New Challenge on Clinical Practice" Microorganisms 10, no. 9: 1721. https://doi.org/10.3390/microorganisms10091721