
A Comparison of Rhizospheric and Endophytic Bacteria in Early and Late-Maturing Pumpkin Varieties


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Site Description
2.2. Test Materials
2.3. Soil Sample Collection
2.4. Soil Physicochemical and Biological Properties
2.5. Soil Bacterial DNA Extraction and Illumina Sequencing
2.6. Statistical Analysis
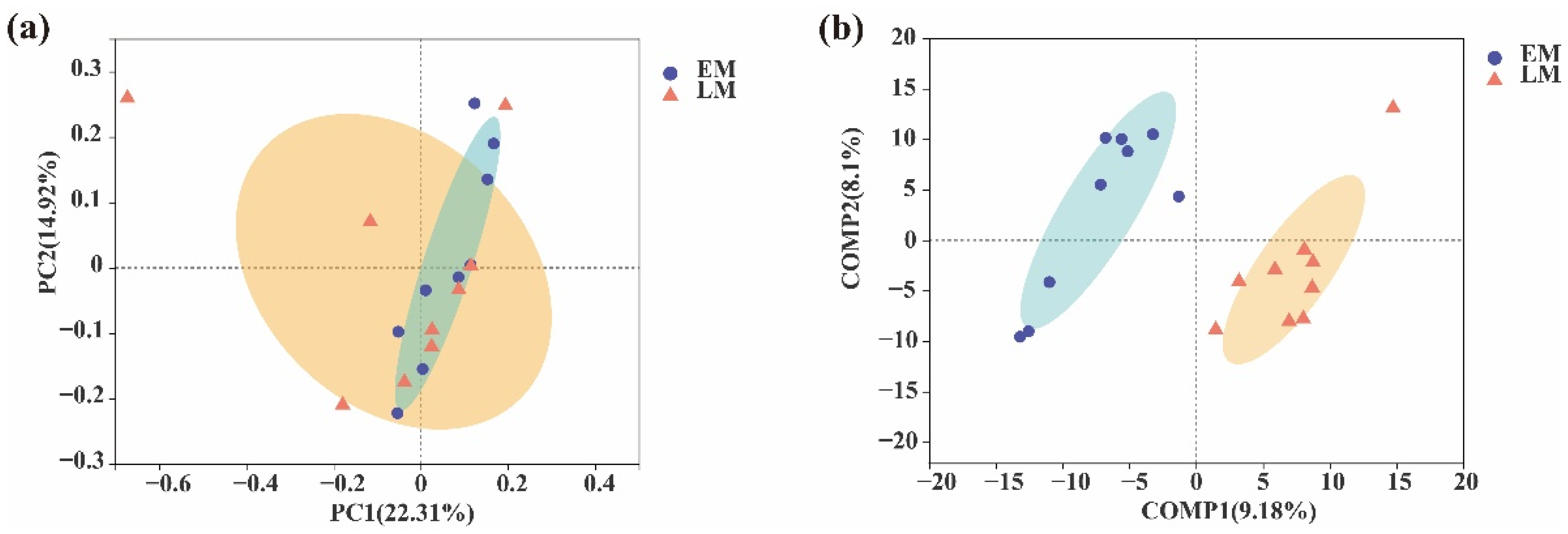
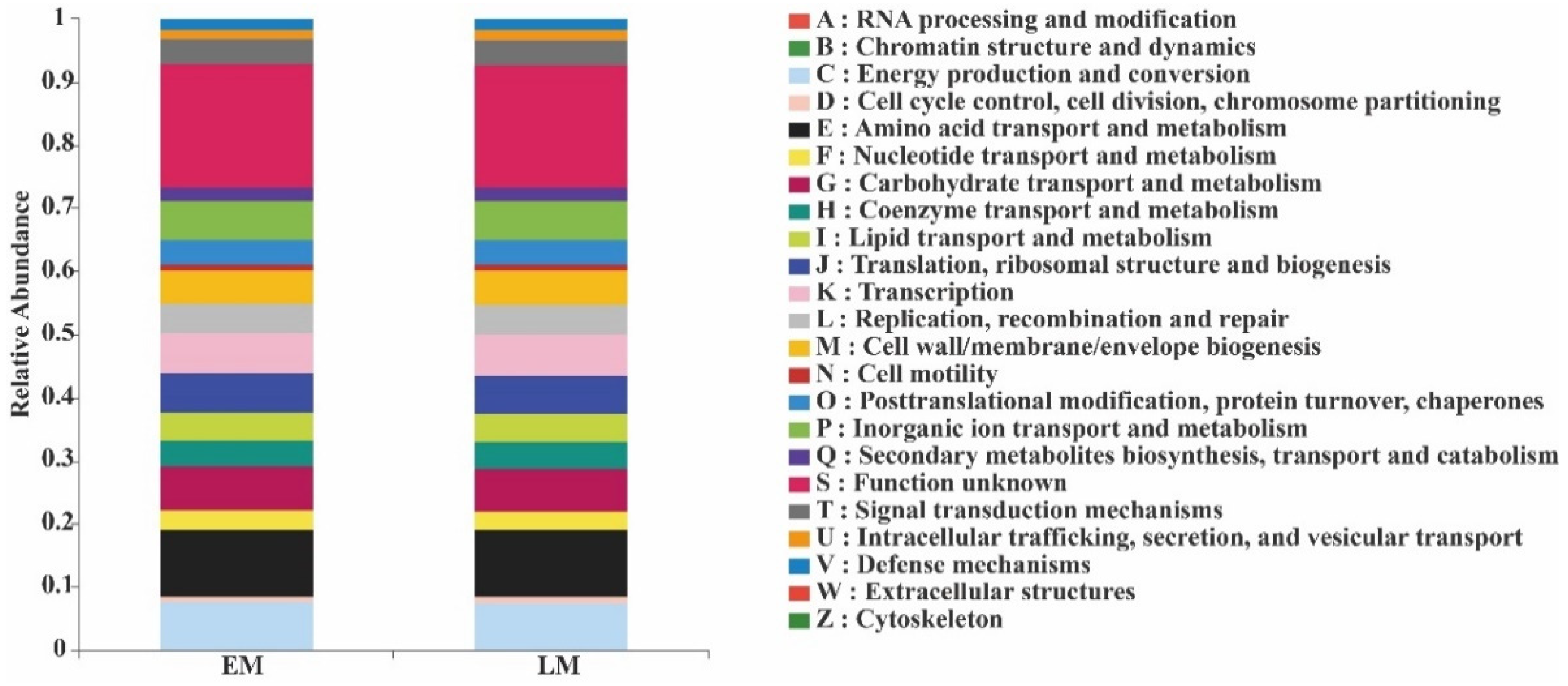
3. Results
4. Discussion
4.1. Effects of Early and Late-Maturing Pumpkin Varieties on Soil Enzymes
4.2. Effects of Early and Late-Maturing Pumpkin Varieties on Rhizosphere and Endophytic Bacteria Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dini, I.; Tenore, G.C.; Dini, A. Effect of industrial and domestic processing on antioxidant properties of pumpkin pulp. LWT Food Sci. Technol. 2013, 53, 382–385. [Google Scholar] [CrossRef]
- Murkovic, M.; Mülleder, U.; Neunteufl, H. Carotenoid Content in Different Varieties of Pumpkins. J. Food Compos. Anal. 2002, 15, 633–638. [Google Scholar] [CrossRef]
- Kinet, J.M. Environmental, Chemical, and Genetic Control of Flowering. In Horticultural Reviews; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; Volume 15. [Google Scholar]
- Amasino, M. Loss of flowering locus C Activity Eliminates the Late-Flowering Phenotype of FRIGIDA and Autonomous Pathway Mutations but Not Responsiveness to Vernalization. Plant Cell 2001, 13, 935–941. [Google Scholar]
- Goldschmidt, E.E.; Tamim, M.; Goren, R. Gibberellins and flowering in citrus and other fruit trees: A critical analysis. Acta Hortic. 1997, 463, 201–208. [Google Scholar] [CrossRef]
- Tzay-Fa, S. Effect of day and night temperature variation and of high temperature on devernalization in radish. Acta Hortic. 2000, 514, 157–162. [Google Scholar] [CrossRef]
- Sheldon, C.C.; Finnegan, E.J.; Rouse, D.T.; Tadege, M.; Dennis, E.S. The control of flowering by vernalization. Curr. Opin. Plant Biol. 2000, 3, 418–422. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed]
- Barry, C.S.; Giovannoni, J.J. Ethylene and fruit ripening. J. Plant Growth Regul. 2007, 26, 143–159. [Google Scholar] [CrossRef]
- Chervin, C.; El-Kereamy, A.; Roustan, J.P.; Latche, A.; Lamon, J.; Bouzayen, M. Ethylene seems required for the berry de-velopment and ripening in grape a non-climacteric fruit. Plant Sci. 2004, 167, 1301–1305. [Google Scholar] [CrossRef]
- Leng, F.; Cao, J.; Wang, S.; Jiang, L.; Li, X.; Sun, C. Transcriptomic Analyses of Root Restriction Effects on Phytohormone Content and Signal Transduction during Grape Berry Development and Ripening. Int. J. Mol. Sci. 2018, 19, 2300. [Google Scholar] [CrossRef]
- Kasahara, H.; Hanada, A.; Kuzuyama, T.; Takagi, M.; Kamiya, Y.; Yamaguchi, S. Contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthesis of gibberellins in Arabidopsis. J. Biol. Chem. 2002, 277, 45188–45194. [Google Scholar] [CrossRef] [PubMed]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Takahashi, A.; Masuda, T.; Noji, S. Light and abscisic acid independently regulated FaMYB10 in Fragaria × ananassa fruit. Planta 2015, 241, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.-F.; Chai, Y.-M.; Li, C.-L.; Lu, D.; Luo, J.-J.; Qin, L.; Shen, Y.-Y. Abscisic Acid Plays an Important Role in the Regulation of Strawberry Fruit Ripening. Plant Physiol. 2011, 157, 188–199. [Google Scholar] [CrossRef]
- Hou, B.-Z.; Li, C.-L.; Han, Y.-Y.; Shen, Y.-Y. Characterization of the hot pepper (Capsicum frutescens) fruit ripening regulated by ethylene and ABA. BMC Plant Biol. 2018, 18, 162. [Google Scholar] [CrossRef]
- Chanclud, E.; Morel, J.-B. Plant hormones: A fungal point of view. Mol. Plant Pathol. 2016, 17, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Gruen, H.E. Auxins and Fungi. Annu. Rev. Plant Physiol. 2003, 10, 405–440. [Google Scholar] [CrossRef]
- Smith, A.M.; Cook, R.J. Implications of ethylene production by bacteria for biological balance of soil. Nature 1974, 252, 703–705. [Google Scholar] [CrossRef]
- DaSilva, E.J.; Henriksson, E.; Henriksson, L.E. Ethylene production by fungi. Plant Sci. Lett. 1974, 2, 63–66. [Google Scholar] [CrossRef]
- Tudzynski, B.; Sharon, A. Biosynthesis, Biological Role and Application of Fungal Phytohormones. In Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar] [CrossRef]
- Chaves, F.C.; Gianfagna, T.J. Necrotrophic phase of Moniliophthora perniciosa causes salicylic acid accumulation in infected stems of cacao. Physiol. Mol. Plant Pathol. 2006, 69, 104–108. [Google Scholar] [CrossRef]
- Cooper, S.J.; Ama, F. Comparison of cytokinin and cytokinin-O-glucoside cleaving β-glucosidase production in vitro by Venturia inaequalis and other phytopathogenic fungi with differing modes of nutrition in planta. Physiol. Mol. Plant Pathol. 1998, 53, 61–72. [Google Scholar] [CrossRef]
- Arshad, M.; Frankenberger, W.T., Jr. Plant Growth-Regulating Substances in the Rhizosphere: Microbial Production and Functions. Adv. Agron. 1997, 62, 45–151. [Google Scholar]
- Ali, S.; Charles, T.C.; Glick, B.R. Endophytic Phytohormones and Their Role in Plant Growth Promotion. In Functional Importance of the Plant Microbiome; Doty, S.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 89–105. [Google Scholar]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Cohen, A.C.; Travaglia, C.N.; Bottini, R.; Piccoli, P.N. Participation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany 2009, 87, 455–462. [Google Scholar] [CrossRef]
- An, M.; Zhou, X.; Wu, F.; Ma, Y.; Yang, P. Rhizosphere soil microorganism populations and community structures of different watermelon cultivars with differing resistance to Fusarium oxysporum f. sp. niveum. Can. J. Microbiol. 2011, 57, 355. [Google Scholar] [CrossRef]
- Sun, J.; Ming, P.; Wang, Y.; Li, W.; Xia, Q. The effects of different disease-resistant cultivars of banana on rhizosphere microbial communities and enzyme activities. FEMS Microbiol. Lett. 2013, 345, 121–126. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Ameloraion of high sality stress damage by plant rowt-promofing aceral endophyes tht contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef]
- Walitang, D.I.; Kim, C.-G.; Kim, K.; Kang, Y.; Kim, Y.K.; Sa, T. The influence of host genotype and salt stress on the seed endophytic community of salt-sensitive. BMC Plant Biol. 2018, 18, 51. [Google Scholar] [CrossRef]
- Dong, M.; Yang, Z.; Cheng, G.; Peng, L.; Xu, Q.; Xu, J. Diversity of the bacterial microbiome in the roots of four Saccharum species: S. spontaneum, S. robustum, S. barberi, and S. officinarum. Front. Microbiol. 2018, 9, 267. [Google Scholar] [CrossRef]
- Niu, H.; Pang, Z.; Fallah, N.; Zhou, Y.; Zhang, C.; Hu, C.; Lin, W.X.; Yuan, Z.N. Diversity of microbial communities and soil nutrients in sugarcane rhizosphere soil under water soluble fertilizer. PLoS ONE 2021, 16, e0245626. [Google Scholar] [CrossRef] [PubMed]
- Hereira-Pacheco, S.E.; Navarro-Noya, Y.E.; Dendooven, L. The root endophytic bacterial community of Ricinus communis L. resembles the seeds community more than the rhizosphere bacteria independent of soil water content. Sci. Rep. 2021, 11, 2173. [Google Scholar] [CrossRef] [PubMed]
- Kent, A.D. Microbial communities and their interactions in soil and rhizosphere ecosystems. Annu. Rev. Microbiol. 2002, 56, 211–236. [Google Scholar] [CrossRef]
- Ma, X.; Liu, Y.; Zarebanadkouki, M.; Razavi, B.S.; Blagodatskaya, E.; Kuzyakov, Y. Spatiotemporal patterns of enzyme activities in the rhizosphere: Effects of plant growth and root morphology. Biol. Fertil. Soils 2018, 54, 819–828. [Google Scholar] [CrossRef]
- Zhu, B.; Panke-Buisse, K.; Kao-Kniffin, J. Nitrogen fertilization has minimal influence on rhizosphere effects of smooth crabgrass (Digitaria ischaemum) and bermudagrass (Cynodon dactylon). J. Plant Ecol. 2015, 8, 390–400. [Google Scholar] [CrossRef]
- Chen, Y.; Han, M.; Yuan, X.; Cao, G.; Zhu, B. Seasonal changes in soil properties, microbial biomass and enzyme activities across the soil profile in two alpine ecosystems. Soil Ecol. Lett. 2021, 3, 383–394. [Google Scholar] [CrossRef]
- Keller-Przybykowicz, S.; Rutkowski, K.P.; Kruczyńska, D.E.; Pruski, K. Changes in gene expression profile during fruit development determine fruit quality. Hortic. Sci. 2016, 43, 1–9. [Google Scholar] [CrossRef]
- Dal Peruffo, A.B.; Pallavicini, C. Enzymatic changes associated with ripening of grape berries. J. Sci. Food Agric. 1975, 26, 559–566. [Google Scholar] [CrossRef]
- Leo, P.D.; Sacher, J.A. Senescense: Association of synthesis of Acid phosphatase with banana ripening. Plant Physiol. 1970, 46, 208–211. [Google Scholar] [CrossRef]
- Guerrero-zuniga, A.L.; Lopez-lopeze, E.; Rodriguez-tovar, A.V.; Rodriguez-dorantes, A. Functional diversity of plant endophytes and their role in assisted phyftoremediation. Bioremediat. Ind. Waste Environ. Saf. 2020, 237–255. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Zhao, J.R.; Li, Y.F.; Wang, Y.D.; Xing, J.F.; Zhang, H.S.; Zhang, C.Y. Breeding and Cultivation Techniques of New Maize Variety Jiyu 5817 with High Quality and Yield and Mid-early Maturing. J. Hebei Agric. Sci. 2015, 5, 10–14. [Google Scholar]
- Olasantan, F.O. Effect of population density and sowing date of pumpkin on soil hydrothermal regime, weed control and crop growth in a yam-pumpkin intercrop. Exp. Agric. 2007, 43, 365–380. [Google Scholar] [CrossRef]
- Astapovich, S.P. Influence of Meteorological Conditions on Growth, Development and Yield of Winter Rape at Various Terms of Sowing in the Conditions of Belarusian Polesye. Arable Farm. Breed. Belarus Collect. Sci. Pap. 2009, 45, 193–202. [Google Scholar]
- Keinath, A.P.; Dubose, V.B. Evaluation of Pumpkin Cultivars for Powdery and Downy Mildew Resistance, Virus Tolerance, and Yield. HortScience 2000, 35, 281–285. [Google Scholar] [CrossRef]
- Aroi Ee, H.; Omidbaigi, R. Effects of nitrogen fertilizer on productivity of medicinal pumpkin. Acta Hortic. 2004, 629, 415–419. [Google Scholar] [CrossRef]
- Riley, D.; Barber, S.A. Salt Accumulation at the Soybean (Glycine Max. (L.) Merr.) Root-Soil Interface1. Soil Sci. Soc. Am. J. 1970, 34, 154–155. [Google Scholar] [CrossRef]
- Hayano, K. A method for the determination of β-glucosidase activity in soil. Soil Sci. Plant Nutr. 1973, 19, 103–108. [Google Scholar] [CrossRef]
- Ladd, J.N. Properties of proteolytic enzymes extracted from soil. Soil Biol. Biochem. 1972, 4, 227–237. [Google Scholar] [CrossRef]
- Yang, S.D.; Xiao, J.; Liang, T.; He, W.Z.; Tan, H.W. Response of soil biological properties and bacterial diversity to different levels of nitrogen application in sugarcane fields. AMB Express. 2021, 11, 172. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Feng, J.; Chen, S.; Zhao, Z.; Li, B.; Wang, Y.; Jia, J.; Li, S.; Wang, Y.; Yan, M.; et al. Geographical Patterns of nirS Gene Abundance and nirS-Type Denitrifying Bacterial Community Associated with Activated Sludge from Different Wastewater Treatment Plants. Microb. Ecol. 2018, 77, 304–316. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Chen, S.Y.; Sun, Y.; Yang, S.D.; He, Y. Differences of rhizospheric and endophytic bacteria are recruited by different watermelon phenotypes relating to rind colors formation. Sci. Rep. 2022, 2022. 12, 6360. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.; Bakker, P. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Okunishi, S.; Sako, K.; Mano, H.; Imamura, A.; Morisaki, H. Bacterial Flora of Endophytes in the Maturing Seed of Cultivated Rice (Oryza sativa). Microbes Environ. 2005, 20, 168–177. [Google Scholar] [CrossRef]
- Li, X.L.; Li, Q.; Peng, W.H.; Gan, B.C.; Zheng, L.Y.; Zhang, X.P. Study on diversity and community structure of bacteria in soils at tobacco maturing stage. Acta Tab. Sin. 2015, 21, 40–47. [Google Scholar]
- Knight, T.R.; Dick, R.P. Differentiating microbial and stabilized β-glucosidase activity relative to soil quality. Soil Biol. Biochem. 2004, 36, 2089–2096. [Google Scholar] [CrossRef]
- Harrison, A.F. Relationship between intensity of phosphatase activity and physico-chemical properties in woodland soils. Soil Biol. Biochem. 1983, 15, 93–99. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Sanaullah, M.; Blagodatskaya, E.; Chabbi, A.; Rumpel, C.; Kuzyakov, Y. Drought effects on microbial biomass and enzyme activities in the rhizosphere of grasses depend on plant community composition. Appl. Soil Ecol. A Sect. Agric. Ecosyst. Environ. 2011, 48, 38–44. [Google Scholar] [CrossRef]
- Chen, P.; Pei, Y.; Liang, B.; Zhang, Y.; Zhai, X.; He, S.; Kai, W.; Sun, Y.; Leng, P. Role of abscisic acid in regulating fruit set and ripening in squash (Cucurbita pepo L.). N. Z. J. Crop Hortic. Sci. 2016, 44, 274–290. [Google Scholar] [CrossRef]
- Li, J.; Tao, X.; Li, L.; Mao, L.; Luo, Z.; Khan, Z.U.; Ying, T. Comprehensive RNA-Seq analysis on the regulation of tomato ripening by exogenous auxin. PLoS ONE 2016, 11, e0156453. [Google Scholar] [CrossRef] [PubMed]
- Ziliotto, F.; Corso, M.; Rizzini, F.M.; Rasori, A.; Botton, A.; Bonghi, C. Grape berry ripening delay induced by a pre-veraison NAA treatment is paralleled by a shift in the expression pattern of auxin-and ethylene-related genes. BMC Plant Biol. 2012, 12, 185. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Mei, Y.; Li, Q.; Ma, H.; Wu, Y.; Yang, Z. Effects of End of Day Far-Red Light on Growth, Histiocyte Morphology and Phytohormones Content of Pumpkin Seedlings. Sci. Agric. Sin. 2020, 53, 4248–4258. [Google Scholar] [CrossRef]
- Martínez, C.; Manzano, S.; Megías, Z.; Garrido, D.; Picó, B.; Jamilena, M. Involvement of ethylene biosynthesis and signalling in fruit set and early fruit development in zucchini squash (Cucurbita pepo L.). BMC Plant Biol. 2013, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Q. Effects of Microbial Community on Crop Growth and Phytohormones; Chinese Academy of Agricultural Sciences: Beijing, China, 2019. [Google Scholar]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Leveau, H.J.; Gerards, S. Discovery of a bacterial gene cluster for catabolism of the plant hormone indole 3-acetic acid. FEMS Microbiol. Ecol. 2008, 65, 238–250. [Google Scholar] [CrossRef]
- Weingart, H.; Vlksch, B.; Ullrich, M.S. Comparison of Ethylene Production by Pseudomonas syringae and Ralstonia solanacearum. Phytopathology 1999, 89, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhu, L.; Xie, Y.; Li, F.; Xiao, X.; Ma, Z.; Wang, J. Bacillus licheniformis SA03 Confers Increased Saline–Alkaline Tolerance in Chrysanthemum Plants by Induction of Abscisic Acid Accumulation. Front. Plant Sci. 2017, 8, 1143. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Radhakrishnan, R.; Kang, S.-M.; You, Y.-H.; Jeong, E.-J.; Kim, J.-G.; Lee, I.-J. Plant growth promoting effect of Bacillus amyloliquefaciens H-2-5 on crop plants and influence on physiological changes in soybean under soil salinity. Physiol. Mol. Biol. Plants 2017, 23, 571–580. [Google Scholar] [CrossRef]
- Vanissa, T.T.G.; Berger, B.; Patz, S.; Becker, M.; Turečková, V.; Novák, O.; Tarkowská, D.; Henri, F.; Ruppel, S. The Response of Maize to Inoculation with Arthrobacter sp. and Bacillus sp. in Phosphorus-Deficient, Salinity-Affected Soil. Microorganisms 2020, 8, 1005. [Google Scholar] [CrossRef]
- Tchuisseu Tchakounté, G.V.; Berger, B.; Patz, S.; Fankem, H.; Ruppel, S. Community structure and plant growth-promoting potential of cultivable bacteria isolated from Cameroon soil. Microbiol. Res. 2018, 214, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Antoun, H.; Beauchamp, C.; Goussard, N.; Chabot, R.; Lalande, R. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effect on radishes (Raphanus sativus L.). Plant Soil 1998, 204, 57–67. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequencing Type | Primer Name | Primer Sequence | Length | Sequencing Platform |
|---|---|---|---|---|
| Rhizospheric bacterial | 338F | 5′-ACTCCTACGGGAGGCAGCAG-3′ | 311 bp | MiSeq PE300 |
| 806R | 5′-GGACTACHVGGGTWTCTAAT-3′ | |||
| Endophytic bacterial | 799F | 5′-AACMGGATTAGATACCCKG-3′ | 394 bp | MiSeq PE250 |
| 1193R | 5′-ACGTCATCCCCACCTTCC-3′ |
| Sample | β-Glucosidase | Phosphatase | Aminopeptidase |
|---|---|---|---|
| EM | 0.27 ± 0.13 a | 2.83 ± 1.11 a | 14.18 ± 1.99 a |
| LM | 2.44 ± 0.69 a | 2.44 ± 0.69 a | 10.81 ± 2.47 b |
| CK | 0.12 ± 0.06 b | 1.14 ± 0.26 b | 9.25 ± 0.67 b |
| Shannon | Invsimpson | Ace | Chao1 | Heip | Coverage | |
|---|---|---|---|---|---|---|
| EM | 6.78 ± 0.17 a | 312.00 ± 61.16 a | 4807.82 ± 464.85 a | 4782.72 ± 433.63 a | 0.26 ± 0.02 a | 0.97 |
| LM | 6.71 ± 0.15 a | 286.85 ± 64.78 a | 4711.95 ± 134.07 a | 4653.41 ± 102.38 a | 0.25 ± 0.03 a | 0.97 |
| CK | 6.36 ± 0.12 b | 180.15 ± 26.42 b | 4058.39 ± 604.8 a | 4036.71 ± 520.56 a | 0.19 ± 0.01 b | 0.98 |
| Shannon | Invsimpson | Ace | Chao1 | Heip | Coverage | |
|---|---|---|---|---|---|---|
| EM | 4.27 ± 0.27 a | 31.96 ± 12.69 a | 455.7 ± 92.83 a | 453.5 ± 90.16 a | 0.2 ± 0.06 a | 0.99 |
| LM | 3.95 ± 1.00 a | 30.07 ± 15.91 a | 479.5 ± 86.69 a | 478.73 ± 92.24 a | 0.17 ± 0.07 a | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Qin, R.; Yang, D.; Liu, W.; Yang, S. A Comparison of Rhizospheric and Endophytic Bacteria in Early and Late-Maturing Pumpkin Varieties. Microorganisms 2022, 10, 1667. https://doi.org/10.3390/microorganisms10081667
Chen S, Qin R, Yang D, Liu W, Yang S. A Comparison of Rhizospheric and Endophytic Bacteria in Early and Late-Maturing Pumpkin Varieties. Microorganisms. 2022; 10(8):1667. https://doi.org/10.3390/microorganisms10081667
Chicago/Turabian StyleChen, Siyu, Renliu Qin, Da Yang, Wenjun Liu, and Shangdong Yang. 2022. "A Comparison of Rhizospheric and Endophytic Bacteria in Early and Late-Maturing Pumpkin Varieties" Microorganisms 10, no. 8: 1667. https://doi.org/10.3390/microorganisms10081667