
Loss of rpoE Encoding the δ-Factor of RNA Polymerase Impacts Pathophysiology of the Streptococcus pyogenes M1T1 Strain 5448

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Insertional Mutagenesis and Complementation
2.3. Murine Subcutaneous Infection Model
2.4. In Vivo Competition Assay
2.5. Lancefield Bactericidal Assay
2.6. In Vitro Growth Studies
2.7. RNA-Seq and Data Analysis
2.8. Extracellular Acidification Assay
2.9. Ethics Statement
3. Results
3.1. An rpoE Mutant in the M1T1 GAS Strain 5448 Is Attenuated for Virulence
3.2. The 5448 WT Strain Outcompetes the rpoE Mutant When Co-Infected in the Skin and Soft-Tissue Infection Model
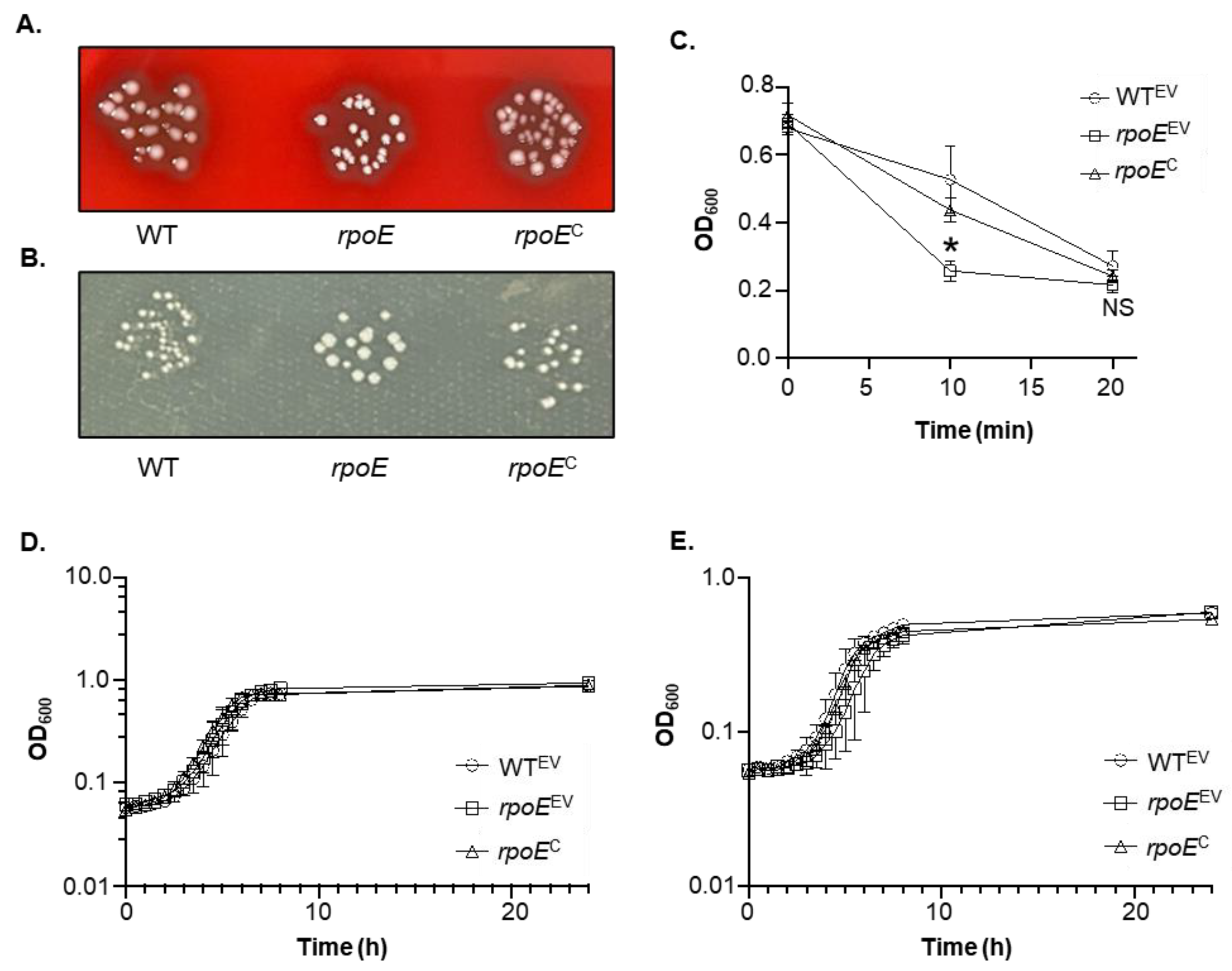
3.3. Mutation of rpoE Limits the Ability of GAS 5448 to Survive in Human Blood
3.4. rpoE Mutants Produce a Small Colony Variant Phenotype, Enhanced Aggregation, and No Changes in Growth Kinetics Compared to WT
3.5. Mutation of rpoE Decreases Fitness at High Temperatures
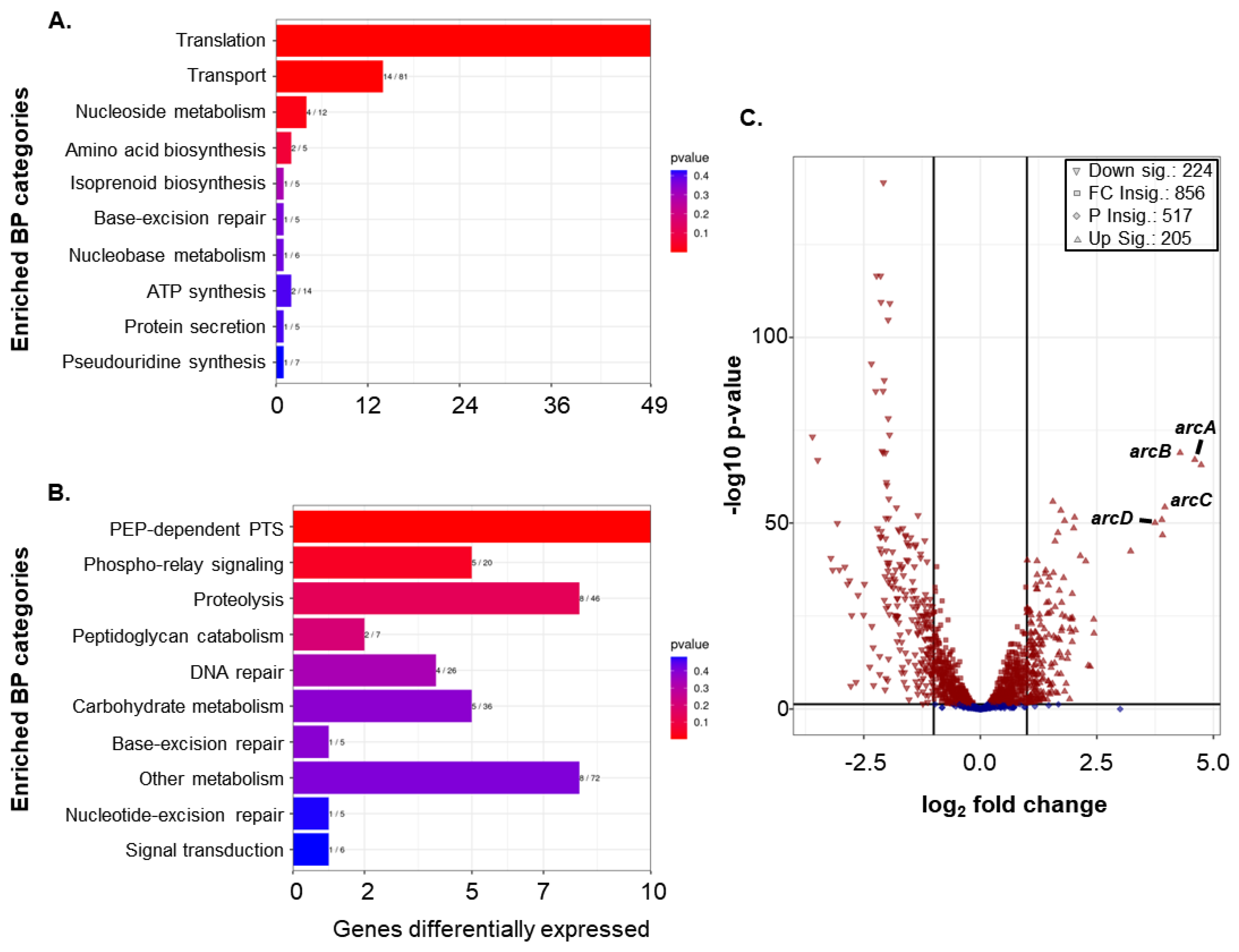
3.6. RNA-Seq Shows That Loss of rpoE Alters Global Gene Expression
3.7. Mutation of rpoE Enhances Arc Operon Expression during Growth of GAS in Liquid Culture Media
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. Lancet Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Macris, M.H.; Hartman, N.; Murray, B.; Klein, R.F.; Roberts, R.B.; Kaplan, E.L.; Horn, D.; Zabriskie, J.B. Studies of the continuing susceptibility of group A streptococcal strains to penicillin during eight decades. Pediatr. Infect. Dis. J. 1998, 17, 377–381. [Google Scholar] [CrossRef]
- Abraham, T.; Sistla, S. Trends in Antimicrobial Resistance Patterns of Group A Streptococci, Molecular Basis and Implications. Indian J. Med. Microbiol. 2018, 36, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Ben Zakour, N.; Davies, M.; You, Y.; Chen, J.H.K.; Forde, B.; Stanton-Cook, M.; Yang, R.; Cui, Y.; Barnett, T.; Venturini, C.; et al. Transfer of scarlet fever-associated elements into the group A Streptococcus M1T1 clone. Sci. Rep. 2015, 5, 15877. [Google Scholar] [CrossRef] [PubMed]
- Seppälä, H.; Skurnik, M.; Soini, H.; Roberts Marilyn, C.; Huovinen, P. A Novel Erythromycin Resistance Methylase Gene (ermTR) in Streptococcus pyogenes. Antimicrob. Agents Chemother. 1998, 42, 257–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, T.M.; Gross, C.A. Multiple Sigma Subunits and the Partitioning of Bacterial Transcription Space. Annu. Rev. Microbiol. 2003, 57, 441–466. [Google Scholar] [CrossRef]
- Ma, C.; Yang, X.; Lewis, P.J. Bacterial Transcription as a Target for Antibacterial Drug Development. Microbiol. Mol. Biol. Rev. 2016, 80, 139–160. [Google Scholar] [CrossRef] [Green Version]
- Pero, J.; Nelson, J.; Fox, T.D. Highly asymmetric transcription by RNA polymerase containing phage-SP01-induced polypeptides and a new host protein. Proc. Natl. Acad. Sci. USA 1975, 72, 1589–1593. [Google Scholar] [CrossRef] [Green Version]
- Feklístov, A.; Sharon, B.D.; Darst, S.A.; Gross, C.A. Bacterial Sigma Factors: A Historical, Structural, and Genomic Perspective. Annu. Rev. Microbiol. 2014, 68, 357–376. [Google Scholar] [CrossRef] [Green Version]
- Helmann, J.D.; Chamberlin, M.J. Structure and Function of Bacterial Sigma Factors. Annu. Rev. Biochem. 1988, 57, 839–872. [Google Scholar] [CrossRef]
- Achberger, E.C.; Whiteley, H.R. The role of the delta peptide of the Bacillus subtilis RNA polymerase in promoter selection. J. Biol. Chem. 1981, 256, 7424–7432. [Google Scholar] [CrossRef]
- Dobinson, K.F.; Spiegelman, G.B. Effect of the delta subunit of Bacillus subtilis RNA polymerase on initiation of RNA synthesis at two bacteriophage phi 29 promoters. Biochemistry 1987, 26, 8206–8213. [Google Scholar] [CrossRef] [PubMed]
- Juang, Y.L.; Helmann, J.D. A Promoter Melting Region in the Primary sigma Factor of Bacillus subtilis: Identification of Functionally Important Aromatic Amino Acids. J. Mol. Biol. 1994, 235, 1470–1488. [Google Scholar] [CrossRef] [PubMed]
- Juang, Y.-L.; Helmann, J.D. Pathway of promoter melting by Bacillus subtilis RNA polymerase at a stable RNA promoter: Effects of temperature, delta protein, and sigma factor mutations. Biochemistry 1995, 34, 8465–8473. [Google Scholar] [CrossRef]
- Weiss, A.; Shaw, L.N. Small things considered: The small accessory subunits of RNA polymerase in Gram-positive bacteria. FEMS Microbiol. Rev. 2015, 39, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Watson, S.P.; Antonio, M.; Foster, S.J. Isolation and characterization of Staphylococcus aureus starvation-induced, stationary-phase mutants defective in survival or recovery. Microbiology 1998, 144, 3159–3169. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.L.; Needham, R.H.V.; Rubens, C.E. The Delta Subunit of RNA Polymerase Is Required for Virulence of Streptococcus agalactiae. Infect. Immun. 2003, 71, 4011–4017. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Tomasch, J.; Sztajer, H.; Wagner-Döbler, I. The Delta Subunit of RNA Polymerase, RpoE, Is a Global Modulator of Streptococcus mutans Environmental Adaptation. J. Bacteriol. 2010, 192, 5081–5092. [Google Scholar] [CrossRef] [Green Version]
- López de Saro, F.J.; Yoshikawa, N.; Helmann, J.D. Expression, abundance, and RNA polymerase binding properties of the delta factor of Bacillus subtilis. J. Biol. Chem. 1999, 274, 15953–15958. [Google Scholar] [CrossRef] [Green Version]
- Rabatinová, A.; Šanderová, H.; Matějčková, J.J.; Korelusová, J.; Sojka, L.; Barvík, I.; Papoušková, V.; Sklenář, V.; Žídek, L.; Krásný, L. The δ Subunit of RNA Polymerase Is Required for Rapid Changes in Gene Expression and Competitive Fitness of the Cell. J. Bacteriol. 2013, 195, 2603–2611. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.L.; Knoll, K.M.; Rubens, C.E. Identification of Streptococcus agalactiae virulence genes in the neonatal rat sepsis model using signature-tagged mutagenesis. Mol. Microbiol. 2000, 37, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Ibarra, J.A.; Paoletti, J.; Carroll, R.K.; Shaw, L.N. The delta Subunit of RNA Polymerase Guides Promoter Selectivity and Virulence in Staphylococcus aureus. Infect. Immun. 2014, 82, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, L.; Huang, H.; Yang, C.; Yang, S.; Gu, Y.; Jiang, W. A Flexible Binding Site Architecture Provides New Insights into CcpA Global Regulation in Gram-Positive Bacteria. mBio 2017, 8, e02004-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, X.; Sztajer, H.; Buddruhs, N.; Petersen, J.; Rohde, M.; Talay, S.R.; Wagner-Döbler, I. Lack of the Delta Subunit of RNA Polymerase Increases Virulence Related Traits of Streptococcus mutans. PLoS ONE 2011, 6, e20075. [Google Scholar] [CrossRef]
- Chatellier, S.; Ihendyane, N.; Kansal, R.G.; Khambaty, F.; Basma, H.; Norrby-Teglund, A.; Low, D.E.; McGeer, A.; Kotb, M. Genetic Relatedness and Superantigen Expression in Group A Streptococcus Serotype M1 Isolates from Patients with Severe and Nonsevere Invasive Diseases. Infect. Immun. 2000, 68, 3523–3534. [Google Scholar] [CrossRef] [Green Version]
- Cleary, P.P.; Kaplan, E.L.; Handley, J.P.; Wlazlo, A.; Kim, M.H.; Hauser, A.R.; Schlievert, P.M. Clonal basis for resurgence of serious Streptococcus pyogenes disease in the 1980s. Lancet 1992, 339, 518–521. [Google Scholar] [CrossRef]
- Freiberg, J.A.; Le Breton, Y.; Harro, J.M.; Allison, D.L.; McIver, K.S.; Shirtliff, M.E. The Arginine Deiminase Pathway Impacts Antibiotic Tolerance during Biofilm-Mediated Streptococcus pyogenes Infections. mBio 2020, 11, e00919–e00920. [Google Scholar] [CrossRef]
- Le Breton, Y.; Belew, A.T.; Freiberg, J.A.; Sundar, G.S.; Islam, E.; Lieberman, J.; Shirtliff, M.E.; Tettelin, H.; El-Sayed, N.M.; McIver, K.S. Genome-wide discovery of novel M1T1 group A streptococcal determinants important for fitness and virulence during soft-tissue infection. PLoS Pathog. 2017, 13, e1006584. [Google Scholar] [CrossRef]
- Lertcanawanichakul, M. Construction of Plasmid Vector for Expression of Bacteriocin N15-Encoding Gene and Effect of Engineered Bacteria on Enterococcus faecalis. Curr. Microbiol. 2007, 54, 108–112. [Google Scholar] [CrossRef]
- Bryan, E.M.; Bae, T.; Kleerebezem, M.; Dunny, G.M. Improved vectors for nisin-controlled expression in gram-positive bac-teria. Plasmid 2000, 44, 183–190. [Google Scholar] [CrossRef]
- Le Breton, Y.; Belew, A.T.; Valdes, K.M.; Islam, E.; Curry, P.; Tettelin, H.; Shirtliff, M.E.; El-Sayed, N.M.; McIver, K.S. Essential Genes in the Core Genome of the Human Pathogen Streptococcus pyogenes. Sci. Rep. 2015, 5, 9838. [Google Scholar] [CrossRef] [Green Version]
- Braza, R.E.; Le Breton, Y.; McIver, K.S. The scfCDE Operon Encodes a Predicted ABC Importer Required for Fitness and Virulence during Group A Streptococcus Invasive Infection. Infect. Immun. 2019, 87, e00613–e00619. [Google Scholar] [CrossRef] [PubMed]
- Valdes, K.M.; Sundar, G.S.; Belew, A.T.; Islam, E.; El-Sayed, N.M.; Le Breton, Y.; McIver, K.S. Glucose Levels Alter the Mga Virulence Regulon in the Group A Streptococcus. Sci. Rep. 2018, 8, 4971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Jaffe, A.E.; Storey, J.D. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics 2012, 28, 882–883. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-Seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, Y.; Yamaguchi, M.; Sumitomo, T.; Nakata, M.; Hanada, T.; Okuzaki, D.; Motooka, D.; Mori, Y.; Kawasaki, H.; Coady, A.; et al. Streptococcus pyogenes upregulates arginine catabolism to exert its pathogenesis on the skin surface. Cell Rep. 2021, 34, 108924. [Google Scholar] [CrossRef] [PubMed]
- Churton, N.W.; Misra, R.V.; Howlin, R.P.; Allan, R.N.; Jefferies, J.; Faust, S.N.; Gharbia, S.E.; Edwards, R.J.; Clarke, S.C.; Webb, J.S. Parallel Evolution in Streptococcus pneumoniae Biofilms. Genome Biol. Evol. 2016, 8, 1316–1326. [Google Scholar] [CrossRef] [Green Version]
- Cusumano, Z.T.; Watson, M.E., Jr.; Caparon, M.G. Streptococcus pyogenes Arginine and Citrulline Catabolism Promotes Infection and Modulates Innate Immunity. Infect. Immun. 2014, 82, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griswold, A.; Chen, Y.-Y.M.; Snyder, J.A.; Burne, R.A. Characterization of the Arginine Deiminase Operon of Streptococcus rattus FA-1. Appl. Environ. Microbiol. 2004, 70, 1321–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease Manifestations and Pathogenic Mechanisms of Group A Streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Kotb, M. Rise and Persistence of Global M1T1 Clone of Streptococcus pyogenes. Emerg. Infect. Dis. 2008, 14, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Okamoto, A.; Miura, Y.; Tatsuno, I.; Maeyama, J.-I.; Hasegawa, T. Streptococcus pyogenes TrxSR Two-Component System Regulates Biofilm Production in Acidic Environments. Infect. Immun. 2021, 89, e0036021. [Google Scholar] [CrossRef]
- Xue, X.; Li, J.; Wang, W.; Sztajer, H.; Wagner-Döbler, I. The global impact of the delta subunit RpoE of the RNA polymerase on the proteome of Streptococcus mutans. Microbiology 2012, 158, 191–206. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid or Strain | Description | Reference |
|---|---|---|
| Plasmid | ||
| pMSP3535 | Derived from pAMβ1 with a ColE1 replicon, ErmR, with nisin inducible system (nisRK PnisA) | [30] |
| pOri253 | Derived from pIL253 (5.2 kb), 0.89-kb fragment of oriColE1 from pBluescript II KS | [29] |
| pPrpoE_rpoE | Derived from pOri253 and 1152 bp fragment of rpoE with ~400 bp of upstream intergenic DNA | This work |
| Strain | ||
| 5448 WT | Parental strain | [26] |
| 5448 EV | Wild type with empty vector pMSP3535 | This work |
| 5448 rpoE | rpoE::pSinS mutant | This work |
| 5448 rpoEEV | rpoE::pSinS mutant with empty pMSP3535 vector | This work |
| 5448 rpoEC | rpoE::pSinS mutant with pPrpoE_rpoE complementation vector | This work |
| 5448 arcA | arcA::pSinS mutant | [27] |
| 5448 Spy_1849 | Spy_1849::pSinS mutant | [28] |
| 5448 covS | covS::pSinS mutant | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rom, J.S.; Le Breton, Y.; Islam, E.; Belew, A.T.; El-Sayed, N.M.; McIver, K.S. Loss of rpoE Encoding the δ-Factor of RNA Polymerase Impacts Pathophysiology of the Streptococcus pyogenes M1T1 Strain 5448. Microorganisms 2022, 10, 1686. https://doi.org/10.3390/microorganisms10081686
Rom JS, Le Breton Y, Islam E, Belew AT, El-Sayed NM, McIver KS. Loss of rpoE Encoding the δ-Factor of RNA Polymerase Impacts Pathophysiology of the Streptococcus pyogenes M1T1 Strain 5448. Microorganisms. 2022; 10(8):1686. https://doi.org/10.3390/microorganisms10081686
Chicago/Turabian StyleRom, Joseph S., Yoann Le Breton, Emrul Islam, Ashton T. Belew, Najib M. El-Sayed, and Kevin S. McIver. 2022. "Loss of rpoE Encoding the δ-Factor of RNA Polymerase Impacts Pathophysiology of the Streptococcus pyogenes M1T1 Strain 5448" Microorganisms 10, no. 8: 1686. https://doi.org/10.3390/microorganisms10081686