
Sediment Bacteria and Phosphorus Fraction Response, Notably to Titanium Dioxide Nanoparticle Exposure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, TiO2 NPs Suspension, and Expose Experiment
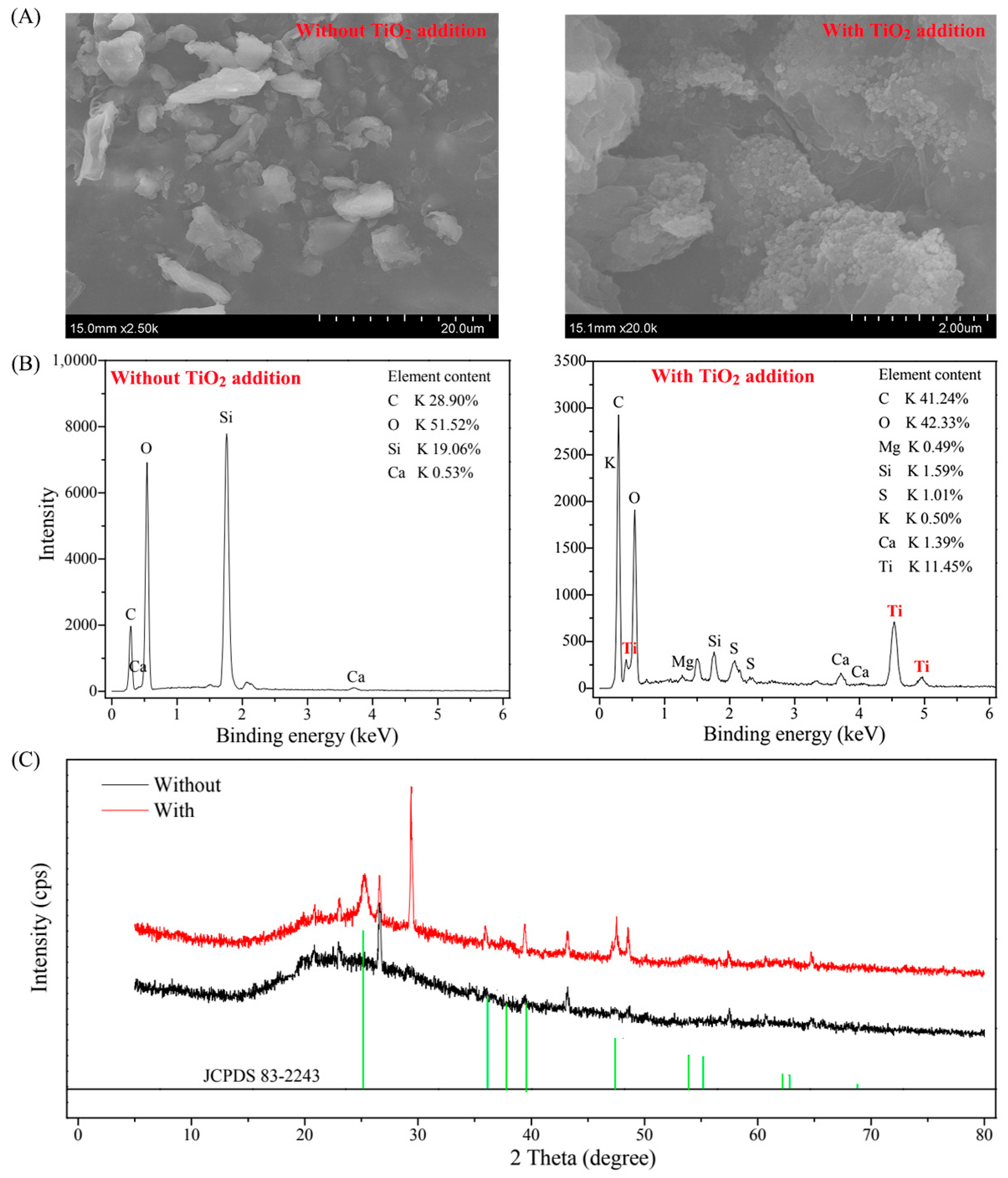
2.2. Detection of TiO2 NPs
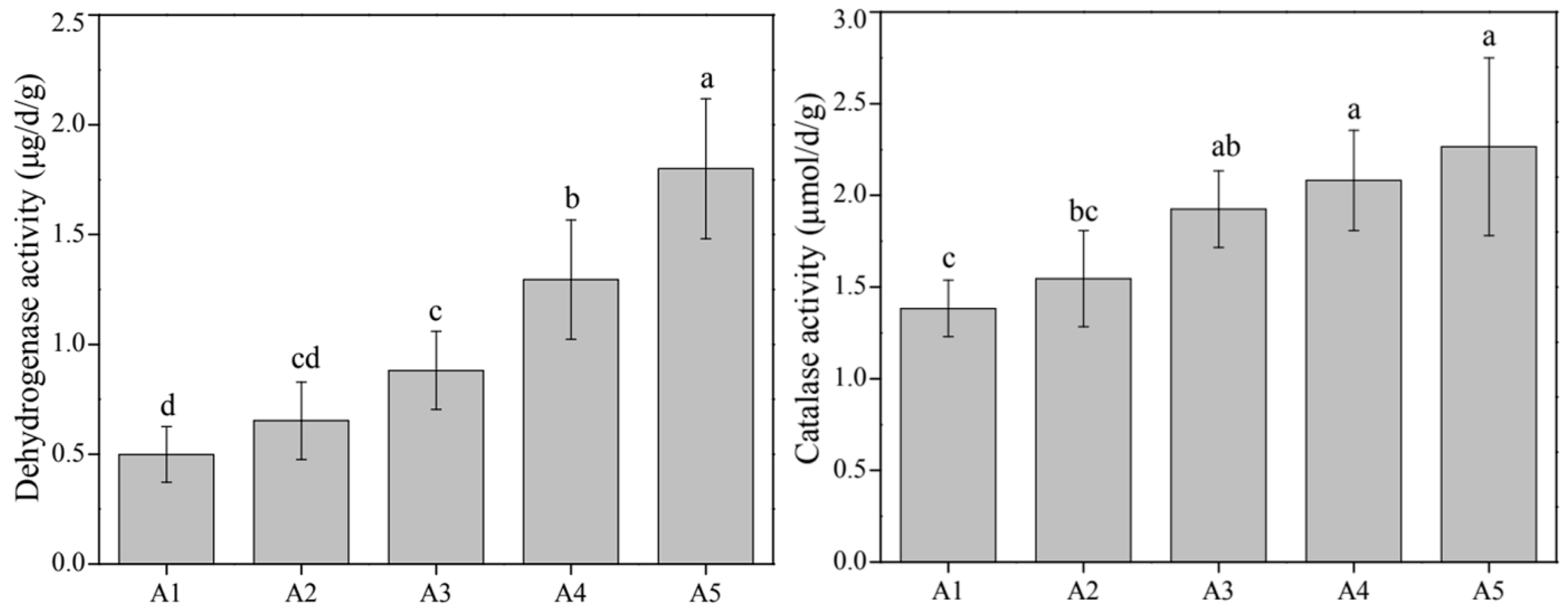
2.3. Determination of Phosphorus Fractions and Sediment Enzyme Assay
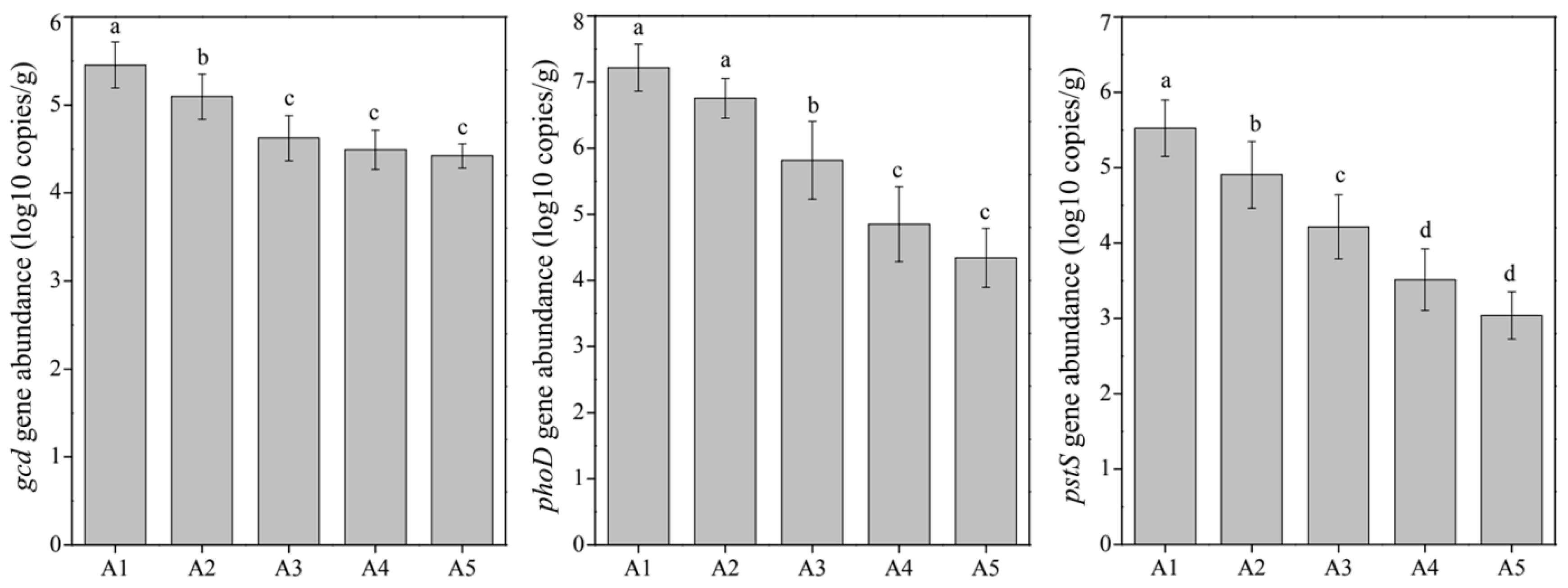
2.4. DNA Extraction, Gene Quantification, MiSeq Sequencing, and Data Processing
2.5. Data Analysis
3. Results and Discussion
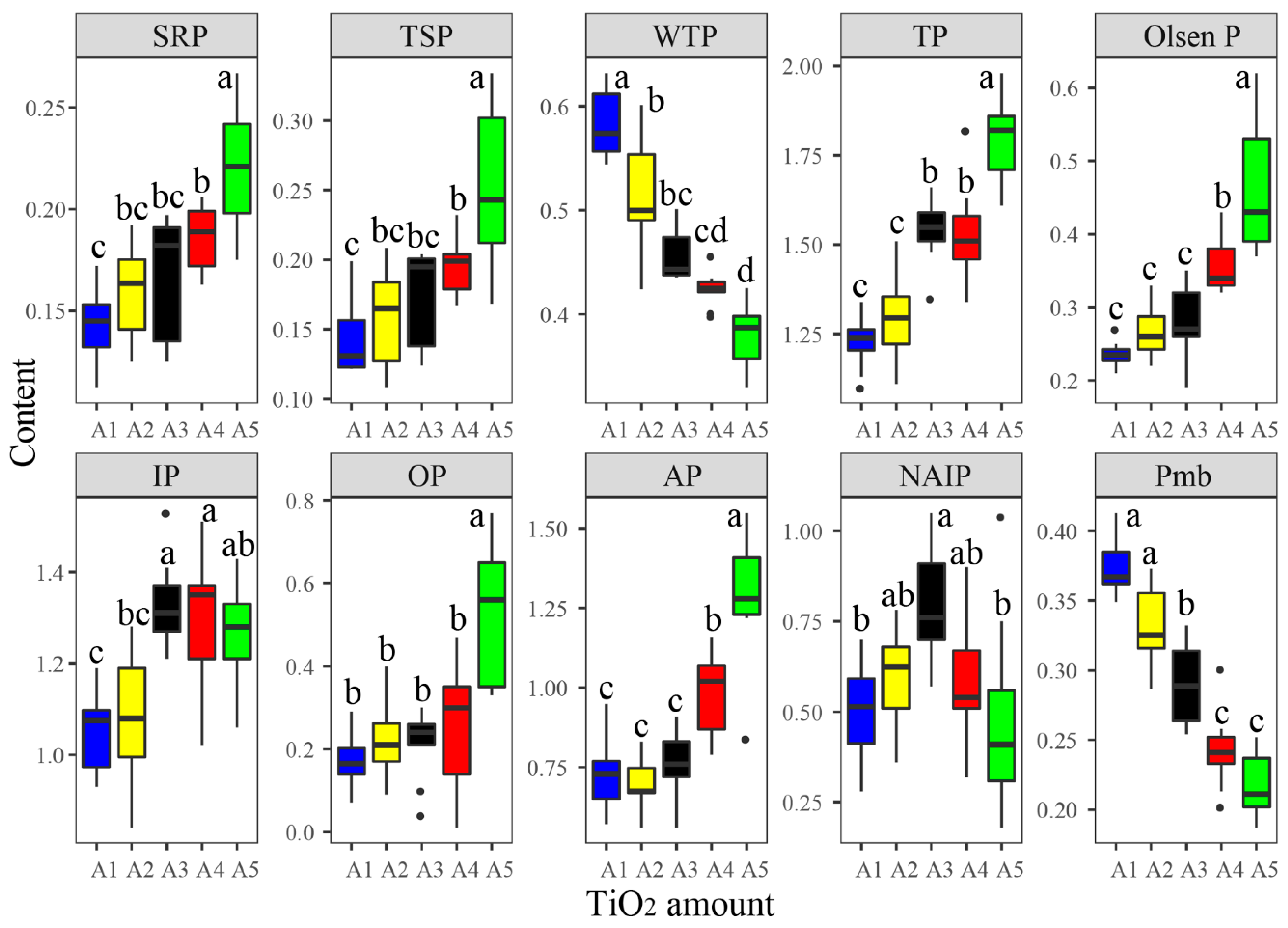
3.1. Responses of Phosphorus Fractions to TiO2 NP Addition
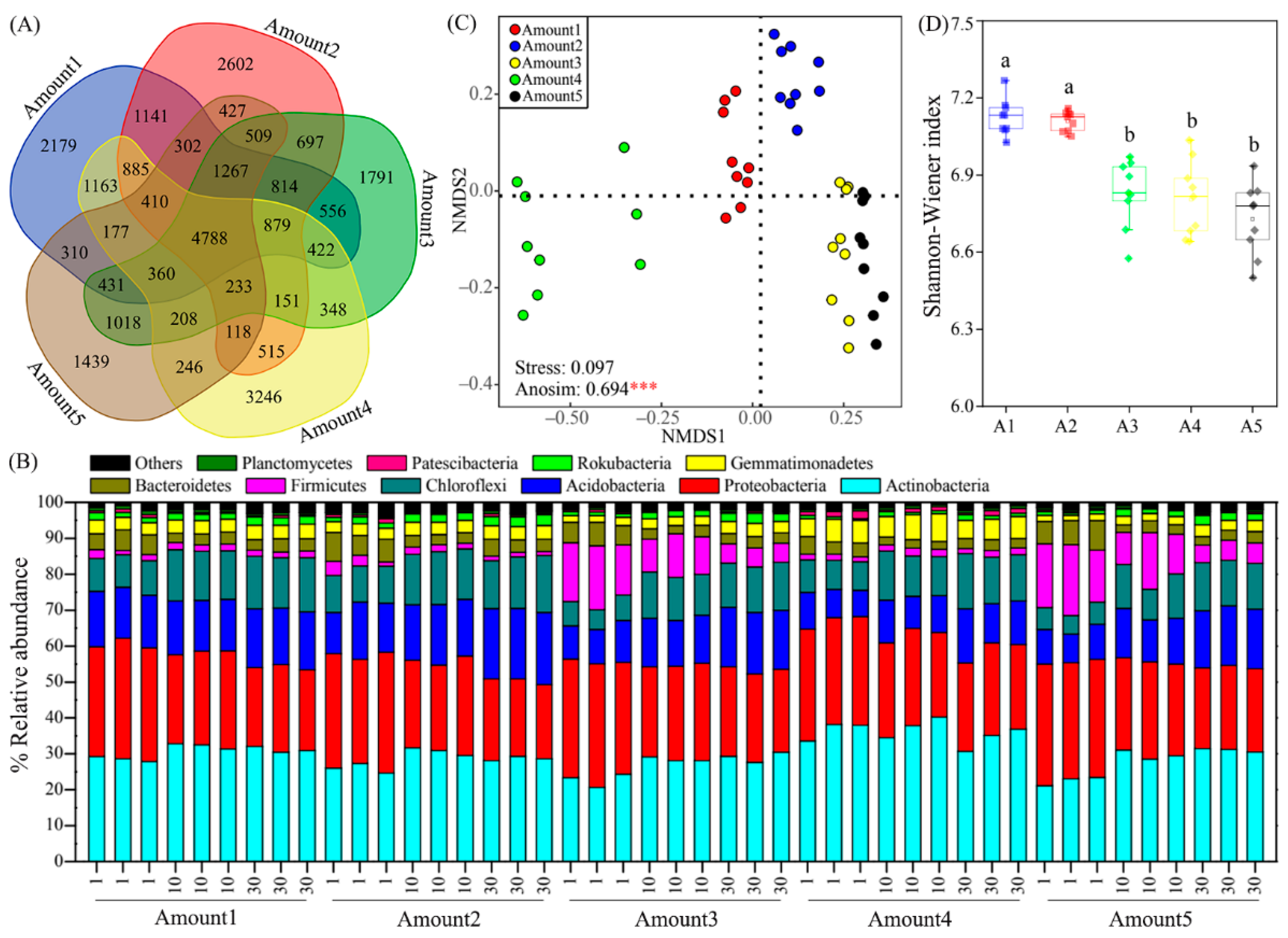
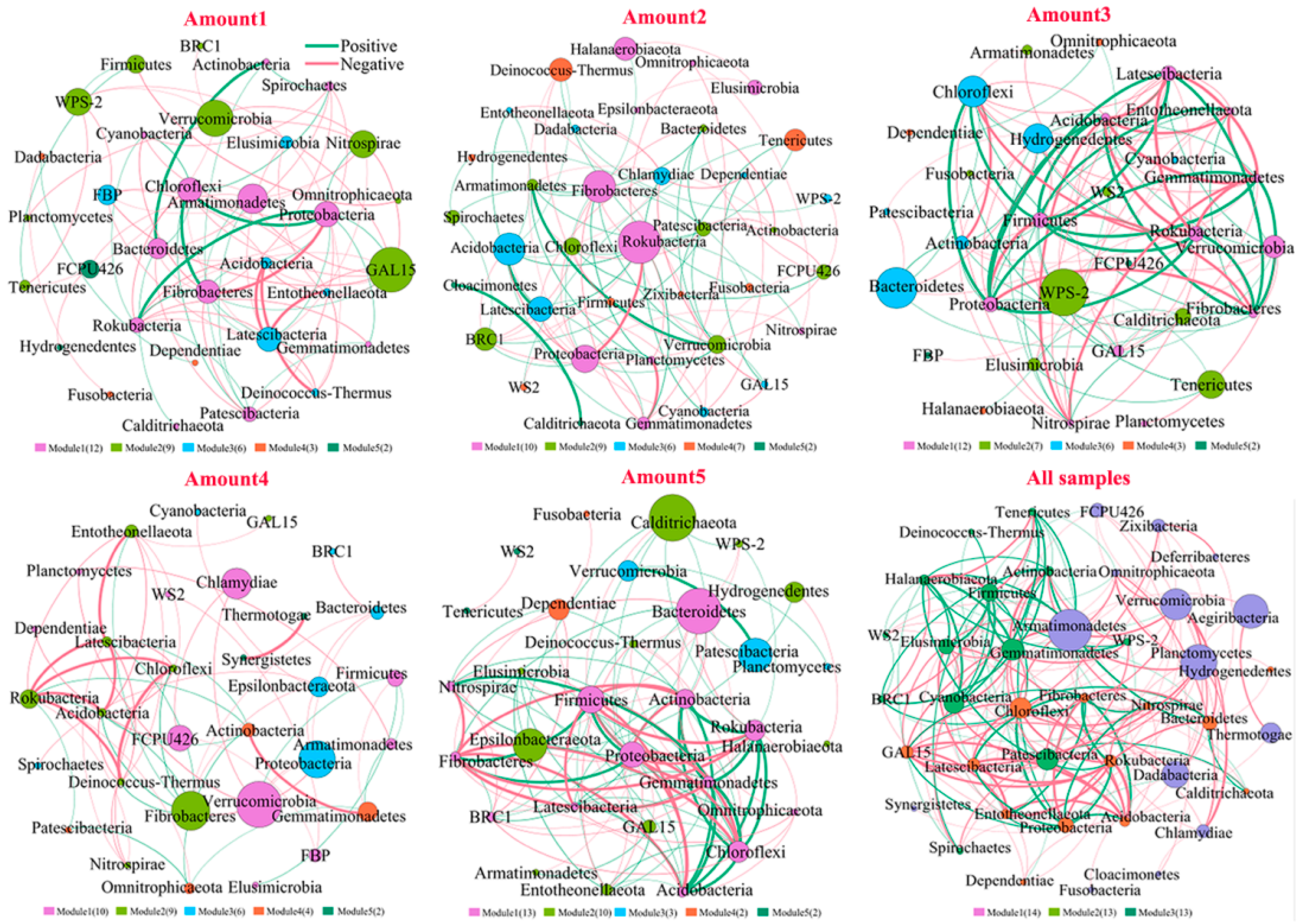
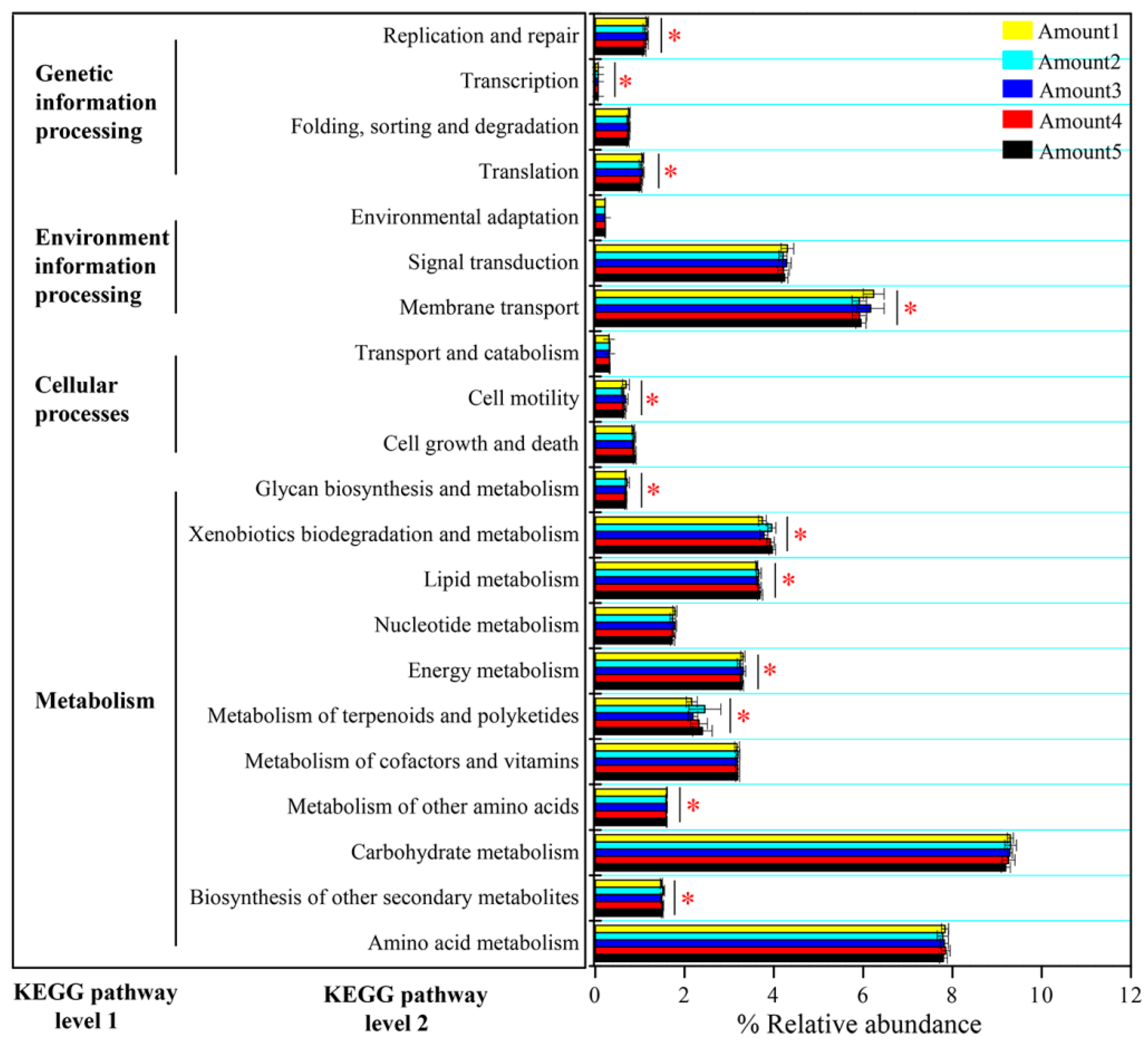
3.2. Responses of the Bacterial Community to TiO2 NP Addition
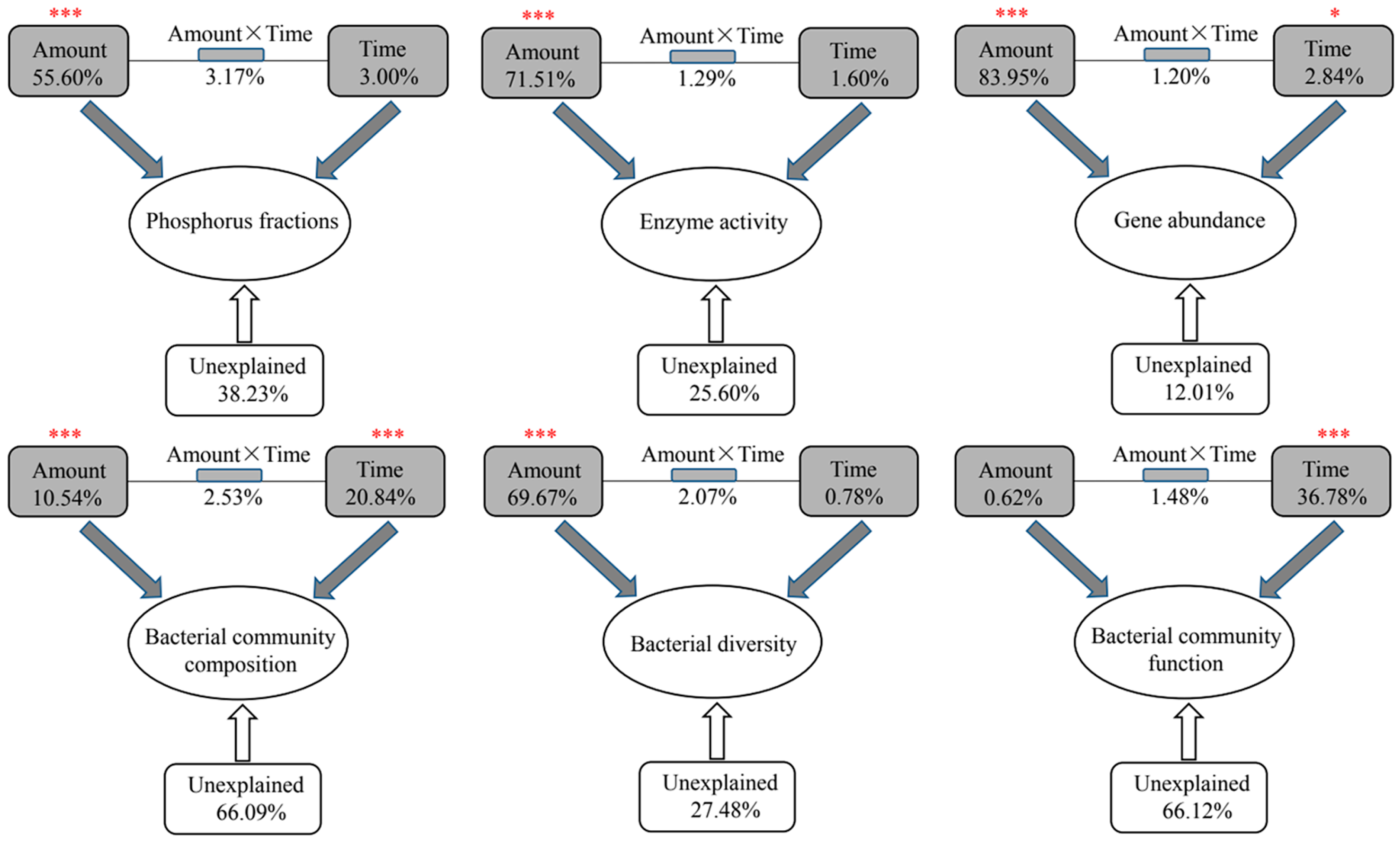
3.3. Potential Effects of Abiotic and Biotic Factors on Phosphorus Fractionation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, C.G.; Javed, H.; Zhang, D.; Kim, J.H.; Westerhoff, P.; Li, Q.; Alvarez, P.J.J. Porous Electrospun Fibers Embedding TiO2 for Adsorption and Photocatalytic Degradation of Water Pollutants. Environ. Sci. Technol. 2018, 52, 4285–4293. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, C.G.; Kang, J.K.; Park, S.J.; Alvarez, P. Simple preparation method for Styrofoam-TiO2 composites and their photocatalytic application for dye oxidation and Cr(VI) reduction in industrial wastewater. Environ. Sci. Wat. Res. 2021, 7, 222–230. [Google Scholar] [CrossRef]
- Yang, X.; Chen, Y.; Guo, F.; Liu, X.; Su, X.; He, Q. Metagenomic analysis of the biotoxicity of titanium dioxide nanoparticles to microbial nitrogen transformation in constructed wetlands. J. Hazard. Mater. 2020, 384, 12137. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, W.; Li, Y.; Li, Y.; Ye, B.; Tang, M.; Wang, D. Small sizes of TiO2-NPs exhibit adverse effects at predicted environmental relevant concentrations on nematodes in a modified chronic toxicity assay system. J. Hazard. Mater. 2012, 243, 161–168. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, C.; Cota-Ruiz, K.; Tan, W.; Reyes, A.; Peralta-Videa, J.R.; Hernandez-Viezcas, J.A.; Li, C.; Gardea-Torresdey, J.L. Effects of different surface-coated nTiO2 on full-grown carrot plants: Impacts on root splitting, essential elements, and Ti uptake. J. Hazard. Mater. 2021, 402, 123768. [Google Scholar] [CrossRef]
- Madhubala, V.; Pugazhendhi, A.; Thirunavukarasu, K. Cytotoxic and immunomodulatory effects of the low concentration of titanium dioxide nanoparticles (TiO2 NPs) on human cell lines—An in vitro study. Proces. Biochem. 2019, 86, 186–195. [Google Scholar] [CrossRef]
- Arrigo, K.R. Marine microorganisms and global nutrient cycles. Science 2005, 437, 349–355. [Google Scholar] [CrossRef]
- Ma, W.; Li, H.; Zhang, W.; Shen, C.; Wang, L.; Li, Y.; Li, Q.; Wang, Y. TiO2 nanoparticles accelerate methanogenesis in mangrove wetlands sediment. Sci. Total Environ. 2020, 713, 136602. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, Z.; Li, Z.Q.; Pan, Q.K.; Yan, C.Z. Effects of titania nanoparticles on phosphorus fractions and its release in resuspended sediments under UV. J. Hazard. Mater. 2010, 174, 477–483. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Zhuo, T.; Ding, Y.; Wan, Q.; Li, S.; Chai, B.; Lei, X. Effects of hydrostatic pressure on phosphorus transformation at the water-sediment interface of a deep reservoir: Novel insights into bacterial community and functional genes. J. Soil Sediment. 2021, 21, 3367–3379. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, Z.; Wei, Q.; Yan, C.; Liu, F. Effects of engineered nano-titanium dioxide on pore surface properties and phosphorus adsorption of sediment: Its environmental implications. J. Hazard. Mater. 2011, 192, 1364–1369. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Xu, N.; Christodoulatos, C.; Wang, D. Synergistic effects of phosphorus and humic acid on the transport of anatase titanium dioxide nanoparticles in water-saturated porous media. Environ. Pollut. 2018, 243, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Nishio, K.; Fujita, K.; Endoh, S.; Miyauchi, A.; Saito, Y.; Iwahashi, H.; Yamamoto, K.; Murayama, H.; Nakano, H.; et al. Protein adsorption of ultrafine metal oxide and its influence on cytotoxicity toward cultured cells. Chem. Res. Toxicol. 2009, 22, 543–553. [Google Scholar] [CrossRef]
- Wu, D.; Yang, S.; Du, W.; Yin, Y.; Zhang, J.; Guo, H. Effects of titanium dioxide nanoparticles on Microcystis aeruginosa and microcystins production and release. J. Hazard. Mater. 2019, 377, 1–7. [Google Scholar] [CrossRef]
- Xu, K.; Li, Z.; Juneau, P.; Xiao, F.; Lian, Y.; Zhang, W.; Shu, L.; Jiang, H.; Zhang, K.; Wang, C.; et al. Toxic and protective mechanisms of cyanobacterium Synechocystis sp. in response to titanium dioxide nanoparticles. Environ. Pollut. 2021, 274, 116508. [Google Scholar] [CrossRef]
- Li, K.; Qian, J.; Wang, P.; Wang, C.; Lu, B.; Jin, W.; He, X.; Tang, S.; Zhang, C.; Gao, P. Effects of aging and transformation of anatase and rutile TiO2 nanoparticles on biological phosphorus removal in sequencing batch reactors and related toxic mechanisms. J. Hazard. Mater. 2020, 398, 123030. [Google Scholar] [CrossRef]
- Li, L.; Li, B.; Wang, Q.; Hou, N.; Li, C.; Cheng, X. Toxicity of TiO2 nanoparticle to denitrifying strain CFY1 and the impact on microbial community structures in activated sludge. Chemosphere 2016, 144, 1334–1341. [Google Scholar] [CrossRef]
- Qiu, T.A.; Meyer, B.M.; Christenson, K.G.; Klaper, R.D.; Haynes, C.L. A mechanistic study of TiO2 nanoparticle toxicity on Shewanella oneidensis MR-1 with UV-containing simulated solar irradiation: Bacterial growth, riboflavin secretion, and gene expression. Chemosphere 2017, 168, 1158–1168. [Google Scholar] [CrossRef]
- Zhai, Y.; Hunting, E.R.; Liu, G.; Baas, E.; Peijnenburg, W.J.G.M.; Vijver, M.G. Compositional alterations in soil bacterial communities exposed to TiO2 nanoparticles are not reflected in functional impacts. Environ. Res. 2019, 178, 108713. [Google Scholar] [CrossRef]
- APAH. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Wan, W.; Li, X.; Han, S.; Wang, L.; Luo, X.; Chen, W.; Huang, Q. Soil aggregate fractionation and phosphorus fraction driven by long-term fertilization regimes affect the abundance and composition of P-cycling-related bacteria. Soil Till. Res. 2020, 196, 104475. [Google Scholar] [CrossRef]
- Wan, W.; Wang, Y.; Tan, J.; Qin, Y.; Zuo, W.; Wu, H.; He, H.; He, D. Alkaline phosphatase-harboring bacterial community and multiple enzyme activity contribute to phosphorus transformation during vegetable waste and chicken manure composting. Bioresour. Technol. 2020, 297, 122406. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.I.; Temple, K.L. Some variables affecting the measurement of catalase activity in soil. Soil Sci. Soc. Am. Pro. 1964, 28, 207–216. [Google Scholar] [CrossRef]
- Öhlinger, R. Dehydrogenase activity with the substrate TTC. In Methods in Soil Biology; Schinner, F., Ohlinger, R., Kandler, E., Margesin, R., Eds.; Springer: Berlin, Germany, 1996; pp. 241–243. [Google Scholar]
- Santos, S.S.; Nielsen, T.K.; Hansen, L.; Winding, A. Comparison of three DNA extraction methods for recovery of soil protist DNA. J. Microbiol. Meth. 2015, 115, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and experimental application of a novel non-degenerate universal primer set that amplifies prokaryotic 16S rRNA genes with a low possibility to amplify eukaryotic rRNA genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Zhang, Y.; Cheng, G.; Li, X.; Qin, Y.; He, D. Dredging mitigates cyanobacterial bloom in eutrophic Lake Nanhu: Shifts in associations between the bacterioplankton community and sediment biogeochemistry. Environ. Res. 2020, 188, 109799. [Google Scholar] [CrossRef]
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: A R-based tool for the rapid prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene marker gene sequences. Environ. Microbiome. 2020, 15, 11. [Google Scholar] [CrossRef]
- Xiong, W.; Tang, Y.; Shao, C.; Zhao, Y.; Jin, B.; Huang, T.; Miao, Y.N.; Shu, L.; Ma, W.; Xu, X.; et al. Prevention of cyanobacterial blooms using nanosilica: A biomineralization-inspired strategy. Environ. Sci. Technol. 2017, 51, 12717–12726. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, S.; Li, D. Decomposition of cyanobacterial bloom contributes to the formation and distribution of iron-bound phosphorus (Fe-P): Insight for cycling mechanism of internal phosphorus loading. Sci. Total Environ. 2019, 652, 696–708. [Google Scholar] [CrossRef]
- Horie, M.; Tabei, Y. Role of oxidative stress in nanoparticles toxicity. Free Radical Res. 2020, 55, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Gong, N.; Shao, K.; Che, C.; Sun, Y. Stability of nickel oxide nanoparticles and its influence on toxicity to marine algae Chlorella vulgaris. Mar. Pollut. Bull. 2019, 149, 110532. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Zhang, L.; Yang, K.; Zhu, L.; Lin, D. Toxicity of iron-based nanoparticles to green algae: Effects of particle size, crystal phase, oxidation state and environmental aging. Environ. Pollut. 2016, 218, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Xiao, Z.; Yue, L.; Wang, J.; Feng, Y.; Zhu, X.; Wang, Z.; Xing, B. Algae response to engineered nanoparticles: Current understanding, mechanisms and implications. Environ. Sci. Nano 2019, 6, 1026–1042. [Google Scholar] [CrossRef]
- Gebicka, L.; Krych-Madej, J. The role of catalases in the prevention/promotion of oxidative stress. J. Inorg. Biochem. 2019, 197, 110699. [Google Scholar] [CrossRef]
- González, J.; Castillo, R.; García-Campos, M.A.; Noriega-Samaniego, D.; Escobar-Sánchez, V.; Romero-Aguilar, L.; Alba-Lois, L.; Segal-Kischinevzky, C. Tolerance to oxidative stress in budding yeast by heterologous expression of catalases A and T from Debaryomyces hansenii. Curr. Microbiol. 2020, 77, 4000–4015. [Google Scholar] [CrossRef]
- Jovanović, B.; Milosević, D.; Piperac, M.S.; Savić, A. In situ effects of titanium dioxide nanoparticles on community structure of freshwater benthic macroinvertebrates. Environ. Pollut. 2016, 213, 278–282. [Google Scholar] [CrossRef]
- Zhai, Y.; Chen, L.; Liu, G.; Song, L.; Arenas-Lago, D.; Kong, L.; Peijnenburg, W.; Vijver, M.G. Compositional and functional responses of bacterial community to titanium dioxide nanoparticles varied with soil heterogeneity and exposure duration. Sci. Total Environ. 2021, 773, 144895. [Google Scholar] [CrossRef]
- Asadishad, B.; Chahal, S.; Akbari, A.; Cianciarelli, V.; Azodi, M.; Ghoshal, S.; Tufenkji, N. Amendment of agricultural soil with metal nanoparticles: Effects on soil enzyme activity and microbial community composition. Environ. Sci. Technol. 2018, 52, 1908–1918. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, M.; Zhang, W.; Gardea-Torresdey, J.L.; White, J.C.; Ji, R.; Zhao, L. Silver nanoparticles alter soil microbial community compositions and metabolite profiles in unplanted and cucumber-planted soil. Environ. Sci. Technol. 2020, 54, 3334–3342. [Google Scholar] [CrossRef]
- Hou, J.; Li, T.; Miao, L.; You, G.; Xu, Y.; Liu, S. Effects of titanium dioxide nanoparticles on algal and bacterial communities in periphytic biofilms. Environ. Pollut. 2019, 251, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Avilés, P.; Caretta, C.A.; Brito, E.M.S.; Bertin, P.; Cuevas-Rodríguez, G.; Duran, R. Changes in bacterial diversity of activated sludge exposed to titanium dioxide nanoparticles. Biodegradation 2021, 32, 313–326. [Google Scholar] [CrossRef]
- Jiao, S.; Lu, Y. Abundant fungi adapt to broader environmental gradients than rare fungi in agricultural fields. Glob. Change Biol. 2020, 26, 4506–4520. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Grossart, H.P.; He, D.; Yuan, W.; Yang, Y. Stronger environmental adaptation of rare rather than abundant bacterioplankton in response to dredging in eutrophic Lake Nanhu (Wuhan, China). Water Res. 2021, 190, 116751. [Google Scholar] [CrossRef] [PubMed]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Env. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef]
- Hou, J.; Li, T.; Miao, L.; You, G.; Xu, Y.; Liu, S. Dynamic responses of community structure and microbial functions of periphytic biofilms during chronic exposure to TiO2 NPs. Environ. Sci. Nano 2020, 7, 665–675. [Google Scholar] [CrossRef]
- Simonin, M.; Richaume, A.; Guyonnet, J.P.; Dubost, A.; Martins, J.M.F.; Pommier, T. Titanium dioxide nanoparticles strongly impact soil microbial function by affecting archaeal nitrifiers. Sci. Rep. 2016, 6, 33643. [Google Scholar] [CrossRef]
- Wan, W.; Hao, X.; Xing, Y.; Liu, S.; Zhang, X.; Li, X.; Chen, W.; Huang, Q. Spatial differences in soil microbial diversity caused by pH-driven organic phosphorus mineralization. Land Degrad. Dev. 2021, 32, 766–776. [Google Scholar] [CrossRef]
- Qin, M.; Wang, X.; Jiang, L.; Wu, N.; Zhou, W. Bacterial diversity in phosphorus immobilization of the South China Sea. Environ. Technol. 2019, 41, 2844–2853. [Google Scholar] [CrossRef]
- Cleton-Jansen, A.M.; Goosen, N.; Fayet, O.; van de Putte, P. Cloning, mapping, and sequencing of the gene encoding Escherichia coli quinoprotein glucose dehydrogenase. J. Bacteriol. 1990, 172, 6308–6315. [Google Scholar] [CrossRef]
- Hsieh, Y.J.; Wanner, B.L. Global regulation by the seven-component pi signaling system. Curr. Opin. Microbiol. 2010, 13, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, M.; Wasaki, J.; Tomizawa, Y.; Shinano, T.; Osaki, M. Analysis of bacterial communities on alkaline phosphatase genes in soil supplied with organic matter. Soil. Sci. Plant Nutr. 2008, 54, 62–71. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Dehydrogenase | Catalase | gcd | phoD | pstS | Diversity | Composition |
|---|---|---|---|---|---|---|---|
| SRP | 0.719 *** | 0.351 * | −0.679 *** | −0.776 *** | −0.777 *** | −0.391 ** | 7.52% ** |
| TSP | 0.716 *** | 0.426 ** | −0.640 *** | −0.731 *** | −0.726 *** | −0.346 * | 7.21% ** |
| WTP | −0.786 *** | −0.754 *** | 0.801 *** | 0.792 *** | 0.817 *** | 0.684 ** | 8.46% *** |
| TP | 0.778 *** | 0.803 *** | −0.743 *** | −0.779 *** | −0.788 *** | −0.763 *** | 11.07% *** |
| Olsen P | 0.690 *** | 0.580 *** | −0.604 *** | −0.703 *** | −0.709 *** | −0.515 *** | 5.75% * |
| IP | 0.476 *** | 0.616 *** | −0.661 *** | −0.496 *** | −0.507 *** | −0.545 *** | 6.27% * |
| OP | 0.582 *** | 0.481 *** | −0.357 * | −0.564 *** | −0.561 *** | −0.495 *** | 6.26% * |
| AP | 0.802 *** | 0.529 *** | −0.543 *** | −0.543 *** | −0.699 *** | −0.503 *** | 5.38% * |
| NAIP | −0.165 | 0.213 | −0.128 | −0.128 | 0.030 | −0.202 | 2.56% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piao, S.; He, D. Sediment Bacteria and Phosphorus Fraction Response, Notably to Titanium Dioxide Nanoparticle Exposure. Microorganisms 2022, 10, 1643. https://doi.org/10.3390/microorganisms10081643
Piao S, He D. Sediment Bacteria and Phosphorus Fraction Response, Notably to Titanium Dioxide Nanoparticle Exposure. Microorganisms. 2022; 10(8):1643. https://doi.org/10.3390/microorganisms10081643
Chicago/Turabian StylePiao, Sixuan, and Donglan He. 2022. "Sediment Bacteria and Phosphorus Fraction Response, Notably to Titanium Dioxide Nanoparticle Exposure" Microorganisms 10, no. 8: 1643. https://doi.org/10.3390/microorganisms10081643