
Potential for Biological Control of Pythium schmitthenneri Root Rot Disease of Olive Trees (Olea europaea L.) by Antagonistic Bacteria

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Preparation
2.2. Bacterial Strains
2.3. In Vitro Confrontation Bioassay
2.4. Effect of the Bacteria on the Cytology of Pythium
2.5. Volatile Organic Compounds (VOCs) Bioassay
2.6. Bacterial Cell-Free Filtrate Effects on Mycelial Growth
2.7. In Vivo Bioassay
2.8. Statistical Analysis
3. Results
3.1. Antagonism Effect
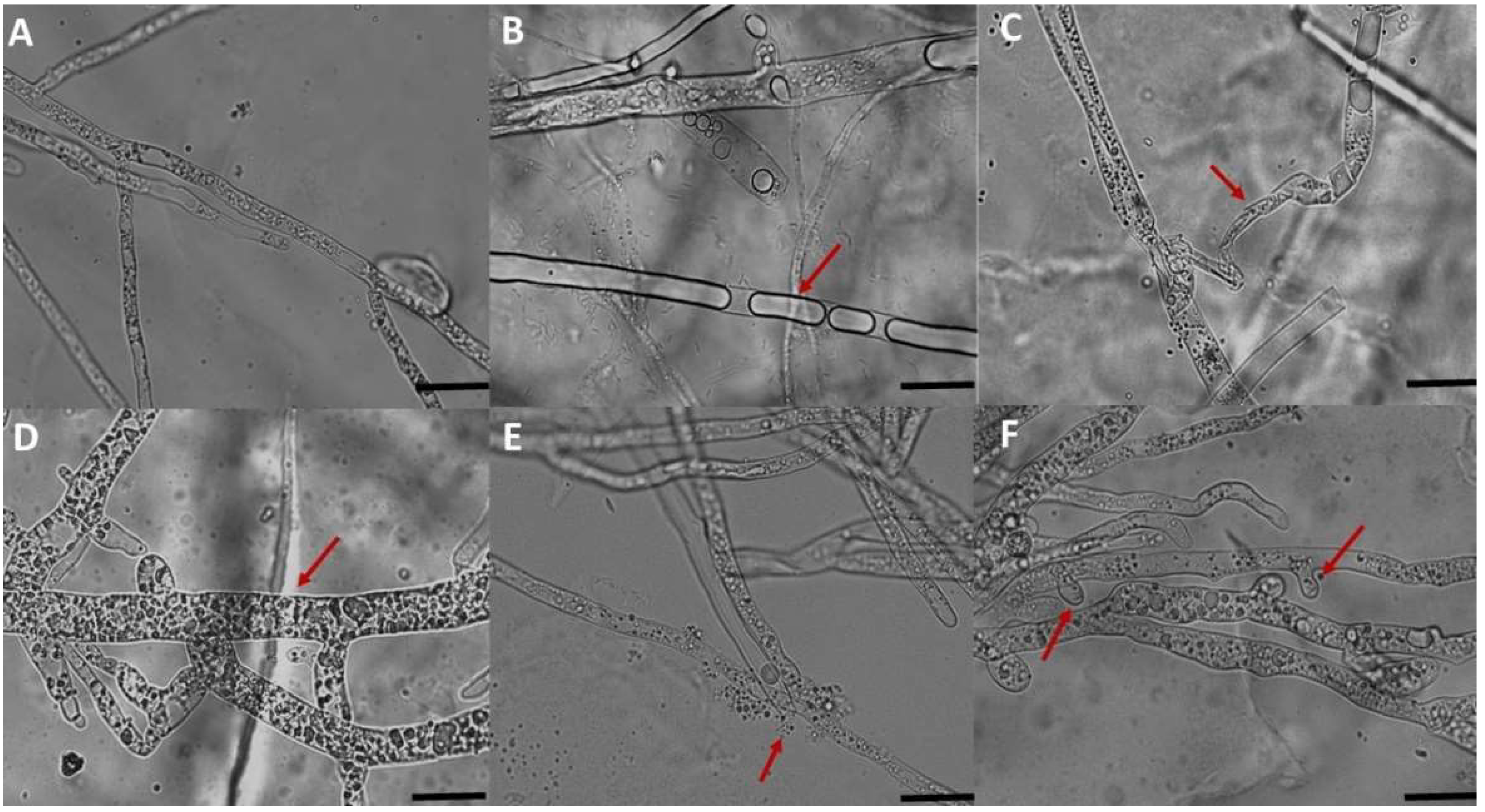
3.2. Microscopic Observation
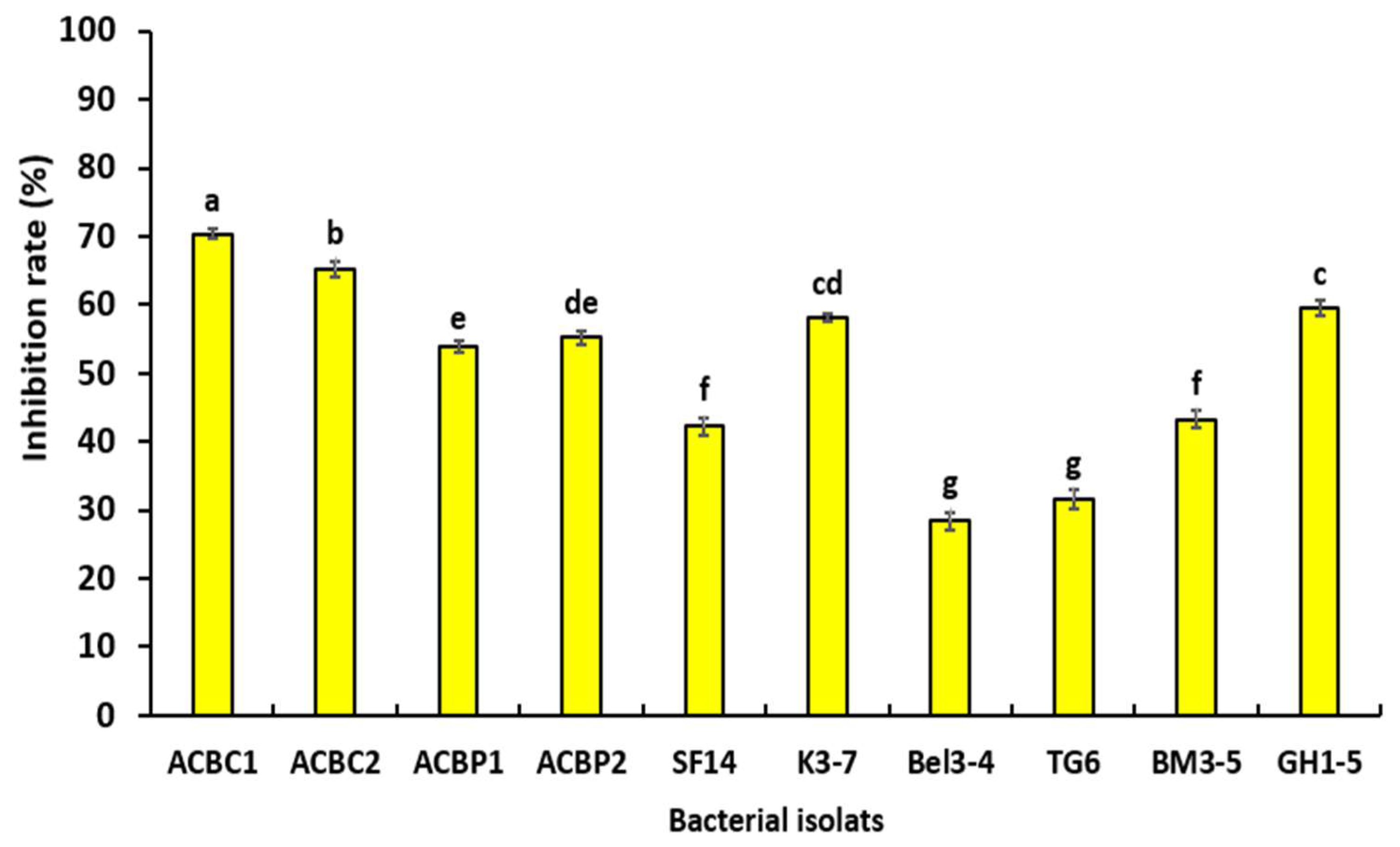
3.3. Effect of Bacterial Volatile Organic Compounds on Mycelial Growth
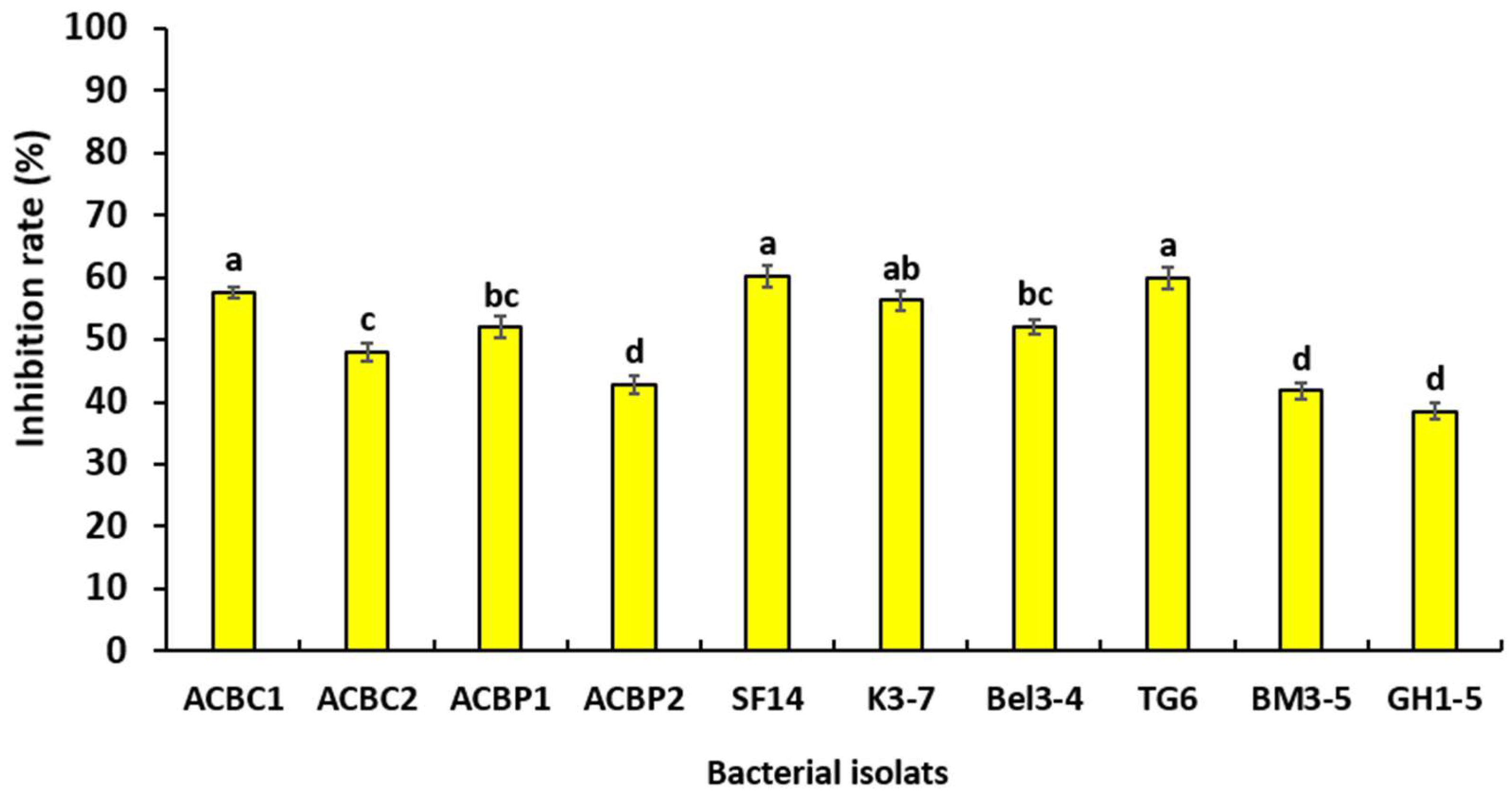
3.4. Effect of Bacterial Filtrates on Mycelial Growth
3.5. In Planta Bioassay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraga, H.; Moriondo, M.; Leolini, L. Mediterranean Olive Orchards under Climate Change: A Review of Future Impacts and Adaptation Strategies. Agronomy 2021, 11, 56. [Google Scholar] [CrossRef]
- Chliyeh, M.; Selmaoui, K.; Touhami, A.O.; Abdelkarim, F. Survey of the Fungal Species Associated to Olive-Tree (Olea europaea L.). Int. J. Recent Biotechnol. 2014, 2, 15–32. [Google Scholar]
- Ben Amira, M.; Lopez, D.; Triki, A.; Khouaja, A.; Chaar, H.; Fumanal, B.; Gousset-dupont, A.; Bonhomme, L.; Label, P.; Goupil, P.; et al. Beneficial Effect of Trichoderma harzianum Strain Ths97 in Biocontrolling Fusarium solani Causal Agent of Root Rot Disease in Olive Trees. Biol. Control 2017, 110, 70–78. [Google Scholar] [CrossRef]
- Mousa, M.S.; Ali, M.K.; Mosa, A.A.; Elewa, I. Root Rot Disease of Olive Transplants and Its Biological Control. Arab Univ. J. Agric. Sci. 2006, 14, 395–409. [Google Scholar] [CrossRef]
- Bouzoumita, A.; Metoui, M.; Jemni, M.; Kabaeir, N.; Belhouchette, K.; Ferchichi, A. The Efficacy of Various Bacterial Organisms for Biocontrol of Fusarium Root Rot of Olive in Tunisia. Pol. J. Environ. Stud. 2020, 29, 11–16. [Google Scholar] [CrossRef]
- Moustafa, S.M.N.; Elkanzi, N.A.A. Effect of the Newly Synthesized Pyrazole, And Pyrazolo Pyrimidine Derivatives on Pythium aphanidermatum (Edson) Fitzp. Egypt. J. Chem. 2021, 64, 2857–2872. [Google Scholar] [CrossRef]
- Gharbi, Y.; Bouazizi, E.; Cheffi, M.; Amar, F. Ben Archives of Phytopathology and Plant Protection Investigation of Soil-Borne Fungi, Causal Agents of Olive Trees Wilt and Dieback in Tunisia. Arch. Phytopathol. Plant Prot. 2020, 53, 828–843. [Google Scholar] [CrossRef]
- González, M.; Serrano, M.S.; Sánchez, M.E. First Report of Pythium spiculum Causing Root Rot on Wild-Olive in Spain. Plant Dis. 2016, 100, 1023. [Google Scholar] [CrossRef]
- Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants 2020, 9, 743. [Google Scholar] [CrossRef]
- González, M.; Pérez-Sierra, A.; Serrano, M.S.; Sánchez, M.E. Two Phytophthora Species Causing Decline of Wild Olive (Olea europaea subsp. europaea Var. sylvestris). Plant Pathol. 2017, 66, 941–948. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Schena, L.; Agosteo, G.E.; Magnano di San Lio, G.; Cacciola, S.O. Phytophthora oleae Sp. Nov. Causing Fruit Rot of Olive in Southern Italy. Plant Pathol. 2018, 67, 1362–1373. [Google Scholar] [CrossRef]
- González, M.; Pérez-Sierra, A.; Sánchez, M.E. Phytophthora oleae, a New Root Pathogen of Wild Olives. Plant Pathol. 2019, 68, 901–907. [Google Scholar] [CrossRef]
- Wu, W.; Ogawa, F.; Ochiai, M.; Yamada, K.; Fukui, H. Common Strategies to Control Pythium Disease. Rev. Agric. Sci. 2020, 8, 58–69. [Google Scholar] [CrossRef]
- Balla, A.; Silini, A.; Cherif-silini, H.; Bouket, A.C.; Moser, W.K.; Nowakowska, J.A.; Oszako, T.; Benia, F.; Belbahri, L. The Threat of Pests and Pathogens and the Potential for Biological Control in Forest Ecosystems. Forests 2021, 12, 1579. [Google Scholar] [CrossRef]
- Bodah, E.T. Root Rot Diseases in Plants: A Review of Common Causal Agents and Management Strategies. Agric. Res. Technol. Open Access J. 2017, 5, 555661. [Google Scholar]
- Quyet, N.T.; Cuong, H.V.; Hong, L.T.; Soytong, K. Control Mechanism of Chaetomium Spp. and Its Biological Control of Citrus Root Rot in Pot and Field Experiments in Vietnam. J. Agric. Technol. 2016, 12, 329–336. [Google Scholar]
- Chliyeh, M.; Selmaoui, K.; Abdelkarim, F.; El Modafar, C. Geographical Distribution of Phytophtora Palmivora in Different Olive Growing Regions in Morocco. Int. J. Plant Anim. Environ. Sci. 2014, 4, 297–303. [Google Scholar]
- Shlevin, E.; Gamliel, A.; Katan, J.; Shtienberg, D. Multi-Study Analysis of the Added Bene Fi Ts of Combining Soil Solarization with Fumigants or Non-Chemical Measures. Crop Prot. 2018, 111, 58–65. [Google Scholar] [CrossRef]
- Porter, L.D.; Hamm, P.B.; David, N.L.; Gieck, S.L.; Miller, J.S.; Gundersen, B.; Inglis, D.A. Metalaxyl-M-Resistant Pythium Species in Potato Production Areas of the Pacific Northwest of the USA. Am. J. Potato Res. 2009, 86, 315–326. [Google Scholar] [CrossRef]
- Hassanisaadi, M.; Hosein, G.; Bonjar, S.; Hosseinipour, A.; Abdolshahi, R. Biological Control of Pythium aphanidermatum, the Causal Agent of Tomato Root Rot by Two Streptomyces Root Symbionts. Agronomy 2021, 11, 846. [Google Scholar] [CrossRef]
- Lookabaugh, E.C.; Kerns, J.P.; Cubeta, M.A.; Shew, B.B.; Pathology, P.; State, N.C. Fitness Attributes of Pythium aphanidermatum with Dual Resistance to Mefenoxam and Fenamidone. Plant Dis. 2018, 102, 1938–1943. [Google Scholar] [CrossRef]
- Halo, B.A.; Al-yahyai, R.A.; Al-sadi, A.M. Biological Control of Pythium aphanidermatum-Induced Cucumber and Radish Damping-off by an Endophytic Fungus, Cladosporium omanense Isolate 31R Cucumber and Radish Damping-o Ff by an Endophytic Fungus. Biocontrol Sci. Technol. 2020, 31, 235–251. [Google Scholar] [CrossRef]
- Cook, R.J. Making Greater Use of Introduced Microorganisms for Biological Control of Plant Pathogens. Annu. Rev. Phytopathol. 1993, 31, 53–80. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Martínez, P.G.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, D.; Singh, R. Biological Control of Postharvest Diseases of Fruits and Vegetables by Microbial Antagonists: A Review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty Years of Postharvest Biocontrol Research: Is It Time for a New Paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- O’Brien, P.A. Biological Control of Plant Diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef]
- Farhaoui, A.; Adadi, A.; Tahiri, A.; El Alami, N.; Khayi, S.; Mentag, R.; Ezrari, S.; Radouane, N.; Mokrini, F.; Belabess, Z.; et al. Biocontrol Potential of Plant Growth-Promoting Rhizobacteria (PGPR) against Sclerotiorum Rolfsii Diseases on Sugar Beet (Beta vulgaris L.). Physiol. Mol. Plant Pathol. 2022, 119, 101829. [Google Scholar] [CrossRef]
- Jimtha, J.C.; Mallikarjunaswamy, G.; Najiya, N. Probiotic Rhizospheric Bacillus Sp. from Zingiber officinale Rosc. Displays Antifungal Activity against Soft Rot Pathogen Pythium Sp. Curr. Plant Biol. 2021, 27, 100217. [Google Scholar] [CrossRef]
- Ben Khedher, S.; Boukedi, H.; Laarif, A.; Tounsi, S. Biosurfactant Produced by Bacillus Subtilis V26: A Potential Biological Control Approach for Sustainable Agriculture Development. Org. Agric. 2020, 10, 117–124. [Google Scholar] [CrossRef]
- Kalantari, S.; Marefat, A.; Naseri, B. Improvement of Bean Yield and Fusarium Root Rot Biocontrol Using Mixtures of Bacillus, Pseudomonas and Rhizobium. Trop. Plant Pathol. 2018, 43, 499–505. [Google Scholar] [CrossRef]
- Zhang, J.; Mavrodi, D.V.; Yang, M.; Thomashow, L.S.; Mavrodi, O.V.; Kelton, J.; Weller, D.M. Pseudomonas Synxantha 2-79 Transformed with Pyrrolnitrin Biosynthesis Genes Has Improved Biocontrol Activity against Soilborne Pathogens of Wheat and Canola. Phytopathology 2020, 110, 1010–1017. [Google Scholar] [CrossRef]
- Vurukonda, S.S.k.P.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol Activity of Streptomyces Spp. as Endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef]
- Newitt, J.T.; Prudence, S.M.M.; Hutchings, M.I.; Worsley, S.F. Biocontrol of Cereal Crop Diseases Using Streptomycetes. Pathogens 2019, 8, 78. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Tchameni, S.N.; Cotârle, M.; Ghinea, I.O.; Ampere, M.; Bedine, B. Involvement of Lytic Enzymes and Secondary Metabolites Produced by Trichoderma Spp. in the Biological Control of Pythium myriotylum. Int. Microbiol. 2019, 23, 179–188. [Google Scholar] [CrossRef]
- Lahlali, R.; Aksissou, W.; Lyousfi, N.; Ezrari, S.; Blenzar, A.; Tahiri, A.; Ennahli, S.; Hrustić, J.; MacLean, D.; Amiri, S. Biocontrol Activity and Putative Mechanism of Bacillus amyloliquefaciens (SF14 and SP10), Alcaligenes faecalis ACBC1, and Pantoea Agglomerans ACBP1 against Brown Rot Disease of Fruit. Microb. Pathog. 2020, 139, 103914. [Google Scholar] [CrossRef]
- Ezrari, S.; Mhidra, O.; Radouane, N.; Tahiri, A.; Polizzi, G.; Lazraq, A.; Lahlali, R. Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot. Plants 2021, 10, 872. [Google Scholar] [CrossRef]
- Bahadou, S.A.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New Potential Bacterial Antagonists for the Biocontrol of Fire Blight Disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef]
- Legrifi, I.; Al Figuigui, J.; Radouane, N.; Ezrari, S.; Belabess, Z.; Tahiri, A.; Amiri, S.; Lahlali, R. First Report of Pythium schmitthenneri on Olive Trees and in Morocco. Australas. Plant Dis. Notes 2022, 17, 3. [Google Scholar] [CrossRef]
- Zhu, C.; Lei, M.; Andargie, M.; Zeng, J.; Li, J. Antifungal Activity and Mechanism of Action of Tannic Acid against Penicillium Digitatum. Physiol. Mol. Plant Pathol. 2019, 107, 46–50. [Google Scholar] [CrossRef]
- Wahyuni, D.K.; Hafida, S.N.; Ermayanti, T.M.; Eko, B.P.; Wardoyo, H.P.; Utami, E.S.W. Hairy Root Induction on Justicia gendarussa by Various Density of Agrobacterium Rhizogenes Strain LB 510. Biosaintifika J. Biol. Biol. Educ. 2017, 9, 26–32. [Google Scholar] [CrossRef]
- Lahlali, R.; Bajii, M.; Jijakli, M.H. Isolation and Evaluation of Bacteria and Fungi as Biological Control Agents. Commun. Agric. Appl. Biol. Sci. 2007, 72, 973–982. [Google Scholar]
- Trivedi, P.; Pandey, A.; Palni, L.M.S. In Vitro Evaluation of Antagonistic Properties of Pseudomonas corrugata. Microbiol. Res. 2008, 163, 329–336. [Google Scholar] [CrossRef]
- Guevara-avendan, E.; Carrillo, J.D.; Moreno, K.; Méndez-Bravo, A.; Guerrero-Analco, J.A.; Reverchon, F. Antifungal Activity of Avocado Rhizobacteria against Fusarium euwallaceae and Graphium Spp., Associated with Euwallacea Spp. Nr. Fornicatus, and Phytophthora cinnamomi. Antonie Van Leeuwenhoek 2018, 111, 563–572. [Google Scholar] [CrossRef]
- Li, Z.; Guo, B.; Wan, K.; Cong, M.; Huang, H.; Ge, Y. Effects of Bacteria-Free Filtrate from Bacillus Megaterium Strain L2 on the Mycelium Growth and Spore Germination of Alternaria Alternata. Biotechnol. Biotechnol. Equip. 2015, 29, 1062–1068. [Google Scholar] [CrossRef]
- Moradi, M.; Nejad, F.J.; Hosein, G.; Bonjar, S.; Fani, S.R. Efficacy of Bacillus Subtilis Native Strains for Biocontrol of Phytophthora Crown and Root Rot of Pistachio in Iran. Trop. Plant Pathol. 2018, 43, 306–313. [Google Scholar] [CrossRef]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First Report of Root Rot Caused by Phytophthora Bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef] [PubMed]
- Simamora, A.V.; Stukely, M.J.C.; Barber, P.A.; Hardy, G.E.S.; Burgess, T.I. Age-Related Susceptibility of Eucalyptus Species to Phytophthora Accepted Article Boodjera. Plant Pathol. 2016, 66, 501–512. [Google Scholar] [CrossRef]
- Jabiri, S.; Bahra, C.; Maclean, D.; Radouane, N.; Barka, E.A. Phytopythium vexans Associated with Apple and Pear Decline in the Saïss Plain of Morocco. Microorganisms 2021, 9, 1916. [Google Scholar] [CrossRef] [PubMed]
- Miao, G.; Han, J.; Wang, C.; Zhang, K.; Miao, G. Growth Inhibition and Induction of Systemic Resistance against Pythium aphanidermatum by Bacillus Simplex Strain HS-2 Growth Inhibition and Induction of Systemic Resistance against. Biocontrol Sci. Technol. 2018, 28, 1114–1127. [Google Scholar] [CrossRef]
- Peter, P.; Divakaran, K.; Kaniyala, H.; Nair, A.R. Endophytic Bacillus Spp. from Zingiber zerumbet Rhizome, Rhizosphere and Axenic Cultures: Characterisation and Identification of Isolates with Plant Growth Promoting Activities and Antagonism to Soft-Rot Causative Pythium myriotylum. Arch. Phytopathol. Plant Prot. 2021, 54, 2007–2022. [Google Scholar] [CrossRef]
- Al-daghari, D.S.S.; Al-abri, S.A.; Al-mahmooli, I.H. Efficacy of Native Antagonistic Rhizobacteria in the Biological Control of Pythium aphanidermatum -Induced Damping-off of Cucumber in Oman. J. Plant Pathol. 2019, 102, 305–310. [Google Scholar] [CrossRef]
- Fatouros, G.; Gkizi, D.; Fragkogeorgi, G.A.; Paplomatas, E.J.; Tjamos, S.E. Biological Control of Pythium, Rhizoctonia and Sclerotinia in Lettuce: The Plant Protective Activity of the Bacterium Paenibacillus Alvei K165 Is Associated with the Induction of Systemic Resistance. Plant Pathol. 2017, 67, 418–425. [Google Scholar] [CrossRef]
- Al-hussini, H.S.; Al-rawahi, A.Y.; Al-marhoon, A.A.; Al-abri, S.A.; Al-mahmooli, I.H. Biological Control of Damping-off of Tomato Caused by Pythium aphanidermatum by Using Native Antagonistic Rhizobacteria Isolated from Omani Soil. J. Plant Pathol. 2018, 101, 315–322. [Google Scholar] [CrossRef]
- Bardin, S.D.; Huang, H.C.; Liu, L.; Yanke, L.J. Control, by Microbial Seed Treatment, of Damping- off Caused by Pythium Sp. on Canola, Safflower, Dry Pea, and Sugar Beet 1. Can. J. Plant Pathol. 2003, 25, 268–275. [Google Scholar] [CrossRef]
- Vasebi, Y.; Alizadeh, A.; Safaie, N. Pantoea Agglomerans ENA1 as a Biocontrol Agent of Macrophomia Phaseolina and Growth Enhancer of Soybean. J. Crop Prot. 2015, 4, 43–57. [Google Scholar]
- Dunne, C.; Crowley, J.J.; Mo, Y.; Dowling, D.N.; Bruijn, S.; Gara, F.O. Biological Control of Pythium Ultimum by Stenotrophomonas maltophilia W81 Is Mediated by an Extracellular Proteolytic Activity. Microbiology 1997, 143, 3921–3931. [Google Scholar] [CrossRef]
- Alijani, Z.; Amini, J.; Ashengroph, M.; Bahramnejad, B. Volatile Compounds Mediated Effects of Stenotrophomonas maltophilia Strain UN1512 in Plant Growth Promotion and Its Potential for the Biocontrol of Colletotrichum nymphaeae. Physiol. Mol. Plant Pathol. 2020, 112, 101555. [Google Scholar] [CrossRef]
- Amna, X.Y.; Farooq, M.A.; Javed, M.T.; Kamran, M.A.; Mukhtar, T.; Ali, J.; Tabassum, T.; ur Rehman, S.; Munis, M.F.H.; Sultan, T.; et al. Plant Physiology and Biochemistry Multi-Stress Tolerant PGPR Bacillus Xiamenensis PM14 Activating Sugarcane (Saccharum officinarum L.) Red Rot Disease Resistance. Plant Physiol. Biochem. 2020, 151, 640–649. [Google Scholar] [CrossRef]
- Jayakumar, A.; Nair, I.C.; Radhakrishnan, E.K. Environmental Adaptations of an Extremely Plant Beneficial Bacillus Subtilis Dcl1 Identified Through the Genomic and Metabolomic Analysis. Microb. Ecol. 2020, 81, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Wise, C.; Novitsky, L.; Tsopmo, A.; Avis, T.J. Production and Antimicrobial Activity of 3-Hydroxypropionaldehyde from Bacillus Subtilis Strain CU12. J. Chem. Ecol. 2012, 38, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Sahu, K.P.; Kumar, A.; Patel, A.; Kumar, M.; Gopalakrishnan, S.; Prakash, G.; Gogoi, R. Rice Blast Lesions: An Unexplored Phyllosphere Microhabitat for Novel Antagonistic Bacterial Species Against Magnaporthe oryzae. Ecology 2020, 81, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Cheffi, M.; Bouket, A.C.; Alenezi, F.N.; Luptakova, L.; Belka, M.; Vallat, A.; Rateb, M.E.; Tounsi, S.; Triki, M.A. Olea europaea L. Root Endophyte Bacillus velezensis OEE1 Counteracts Oomycete and Fungal Harmful Pathogens and Harbours a Large Repertoire of Secreted and Volatile Metabolites and Beneficial Functional Genes. Microorganisms 2019, 7, 314. [Google Scholar] [CrossRef]
- Jimtha, J.C.; Jishma, P.; Arathy, G.B.; Anisha, C.; Radhakrishnan, E.K. Identification of Plant Growth Promoting Rhizosphere Bacillus Sp. WG4 Antagonistic to Pythium myriotylum and Its Enhanced Antifungal Effect in Association with Trichoderma. J. Soil Sci. Plant Nutr. 2016, 16, 578–590. [Google Scholar] [CrossRef]
- Gong, A.; Wu, N.; Kong, X.; Zhang, Y.; Hu, M. Inhibitory Effect of Volatiles Emitted from Alcaligenes faecalis N1-4 on Aspergillus flavus and Aflatoxins in Storage. Front. Microbiol. 2019, 10, 1419. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Wang, W.; Wang, M.; Sun, B. The Symbiotic Bacteria Alcaligenes faecalis of the Entomopathogenic Nematodes Oscheius Spp. Exhibit Potential Biocontrol of Plant- and Entomopathogenic Fungi. Microb. Biotechnol. 2019, 12, 459–471. [Google Scholar] [CrossRef]
- Kilany, M.; Ibrahim, E.H.; Al Amry, S.; Roman, S. Microbial Suppressiveness of Pythium Damping-Off Diseases. In Organic Amendments and Soil Suppressiveness in Plant Disease Management; Springer: Cham, Switzerland, 2015; pp. 187–206. [Google Scholar] [CrossRef]
- Singh, R.; Sachan, N.S. Review on Biological Control of Soil Borne Fungi in Vegetable Crops. HortFlora Res. Spectr. 2013, 2, 72–76. [Google Scholar]
- Abd-el-kareem, F.; Elshahawy, I.E.; Abd-elgawad, M.M.M. Application of Bacillus Pumilus Isolates for Management of Black Rot Disease in Strawberry. Egypt. J. Biol. Pest Control 2021, 31, 25. [Google Scholar] [CrossRef]
- Montes-osuna, N.; Cernava, T.; Carmen, G.; Berg, G. Identification of Volatile Organic Compounds Emitted by Two Beneficial Endophytic Pseudomonas Strains from Olive Roots. Plants 2022, 11, 318. [Google Scholar] [CrossRef]
- Pascal, M.; Dietel, K.; Grosch, R.; Cernava, T.; Berg, G. Microbiota Associated with Sclerotia of Soilborne Fungal Pathogens—A Novel Source of Biocontrol Agents Producing Bioactive Volatiles. Phytobiomes J. 2019, 3, 125–136. [Google Scholar] [CrossRef]
- Tilocca, B.; Cao, A.; Migheli, Q. Scent of a Killer: Microbial Volatilome and Its Role in the Biological Control of Plant Pathogens. Front. Microbiol. 2020, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xiao, J.; Zhang, Y.; Li, R.; Liu, L.; Deng, J. Postharvest Biology and Technology Biocontrol Ability and Action Mechanism of Bacillus halotolerans against Botrytis Cinerea Causing Grey Mould in Postharvest Strawberry Fruit. Postharvest Biol. Technol. 2021, 174, 111456. [Google Scholar] [CrossRef]
- Slama, H.B.; Cherif-silini, H.; Bouket, A.C.; Qader, M. Screening for Fusarium Antagonistic Bacteria from Contrasting Niches Designated the Endophyte Bacillus halotolerans as Plant Warden against Fusarium. Front. Microbiol. 2019, 9, 3236. [Google Scholar] [CrossRef]
- Duan, Y.; Chen, R.; Zhang, R.; Jiang, W.; Chen, X.; Yin, C. Isolation, Identification, and Antibacterial Mechanisms of Bacillus amyloliquefaciens QSB-6 and Its Effect on Plant Roots. Front. Microbiol. 2021, 12, 746799. [Google Scholar] [CrossRef]
- Sánchez-fernández, R.E.; Diaz, D.; Duarte, G.; Lappe-oliveras, P.; Sánchez, S.; Macías-rubalcava, M.L.; Macías-rubalcava, M.L. Antifungal Volatile Organic Compounds from the Endophyte Nodulisporium Sp. Strain GS4d2II1a: A Qualitative Change in the Intraspecific and Interspecific Interactions with Pythium aphanidermatum Interaction. Ecology 2015, 71, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal Volatile Organic Compounds: A Review with Emphasis on Their Biotechnological Potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Maung, E.C.H.; Choi, T.G.; Nam, H.H.; Kim, K.Y. Role of Bacillus amyloliquefaciens Y1 in the Control of Fusarium Wilt Disease and Growth Promotion of Tomato Role of Bacillus amyloliquefaciens Y1 in the Control of Fusarium Wilt Disease and Growth Promotion of Tomato. Biocontrol Sci. Technol. 2017, 27, 1400–1415. [Google Scholar] [CrossRef]
- Khabbaz, S.E.; Zhang, L.; Cáceres, L.A.; Sumarah, M.; Wang, A.; Abbasi, P.A. Characterisation of Antagonistic Bacillus and Pseudomonas Strains for Biocontrol Potential and Suppression of Damping-off and Root Rot Diseases. Ann. Appl. Biol. 2015, 166, 456–471. [Google Scholar] [CrossRef]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.M.; Ramaraj, G.B.; Ponnusamy, P.; Sekar, J.; Ramalingam, P.V. Biocontrol Efficiency of Native Plant Growth Promoting Rhizobacteria against Rhizome Rot Disease of Turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Salem, A.A.; Abdel-Rahman, H.M. Cellulolytic Activity of Trichoderma Reesei and Bacillus Subtilis against the Plant Pathogen Pythium Debaryanum. Environ. Biodivers. Soil Secur. 2021, 5, 105–119. [Google Scholar] [CrossRef]
- Aydi Ben Abdallah, R.; Mokni-Tlili, S.; Nefzi, A.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Biocontrol of Fusarium Wilt and Growth Promotion of Tomato Plants Using Endophytic Bacteria Isolated from Nicotiana Glauca Organs. Biol. Control 2016, 97, 80–88. [Google Scholar] [CrossRef]
- Sen, K.S.; Dora, K.T.; Bandyopadhyay, B.; Das Mohapatra, K.P.; Raut, S. Thermostable Alpha-Amylase Enzyme Production from Hot Spring Isolates Alcaligenes faecalis SSB17—Statistical Optimization. Biocatal. Agric. Biotechnol. 2014, 3, 218–226. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Hydrogen Cyanide Production by Soil Bacteria: Biological Control of Pests and Promotion of Plant Growth in Sustainable Agriculture. Pedosphere Int. J. 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Sendi, Y.; Pfeiffer, T.; Koch, E.; Mhadhbi, H.; Mrabet, M. Potential of Common Bean (Phaseolus vulgaris L.) Root Microbiome in the Biocontrol of Root Rot Disease and Traits of Performance. J. Plant Dis. Prot. 2020, 127, 453–462. [Google Scholar] [CrossRef]
- Jha, B.K.; Gandhi Pragash, M.; Raman, G.; Sakthivel, N. Simultaneous Phosphate Solubilization Potential and Antifungal Activity of New Fluorescent Pseudomonad Strains, Pseudomonas aeruginosa, P. plecoglossicida and P. mosselii. World J. Microbiol. Biotechnol. 2008, 25, 573–581. [Google Scholar] [CrossRef]
- Habiba, R.N.; Ali, S.A.; Sultana, V.; Ara, J.; Ehteshamul-Haque, S. Evaluation of Biocontrol Potential of Epiphytic Fluorescent Pseudomonas Associated with Healthy Fruits and Vegetables against Root Rot and Root Knot Pathogens of Mungbean. Pak. J. Bot. 2016, 48, 1299–1303. [Google Scholar]
- Rehman, F.U.; Kalsoom, M.; Adnan, M.; Toor, M.D. Plant Growth Promoting Rhizobacteria and Their Mechanisms Involved in Agricultural Crop Production: A Review SunText Review of Biotechnology Plant Growth Promoting Rhizobacteria and Their Mechanisms Involved in Agricultural Crop Production: A Review. SunText Rev. BioTechnol. 2020, 35, 1044–1051. [Google Scholar] [CrossRef]
- Defilippi, S.; Groulx, E.; Megalla, M.; Mohamed, R.; Avis, T.J. Fungal Competitors Affect Production of Antimicrobial Lipopeptides in Bacillus Subtilis Strain B9-5. J. Chem. Ecol. 2018, 44, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Farace, G.; Fernandez, O.; Jacquens, L.; Coutte, F.; Krier, F.; Jacques, P.; Clément, C.; Barka, E.A.I.T.; Jacquard, C.; Dorey, S. Cyclic Lipopeptides from Bacillus Subtilis Activate Distinct Patterns of Defence Responses in Grapevine. Mol. Plant Pathol. 2014, 16, 177–187. [Google Scholar] [CrossRef]
- de Souza Freitas, F.; Coelho de Assis Lage, T.; Ayupe, B.A.L.; de Paula Siqueira, T.; de Barros, M.; Tótola, M.R. Bacillus Subtilis TR47II as a Source of Bioactive Lipopeptides against Gram-Negative Pathogens Causing Nosocomial Infections. 3 Biotech 2020, 10, 474. Available online: https://Link.Springer.Com/Article/10.1007/S13205-020-02459-Z (accessed on 12 May 2022). [CrossRef] [PubMed]
- Raaijmakers, J.M.; de Bruijn, I.; Nybroe, O.; Ongena, M. Natural Functions of Lipopeptides from Bacillus and Pseudomonas: More than Surfactants and Antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef]
- Ali, N.; Pang, Z.; Wang, F.; Xu, B.; El-Seedi, H.R. Lipopeptide Biosurfactants from Bacillus Spp.: Types, Production, Biological Activities, and Applications in Food. J. Food Qual. 2022, 2022, 3930112. [Google Scholar] [CrossRef]
- Cochrane, S.A.; Vederas, J.C. Lipopeptides from Bacillus and Paenibacillus Spp.: A Gold Mine of Antibiotic Candidates. Med. Res. Rev. 2016, 36, 4–31. [Google Scholar] [CrossRef] [PubMed]
- Stincone, P.; Veras, F.F.; Pereira, J.Q.; Mayer, F.Q.; Varela, A.P.M.; Brandelli, A. Diversity of Cyclic Antimicrobial Lipopeptides from Bacillus P34 Revealed by Functional Annotation and Comparative Genome Analysis. Microbiol. Res. 2020, 238, 126515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Khabbaz, S.E.; Wang, A.; Li, H.; Abbasi, P.A. Detection and Characterization of Broad-Spectrum Anti-Pathogen Activity of Novel Rhizobacterial Isolates and Suppression of Fusarium Crown and Root Rot Disease of Tomato. J. Appl. Microbiol. 2015, 118, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Park, D.; Kim, K.; Lim, S.M.; Yu, N.H.; Kim, S.; Kim, H.; Jung, K.S.; Jang, J.Y.; Park, J.; et al. Characterization of Bacillus amyloliquefaciens DA12 Showing Potent Antifungal Activity against Mycotoxigenic Fusarium Species. Plant Pathol. 2017, 33, 499–507. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Composition | Repetitions | Period |
|---|---|---|---|
| ACBC1 | SS + pathogen + ACBC1 | 6 | 2 months |
| ACBC2 | SS + pathogen + ACBC2 | ||
| ACBP1 | SS + pathogen + ACBP1 | ||
| ACBP2 | SS + pathogen + ACBP2 | ||
| SF14 | SS + pathogen + SF14 | ||
| K3-7 | SS + pathogen + K3-7 | ||
| Bel3-4 | SS + pathogen + Bel3-4 | ||
| TG6 | SS + pathogen + TG6 | ||
| BM3-5 | SS + pathogen + BM3-5 | ||
| GH1-5 | SS + pathogen + GH1-5 | ||
| C+ | SS + Pathogen | ||
| C− | SS alone |
| Bacterial Isolate Code | Species | Accession Numbers | Inhibition Rates (%) |
|---|---|---|---|
| ACBC1 | Alcaligenes faecalis | KY357285 | 85.14 ± 1.13 a |
| ACBC2 | Pantoea agglomerans | KY357286 | 80.58 ± 1.31 bc |
| ACBP1 | P. agglomerans | KY357287 | 77.79 ± 0.64 cde |
| ACBP2 | P. agglomerans | KY357288 | 78.38 ± 0.64 bcd |
| SF14 | Bacillus amyloliquefaciens | KY357298 | 81.76 ± 1.76 ab |
| K3-7 | B. xiamenensis | MW843010 | 77.89 ± 1.96 cd |
| Bel3-4 | Sphingobacterium multivorum | MW856827 | 73.97 ± 0.87 f |
| TG6 | B. subtilis | MW847628 | 74.45 ± 1.48 ef |
| BM3-5 | B. halotolerans | MW847951 | 76.49 ± 0.53 def |
| GH1-5 | Stenotrophomonas maltophilia | MW848819 | 60.47 ± 1.04 g |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legrifi, I.; Al Figuigui, J.; El Hamss, H.; Lazraq, A.; Belabess, Z.; Tahiri, A.; Amiri, S.; Barka, E.A.; Lahlali, R. Potential for Biological Control of Pythium schmitthenneri Root Rot Disease of Olive Trees (Olea europaea L.) by Antagonistic Bacteria. Microorganisms 2022, 10, 1635. https://doi.org/10.3390/microorganisms10081635
Legrifi I, Al Figuigui J, El Hamss H, Lazraq A, Belabess Z, Tahiri A, Amiri S, Barka EA, Lahlali R. Potential for Biological Control of Pythium schmitthenneri Root Rot Disease of Olive Trees (Olea europaea L.) by Antagonistic Bacteria. Microorganisms. 2022; 10(8):1635. https://doi.org/10.3390/microorganisms10081635
Chicago/Turabian StyleLegrifi, Ikram, Jamila Al Figuigui, Hajar El Hamss, Abderrahim Lazraq, Zineb Belabess, Abdessalem Tahiri, Said Amiri, Essaid Ait Barka, and Rachid Lahlali. 2022. "Potential for Biological Control of Pythium schmitthenneri Root Rot Disease of Olive Trees (Olea europaea L.) by Antagonistic Bacteria" Microorganisms 10, no. 8: 1635. https://doi.org/10.3390/microorganisms10081635