
Uncovering New Diversity of Photosynthetic Microorganisms from the Mediterranean Region



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Strains Isolation, and Growth
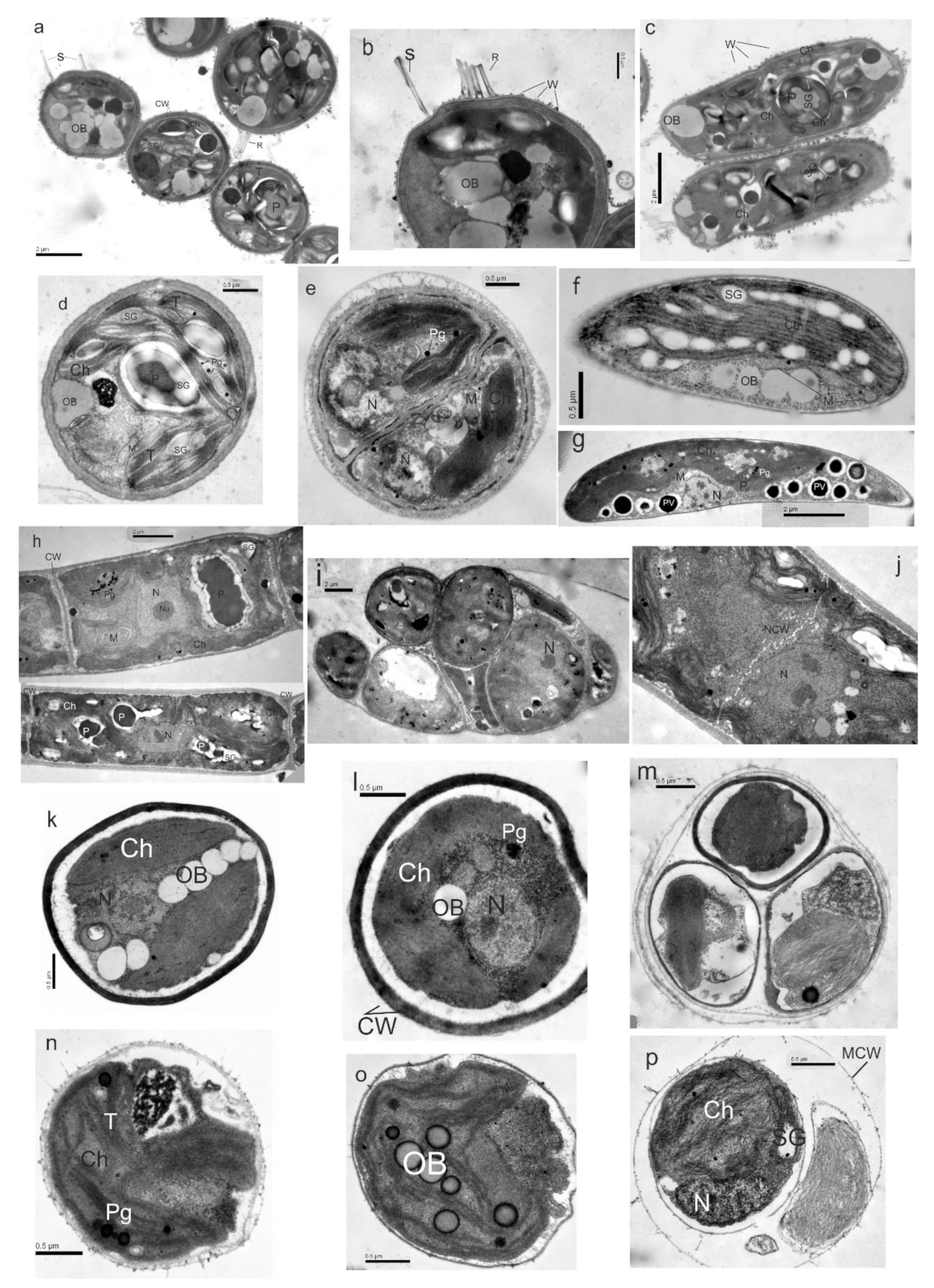
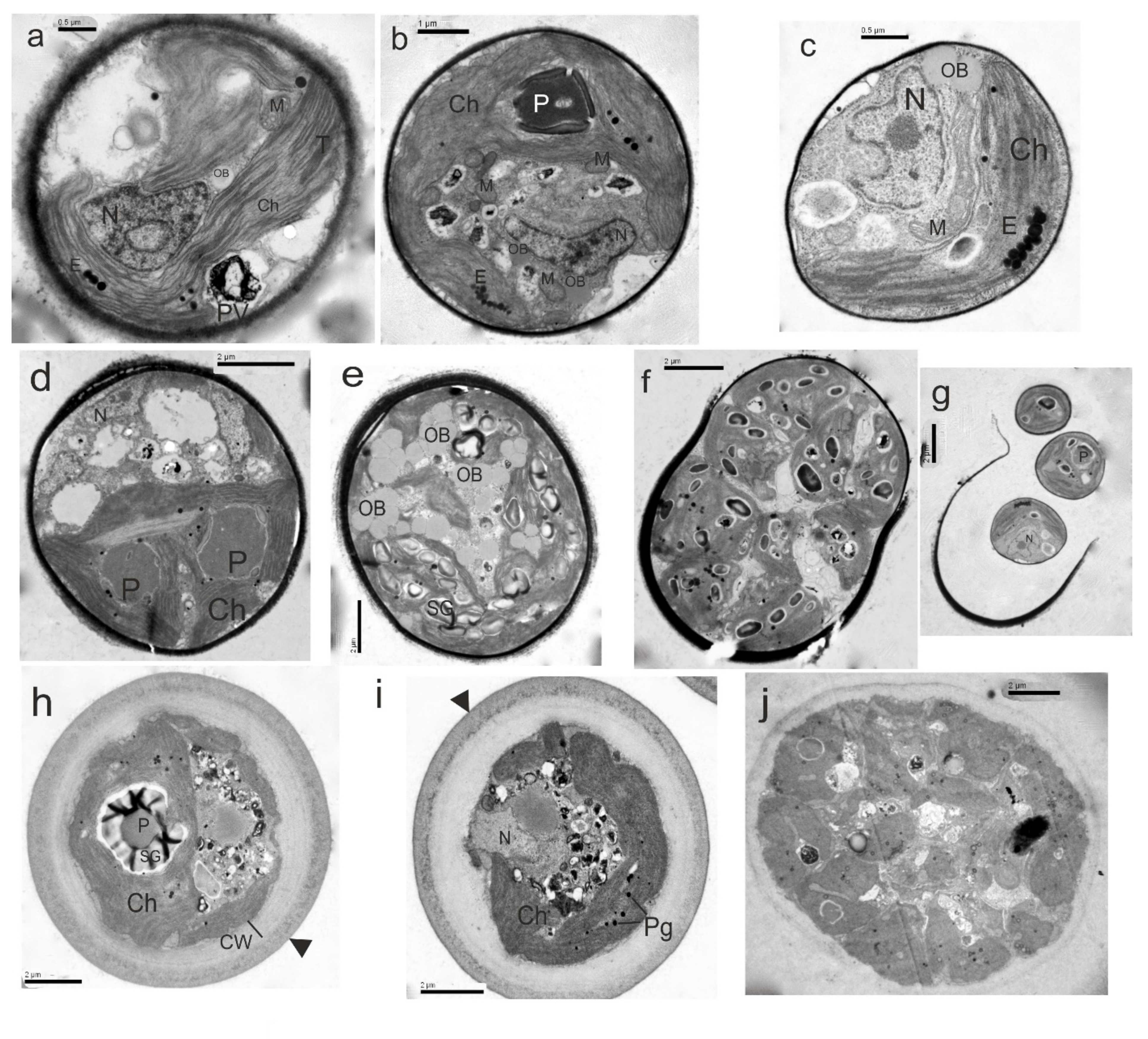
2.2. Morphology and Ultrastructure
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analysis
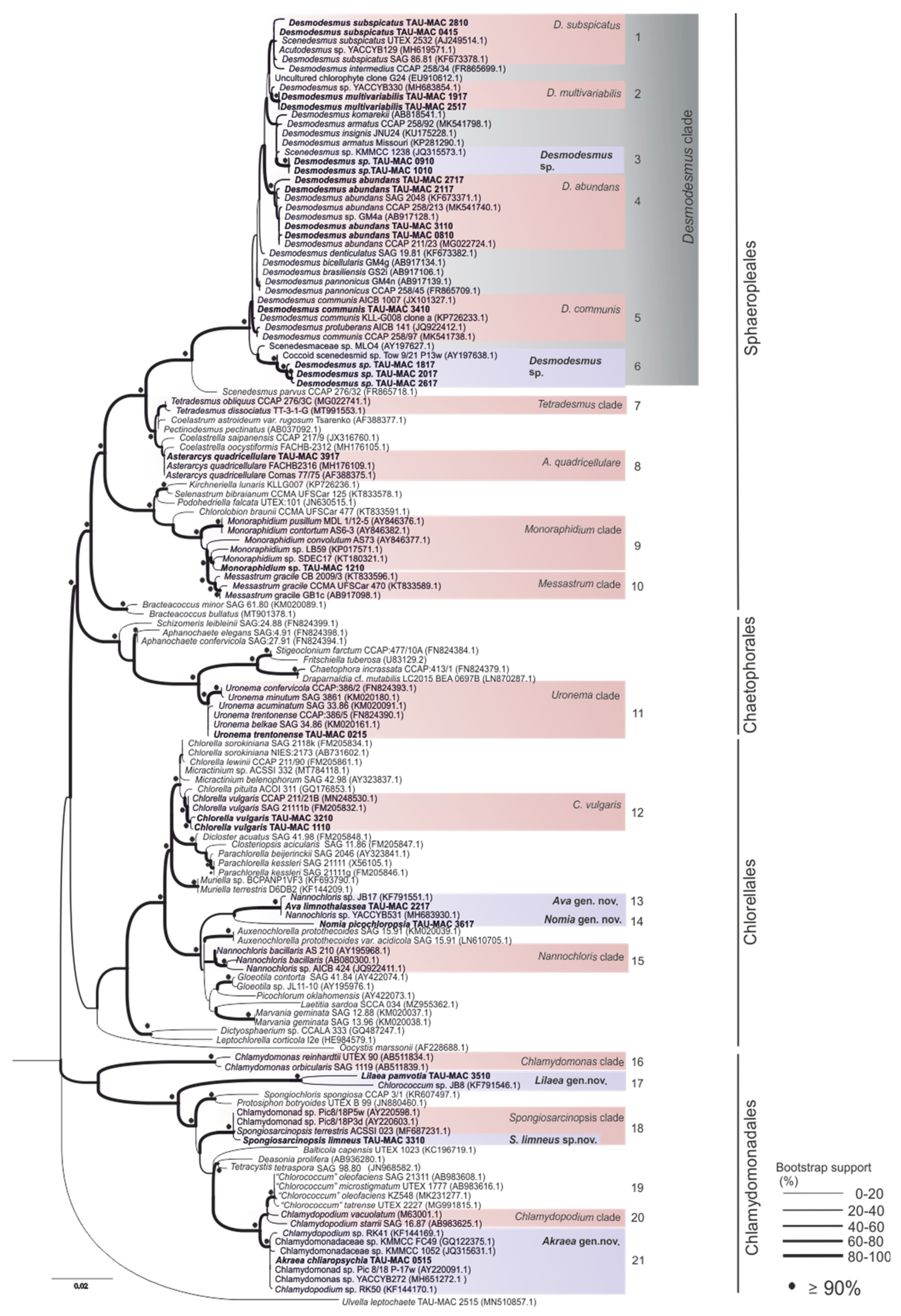
3. Results and Discussion
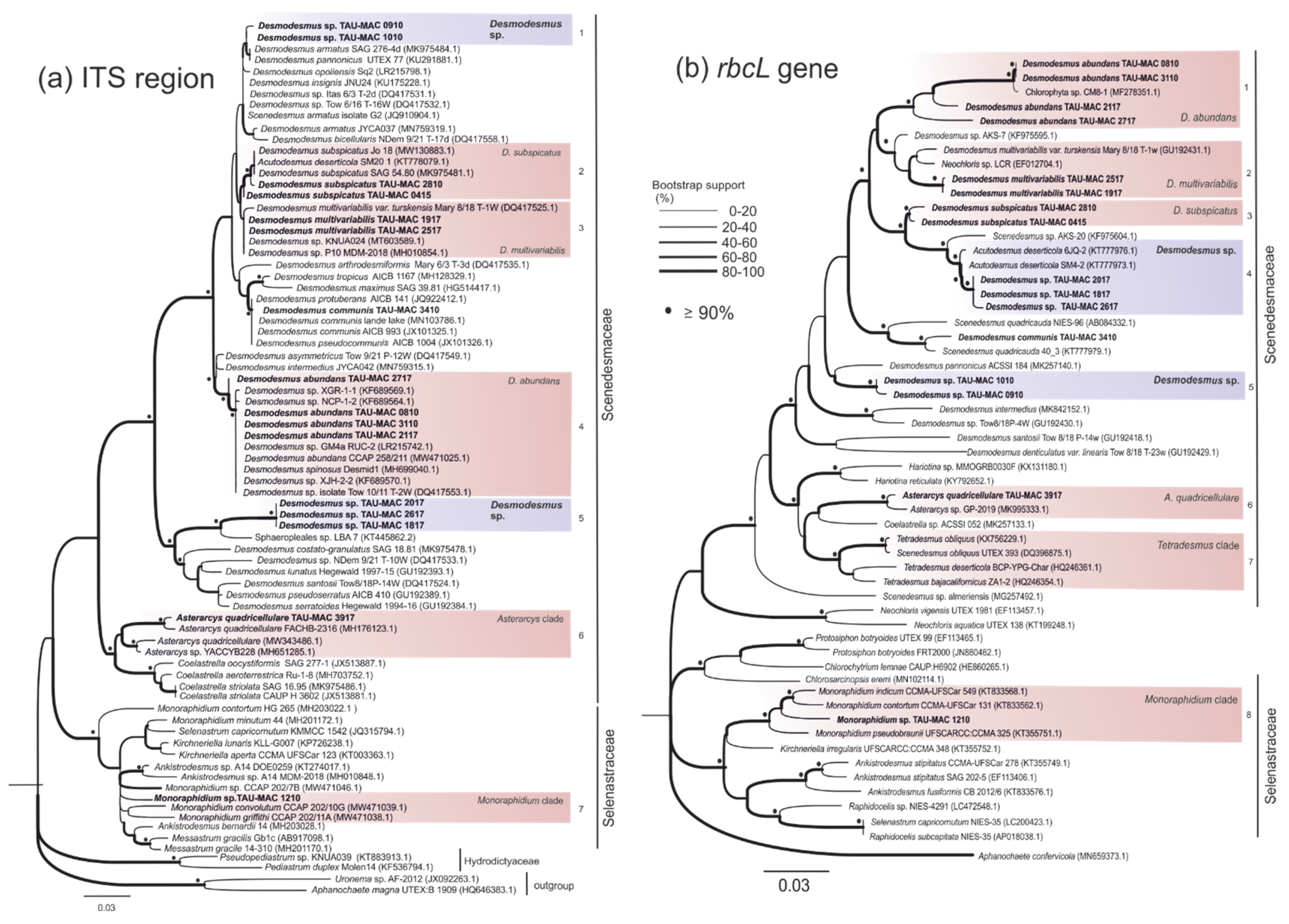
3.1. Sphaeropleales
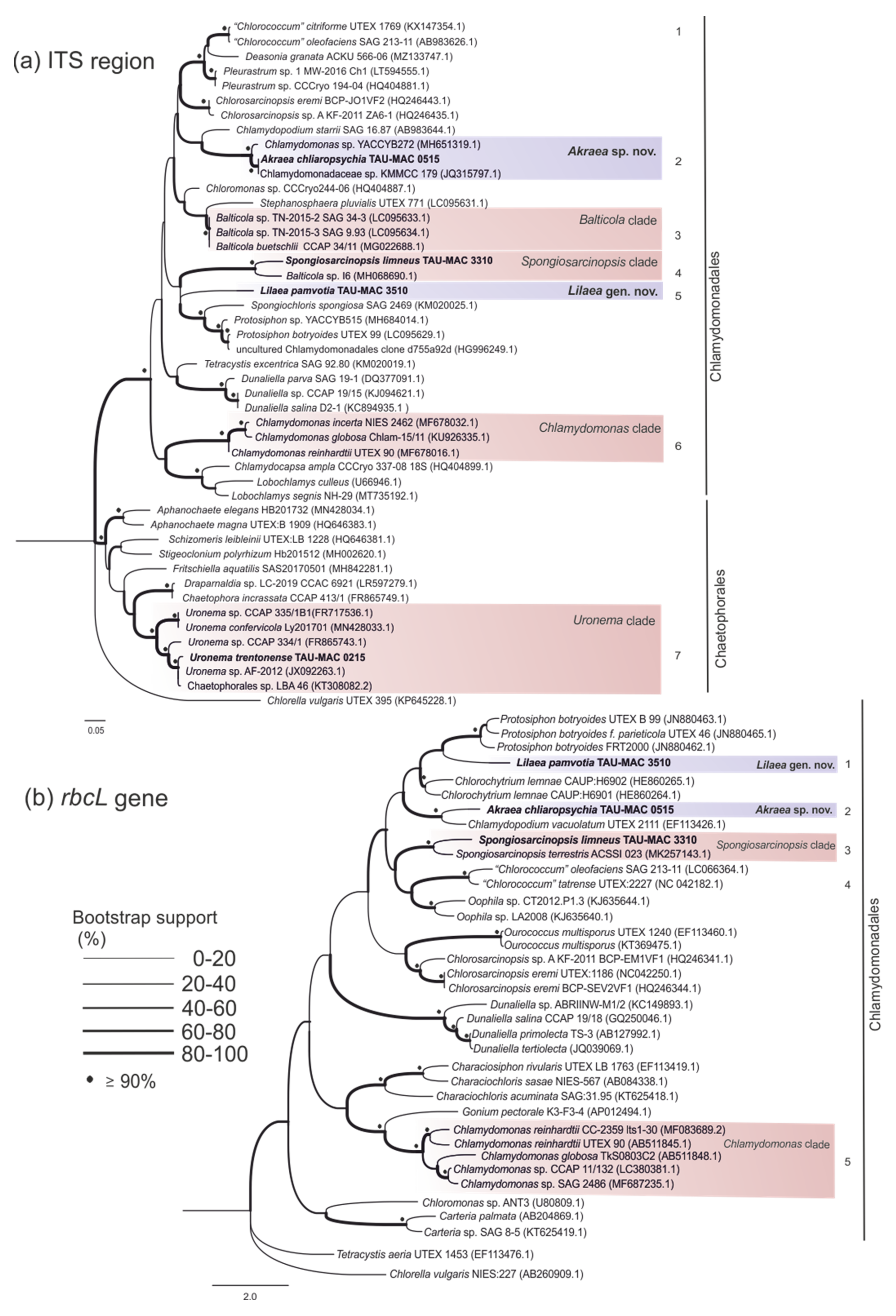
3.2. Chaetophorales
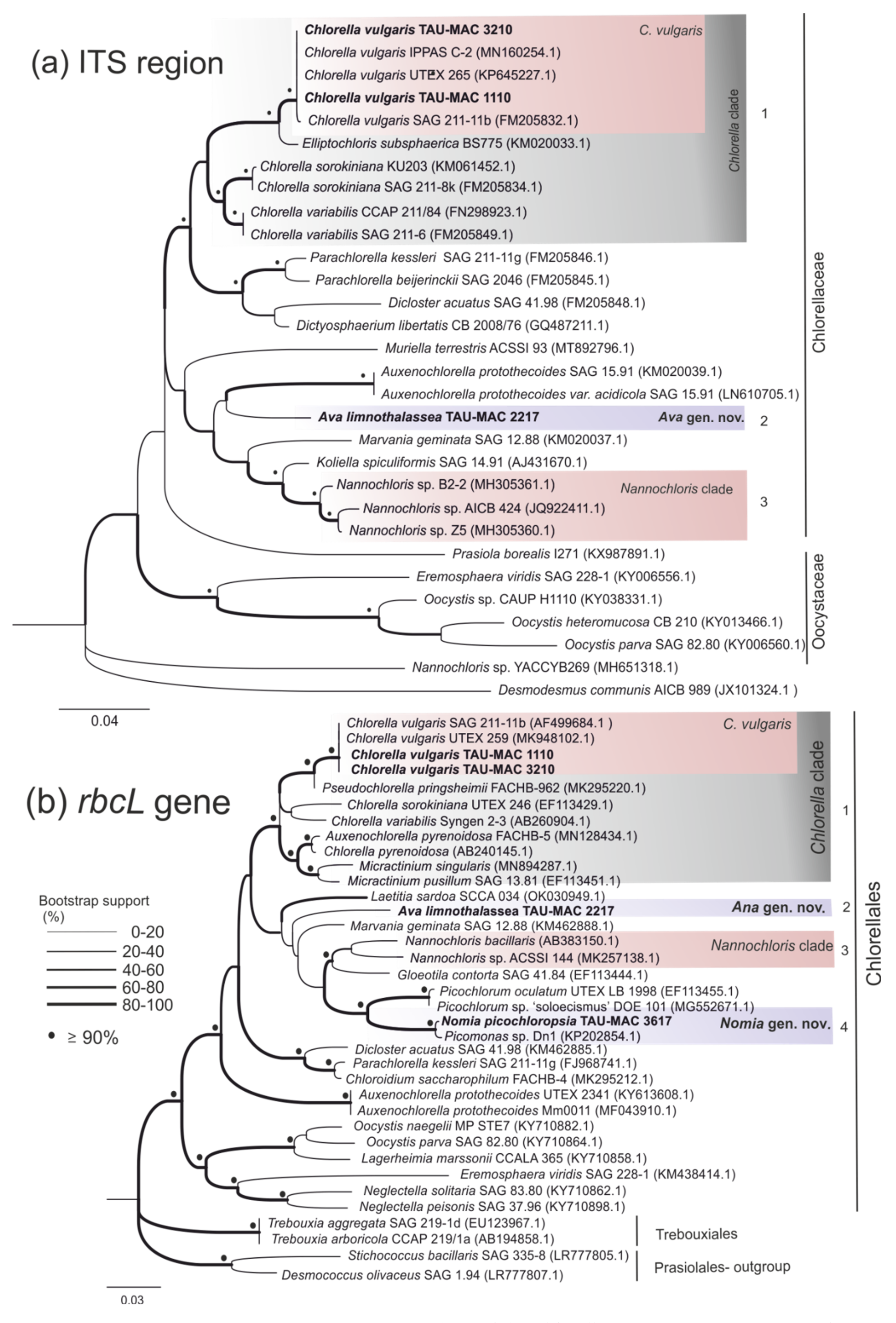
3.3. Chlorellales
3.4. Chlamydomonadales
3.5. Taxonomic Descriptions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; De Clerck, O. Phylogeny and Molecular Evolution of the Green Algae. CRC Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Fei, C.; Wang, C.; Gao, Z.; Bao, Y.; He, M.; Wang, C. How DNA Barcoding Can Be More Effective in Microalgae Identification: A Case of Cryptic Diversity Revelation in Scenedesmus (Chlorophyceae). Sci. Rep. 2016, 6, 36822. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.A.; McCourt, R.M. Green Algae and the Origin of Land Plants. Am. J. Bot. 2004, 91, 1535–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fučíková, K.; Lewis, P.O.; Lewis, L.a. Putting Incertae Sedis Taxa in Their Place: A Proposal for Ten New Families and Three New Genera in Sphaeropleales (Chlorophyceae, Chlorophyta). J. Phycol. 2014, 50, 14–25. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: https://www.algaebase.org (accessed on 5 September 2021).
- Lemieux, C.; Vincent, A.T.; Labarre, A.; Otis, C.; Turmel, M. Chloroplast Phylogenomic Analysis of Chlorophyte Green Algae Identifies a Novel Lineage Sister to the Sphaeropleales (Chlorophyceae) Phylogenetics and Phylogeography. BMC Evol. Biol. 2015, 15, 264. [Google Scholar] [CrossRef] [Green Version]
- Temraleeva, A.; Moskalenko, S.; Mincheva, E.; Bukin, Y.; Sinetova, M. Spongiosarcinopsis terrestris Gen. et Sp. Nov. (Chlorophyta, Chlorophyceae): A New Genus of Green Algae from Gray Forest Soil, Russia. Phytotaxa 2018, 376, 291–300. [Google Scholar] [CrossRef]
- Hoffman, R.; Wynne, M.J.; Schils, T.; Lopez-Bautista, J.; Verbruggen, H. Codium pulvinatum (Bryopsidales, Chlorophyta), a New Species from the Arabian Sea, Recently Introduced into the Mediterranean Sea. Phycologia 2018, 57, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.; Wolf, M.A.; Moro, I.; Sfriso, A.A.; Buosi, A.; Verlaque, M. Rediscovery of a Forgotten Mediterranean Chaetomorpha Species in the Venice Lagoon (North Adriatic Sea): Chaetomorpha stricta Schiffner (Cladophorales, Chlorophyta). Cryptogam. Algol. 2018, 39, 293–312. [Google Scholar] [CrossRef]
- Chemodanov, A.; Robin, A.; Jinjikhashvily, G.; Yitzhak, D.; Liberzon, A.; Israel, A.; Golberg, A. Feasibility Study of Ulva sp. (Chlorophyta) Intensive Cultivation in a Coastal Area of the Eastern Mediterranean Sea. Biofuels Bioprod. Biorefin. 2019, 13, 864–877. [Google Scholar] [CrossRef]
- Rindi, F.; Pasella, M.M.; Lee, M.F.E.; Verbruggen, H. Phylogeography of the Mediterranean Green Seaweed Halimeda tuna (Ulvophyceae, Chlorophyta). J. Phycol. 2020, 56, 1109–1113. [Google Scholar] [CrossRef]
- Lürling, M. Phenotypic Plasticity in the Green Algae Desmodesmus and Scenedesmus with Special Reference to the Induction of Defensive Morphology. Ann. Limnol.—Int. J. Limnol. 2003, 39, 85–101. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.D.; Lewis, L.A. An Assessment of Proposed DNA Barcodes in Freshwater Green Algae. Cryptogam. Algol. 2010, 31, 529–555. [Google Scholar]
- Barcytė, D.; Hodač, L.; Nedbalová, L. Lunachloris lukesovae Gen. et Sp. Nov. (Trebouxiophyceae, Chlorophyta), a Novel Coccoid Green Alga Isolated from Soil in South Bohemia, Czech Republic. Eur. J. Phycol. 2017, 52, 281–291. [Google Scholar] [CrossRef]
- Krienitz, L.; Bock, C. Present State of the Systematics of Planktonic Coccoid Green Algae of Inland Waters. Hydrobiologia 2012, 698, 295–326. [Google Scholar] [CrossRef]
- Singh, J.; Saxena, R.C. An Introduction to Microalgae: Diversity and Significance. In Handbook of Marine Microalgae; Elsevier: Amsterdam, The Netherlands, 2015; pp. 11–24. [Google Scholar]
- Moya, P.; Škaloud, P.; Chiva, S.; García-Breijo, F.J.; Reig-Armiñana, J.; Vančurová, L.; Barreno, E. Molecular Phylogeny and Ultrastructure of the Lichen Microalga Asterochloris mediterranea Sp. Nov. from Mediterranean and Canary Islands ecosystems. Int. J. Syst. Evol. Microbiol. 2015, 65, 1838–1854. [Google Scholar] [CrossRef]
- Lortou, U.; Gkelis, S. Polyphasic Taxonomy of Green Algae Strains Isolated from Mediterranean Freshwaters. J. Biol. Res. 2019, 26, 11. [Google Scholar] [CrossRef] [Green Version]
- Konstantinou, D.; Kakakiou, R.; Panteris, E.; Voultsiadou, E.; Gkelis, S. Photosynthetic Sponge-associated Eukaryotes in the Aegean Sea: A Culture-dependent Approach. J. Eukaryot. Microbiol. 2020, 67, 660–670. [Google Scholar] [CrossRef]
- Panou, M.; Gkelis, S. Unravelling Unknown Cyanobacteria Diversity Linked with HCN Production. Mol. Phylogenet. Evol. 2021, 166, 107322. [Google Scholar] [CrossRef]
- Barinova, S.S.; Anissimova, O.V.; Nevo, E.; Jarygin, M.M.; Wasser, S.P. Diversity and Ecology of Algae from the Nahal Qishon River, Northern Israel. Plant Biosyst. 2010, 138, 245–259. [Google Scholar] [CrossRef]
- Fadel, A.; Atoui, A.; Lemaire, B.J.; Vinçon-leite, B.; Slim, K. Environmental Factors Associated with Phytoplankton Succession in a Mediterranean Reservoir with a Highly Fluctuating Water Level. Environ. Monit. Assess. 2015, 187, 633. [Google Scholar] [CrossRef] [Green Version]
- Padedda, M.B.; Sechi, N.; Grazia, G.; Mariani, M.A.; Pulina, S.; Sarria, M.; Satta, T.C.; Virdis, T.; Buscarinu, P.; Luglie, A. Consequences of Eutrophication in the Management of Water Resources in Mediterranean Reservoirs: A Case Study of Lake Cedrino (Sardinia, Italy). Glob. Ecol. Conserv. 2017, 12, 21–35. [Google Scholar] [CrossRef]
- Chrisostomou, A.; Moustaka-Gouni, M.; Sgardelis, S.; Lanaras, T. Air-Dispersed Phytoplankton in a Mediterranean River-Reservoir System (Aliakmon-Polyphytos, Greece). J. Plankton Res. 2009, 31, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Panou, M.; Chronis, I.; Zervou, S.-K.; Christophoridis, C.; Manolidi, K.; Ntislidou, C.; Triantis, T.M.; Kaloudis, T.; Hiskia, A.; et al. Monitoring a Newly Re-Born Patient: Water Quality and Cyanotoxin Occurrence in a Reconstructed Shallow Mediterranean Lake. Adv. Oceanogr. Limnol. 2017, 8, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Genitsaris, S.; Kormas, K.A.; Moustaka-Gouni, M. Airborne Algae and Cyanobacteria: Occurrence and Related Health Effects. Front. Biosci. 2011, 3, 772–787. [Google Scholar]
- Elkomy, R.; Ibraheem, I.B.M.; Shreadah, M.; Mohammed, R.; Ismael, A. Antimicrobial Activity of Three Microalgae Isolated from Mediterranean Sea Coast, Egypt. J. Pure Appl. Microbiol. 2015, 9, 2751–2758. [Google Scholar]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giu, D.; Oulad, Y.; Majdoub, E.; Kounnoun, A.; Miceli, N.; Taviano, M.F.; et al. The Contribution of Carotenoids, Phenolic Compounds, and Flavonoids to the Antioxidative Properties of Marine Microalgae Isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haoujar, I.; Cacciola, F.; Manchado, M.; Abrini, J.; Haoujar, M.; Chebbaki, K.; Oteri, M.; Rigano, F.; Mangraviti, D.; Mondello, L.; et al. Isolation of Microalgae from Mediterranean Seawater and Production of Lipids in the Cultivated Species. Foods 2020, 9, 1601. [Google Scholar] [CrossRef]
- Psachoulia, P.; Schortsianiti, S.N.; Lortou, U.; Gkelis, S.; Chatzidoukas, C.; Samaras, P. Assessment of Nutrients Recovery Capacity and Biomass Growth of Four Microalgae Species in Anaerobic Digestion Effluent. Water 2022, 14, 221. [Google Scholar] [CrossRef]
- Legakis, A.; Constantinidis, T.; Petrakis, P.V. Biodiversity in Greece. In Global Biodiversity: Selected countries in Europe; Pullaiah, T., Ed.; Apple Academic Press: Palm Bay, FL, USA, 2018; Volume 2. [Google Scholar]
- Anastasiadis, E.; Seferlis, M.; Fitoka, E.; Mihalatu, E.; Papadimos, D.; Kastavouni, A.; Apostolakis, M.; Katsakiori, M.; Kazantzidis, S. Restoration and Conservation of the Kalohori Lagoon, Thessaloniki. In Proceedings of the 10th MEDECOS Conference, Rhodes, Greece, 25 April–1 May 2004. [Google Scholar]
- Malea, P.; Kevrekidis, T. Trace Element Patterns in Marine Macroalgae. Sci. Total Environ. 2014, 494–495, 144–157. [Google Scholar] [CrossRef]
- Gkelis, S.; Zaoutsos, N. Cyanotoxin Occurrence and Potentially Toxin Producing Cyanobacteria in Freshwaters of Greece: A Multi-Disciplinary Approach. Toxicon 2014, 78, 1–9. [Google Scholar] [CrossRef]
- Gkelis, S.; Fernández Tussy, P.; Zaoutsos, N. Isolation and Preliminary Characterization of Cyanobacteria Strains from Freshwaters of Greece. Open Life Sci. 2015, 10, 52–60. [Google Scholar] [CrossRef]
- Rippka, R. Isolation and Purification of Cyanobacteria. Methods Enzymol. 1988, 197, 3–27. [Google Scholar]
- Gkelis, S.; Panou, M. Capturing Biodiversity: Linking a Cyanobacteria Culture Collection to the “Scratchpads” Virtual Research Environment Enhances Biodiversity Knowledge. Biodivers. Data J. 2016, 4, e7965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattox, K.R.; Bold, H.C., III. Taxonomy of Certain Ulothrichacean Algae. In Phycological Studies; The University of Texas Publication: Austin, TX, USA, 1962. [Google Scholar]
- Lee, W.K. Uronema trentonense sp. nov. (Chlorophycophyta, Ulotrichales): A New Edaphic Alga from New Jersey. Phycologia 1978, 17, 191–195. [Google Scholar] [CrossRef]
- Ettl, H.; Chlorophyta, I. Phytomonadina. In Süwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Mollenhauer, D., Eds.; Gustav Fischer: Stuttgart, Germany, 1983; p. 807. [Google Scholar]
- Komárek, J.; Fott, B. Chlorophyceae (Grünalgen), Ordnung: Chlorococcales. In Das Phytoplankton des Süßwassers. Systematic und Biologie; Huber-Pestalozzi, G., Ed.; E. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller): Stuttgart, Germany, 1983; p. 1044. [Google Scholar]
- Ettl, H.; Gartner, G. Chlorophyta II. Tetrasporales, Chlorococcales, Gloeodendrales. In Süwasserflora von Mitteleuropa 10; Fischer: Stuttgart,Germany; New York, NY, USA, 1988. [Google Scholar]
- Hegewald, E.; Schnepf, E. Scenedesmus Abundans (Kirchn.) Chod., an Older Name for Chlorella fusca SHIH. et KRAUSS. Arch. Für Protistenkd. 1991, 139, 133–176. [Google Scholar] [CrossRef]
- Hegewald, E.H. New Combinations in the Genus Desmodesmus (Chlorophyceae, Scenedesmaceae). Arch. Hydrobiol. Suppl. Algol. Stud. 2000, 96, 1–18. [Google Scholar] [CrossRef]
- Hegewald, E.H.; Schmidt, A. Revision of the Desmodesmus Species with Lateral Spines. 1. Desmodesmus subspicatus (R.CHOD.) E. HEGEW. et A. SCHMIDT. Arch. Hydrobiol. Suppl. Algol. Stud. 2001, 101, 1–26. [Google Scholar]
- John, D.M.; Museum, N.H. The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae. Choice Rev. Online 2013, 49, 49–6880. [Google Scholar] [CrossRef]
- Turland, N.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Marhold, K.; May, T.W.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Regnum Vegetabile 159; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Atashpaz, S.; Khani, S.; Barzegari, A.; Barar, J.; Vahed, S.Z.; Azarbaijani, R.; Omidi, Y. A Robust Universal Method for Extraction of Genomic DNA from Bacterial Species. Microbiology 2010, 79, 538–542. [Google Scholar] [CrossRef]
- Maidak, B.L.; Cole, J.R.; Lilburn, T.G.; Parker, C.T.; Saxman, P.R.; Farris, R.J.; Garrity, G.M.; Olsen, G.J.; Schmidt, T.M.; Tiedje, J.M. The RDP-II (Ribosomal Database Project). Nucleic Acids Res. 2001, 29, 173–174. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information—GenBank. Available online: www.ncbi.nlm.nih.gov (accessed on 5 August 2021).
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Vanormelingen, P.; Hegewald, E.; Braband, A.; Kitschke, M.; Friedl, T.; Sabbe, K.; Vyverman, W. The Systematics of a Small Spineless Desmodesmus Species, D. costato-granulatus (Sphaeropleales, Chlorophyceae), Based on Its2 RDNA Sequence Analyses and Cell Wall Morphology. J. Phycol. 2007, 43, 378–396. [Google Scholar] [CrossRef]
- Egan, P.F.; Trainor, F.R. Phenotypic Plasticity in Scenedesmus communis Hegew. (Chlorophyceae). II. Examples of Algal Cyclo- and Nonocyclomorphosis. Am. J. Bot. 1991, 7, 37–52. [Google Scholar] [CrossRef]
- An, S.S.; Friedel, T.; Hegewald, E. Phylogenetic Relationships of Scenedesmus and Scenedesmus-like Coccoid Green Algae as Inferred from ITS-2 RDNA Sequence Comparison. Plant Biol. 1999, 1, 418–428. [Google Scholar] [CrossRef]
- Shubert, E.; Wilk-Woźniak, E.; Ligęza, S. An Autecological Investigation of Desmodesmus: Implications for Ecology and Taxonomy. Plant Ecol. Evol. 2014, 147, 202–212. [Google Scholar] [CrossRef]
- De Cambiaire, J.C.; Otis, C.; Lemieux, C.; Turmel, M. The Complete Chloroplast Genome Sequence of the Chlorophycean Green Alga Scenedesmus obliquus Reveals a Compact Gene Organization and a Biased Distribution of Genes on the Two DNA Strands. BMC Evol. Biol. 2006, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Wynne, M.J.; Hallan, J.K. Reinstatement of Tetradesmus G. M. Smith (Sphaeropleales, Chlorophyta). Feddes Repert. 2016, 126, 83–86. [Google Scholar] [CrossRef]
- Hegewald, E.; Braband, A. A Taxonomic Revision of Desmodesmus Serie Desmodesmus (Sphaeropleales, Scenedesmaceae). J. Czech Phycol. 2017, 17, 191–208. [Google Scholar] [CrossRef]
- Tsarenko, P.M.; Hegewald, E.; Braband, A. Scenedesmus-like Algae of Ukraine. 1. Diversity of Taxa from Water Bodies in Volyn Polissia. Arch. Hydrobiol. Suppl. Algol. Stud. 2005, 118, 1–45. [Google Scholar] [CrossRef]
- Gorelova, O.A.; Baulina, O.I.; Solovchenko, A.E.; Chekanov, K.A.; Chivkunova, O.B.; Fedorenko, T.A.; Lobakova, E.S. Similarity and Diversity of the Desmodesmus Spp. Microalgae Isolated from Associations with White Sea Invertebrates. Protoplasma 2014, 252, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Mandotra, S.K.; Kumar, P.; Suseela, M.R.; Ramteke, P.W. Scenedesmus abundans: A Potential Feedstock for High Quality Biodiesel Production. Bioresour. Technol. 2014, 156, 42–47. [Google Scholar] [CrossRef]
- Hegewald, E.; Wolf, M.; Keller, A.; Friedl, T.; Krienitz, L. ITS2 Sequence-Structure Phylogeny in the Scenedesmaceae with Special Reference to Coelastrum (Chlorophyta, Chlorophyceae), Including the New Genera Comasiella and Pectinodesmus. Phycologia 2010, 49, 325–335. [Google Scholar] [CrossRef]
- Hong, J.-W.; Kim, S.-A.; Chang, J.-W.; Yi, J.; Jeong, J.-E.; Kim, S.-H.; Kim, S.-H.; Yoon, H.-S. Isolation and Description of a Korean Microalga, Asterarcys quadricellulare KNUA020, and Analysis of Its Biotechnological Potential. Algae 2012, 27, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Saber, A.A.; Fučíková, K.; McManus, H.A.; Guella, G.; Cantonati, M. Novel Green Algal Isolates from the Egyptian Hyper-Arid Desert Oases: A Polyphasic Approach with a Description of Pharao desertorum Gen. et Sp. Nov. (Chlorophyceae, Chlorophyta). J. Phycol. 2018, 54, 342–357. [Google Scholar] [CrossRef]
- Kaufnerová, V.; Eliáš, M. The Demise of the Genus Scotiellopsis Vinatzer (Chlorophyta). Nov. Hedwig. 2013, 97, 415–428. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Song, H.; Liu, X.; Liu, B.; Hu, Z.; Liu, G. Morphology and Molecular Phylogeny of Coccoid Green Algae Coelastrella sensu lato (Scenedesmaceae, Sphaeropeales), Including the Description of Three New Species and Two New Varieties. J. Phycol. 2019, 55, 1290–1305. [Google Scholar] [CrossRef]
- Comas Gonzales, A. New Coenobial Chlorococcales (Chlorophyceae) from Cuba. Arch. Hydrobiol. Algol. Stud. 1981, 28, 213–219. [Google Scholar]
- Da Silva, T.G.; Bock, C.; Sant’Anna, C.L.; Bagatini, I.L.; Wodniok, S.; Vieira, A.A.H. Selenastraceae (Sphaeropleales, Chlorophyceae): RbcL, 18S RDNA and ITS–2 Secondary Structure Enlightens Traditional Taxonomy, with Description of Two New Genera, Messastrum Gen. Nov. and Curvastrum Gen. Nov. Fottea 2017, 17, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Krienitz, L.; Ustinova, I.; Friedl, T.; Huss, V.A.R. Traditional Generic Concepts versus 18S RRNA Gene Phylogeny in the Green Algal Family Selenastraceae (Chlorophyceae, Chlorophyta). J. Phycol. 2001, 37, 852–865. [Google Scholar] [CrossRef] [Green Version]
- Krienitz, L.; Bock, C.; Nozaki, H.; Wolf, M. Ssu RRNA Gene Phylogeny of Morphospecies Affiliated to the Bioassay Alga “Selenastrum capricornutum” Recovered the Polyphyletic Origin of Crescent-Shaped Chlorophyta. J. Phycol. 2011, 47, 880–893. [Google Scholar] [CrossRef]
- Fawley, M.W.; Dean, M.L.; Dimmer, S.K.; Fawley, K.P. Evaluating the Morphospecies Concept in the Selenastraceae (Chlorophyceae, Chlorophyta). J. Phycol. 2006, 42, 142–154. [Google Scholar] [CrossRef]
- Komarkova-Legnerova, J. The Systematics and Ontogenesis of the Genera Ankistrodesmus Corda and Monoraphidium Gen. Nov. In Studies in Phycology; Academia: Praha, Czech Republic, 1969; pp. 75–122. [Google Scholar]
- Krienitz, L.; Klein, G. Morphologie Und Ultrastruktur Einiger Arten Der Gattung Monoraphidium III. Arch. Hydrobiol. 1988, 78, 447–463. [Google Scholar]
- Da Silva, D.; Sant’Anna, C.L.; Tucci, A.; Comas, A. New Planktic Species of Kirchneriella Schmidle (Chlorophyceae, Selenastraceae) from Brazilian Freshwaters. Braz. J. Bot. 2013, 36, 153–157. [Google Scholar] [CrossRef]
- Gkelis, S.; Papadimitriou, T.; Zaoutsos, N.; Leonardos, I. Anthropogenic and Climate-Induced Change Favors Toxic Cyanobacteria Blooms: Evidence from Monitoring a Highly Eutrophic, Urban Mediterranean Lake. Harmful Algae 2014, 39, 322–333. [Google Scholar] [CrossRef]
- Leliaert, F.; Rueness, J.; Boedeker, C.; Maggs, C.A.; Cocquyt, E.; Verbruggen, H.; De Clerck, O. Systematics of the Marine Microfilamentous Green Algae Uronema curvatum and Urospora microscopica (Chlorophyta). Eur. J. Phycol. 2009, 44, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nozaki, H.; Miyazawa, Y.; Koide, T.; Kawano, S. Relationship between Presence of a Mother Cell Wall and Speciation in the Unicellular Microalga Nannochloris (Chlorophyta). J. Phycol. 2003, 39, 172–184. [Google Scholar] [CrossRef]
- Sumiya, N.; Owari, S.; Watanabe, K.; Kawano, S. Role of Multiple Ftsz Rings in Chloroplast Division under Oligotrophic and Eutrophic Conditions in the Unicellular Green Alga Nannochloris bacillaris (Chlorophyta, Trebouxiophyceae). J. Phycol. 2012, 48, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Yamamoto, M.; Sakamoto, W.; Kawano, S. Isolation and Molecular Characterization of RbcS in the Unicellular Green Alga Nannochloris bacillaris (Chlorophyta, Trebouxiophyceae). Phycol. Res. 2005, 53, 67–76. [Google Scholar] [CrossRef]
- Naumann, E. Notizen zur Systematik Der Süsswasseralgen. Ark. För Bot. 1919, 16, 1–19. [Google Scholar]
- Henley, W.J.; Hironaka, J.L.; Guillou, L.; Buchheim, M.A.; Buchheim, J.A.; Fawley, M.W.; Fawley, K.P. Phylogenetic Analysis of the “Nannochloris-like” Algae and Diagnoses of Picochlorum oklahomensis Gen. et Sp Nov (Trebouxiophyceae, Chlorophyta). Phycologia 2004, 43, 641–652. [Google Scholar] [CrossRef]
- Malavasi, V.; Škvorová, Z.; Němcová, Y.; Škaloud, P. Laetitia sardoa gen. & sp. nov., a new member of the Chlorellales (Trebouxiophyceae, Chlorophyta) isolated from Sardinia Island. Phycologia 2022, 61, 375–383. [Google Scholar] [CrossRef]
- Seenivasan, R.; Sausen, N.; Medlin, L.K.; Melkonian, M. Picomonas judraskeda Gen. Et Sp. Nov.: The First Identified Member of the Picozoa Phylum Nov., a Widespread Group of Picoeukaryotes, Formerly Known as “Picobiliphytes”. PLoS ONE 2013, 8, e59565. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Nelson, D.R.; Abdrabu, R.; Khraiwesh, B.; Jijakli, K.; Arnoux, M.; O’Connor, M.J.; Bahmani, T.; Cai, H.; Khapli, S.; et al. An Integrative Raman Microscopy-Based Workflow for Rapid in Situ Analysis of Microalgal Lipid Bodies. Biotechnol. Biofuels 2015, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Qiao, K.; Takano, T.; Liu, S. Discovery of Two Novel Highly Tolerant NaHCO3 Trebouxiophytes: Identification and Characterization of Microalgae from Extreme Saline-Alkali Soil. Algal Res. 2015, 9, 245–253. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Nakada, T.; Tomita, M. Taxonomic Revision of Oil-Producing Green Algae, Chlorococcum oleofaciens (Volvocales, Chlorophyceae), and Its Relatives. J. Phycol. 2015, 51, 1000–1016. [Google Scholar] [CrossRef]
- Temraleeva, A.D.; Moslalenko, S. V Application of Morphological and Molecular Systematics for Identification of Green Microalgae of the Genus Chlorococcum and Some Closely Related Taxa. Microbiology 2019, 88, 27–38. [Google Scholar] [CrossRef]
- Yumoto, K.; Kasai, F.; Kawachi, M. Taxonomic Re-Examination of Chlamydomonas Strains Maintained in the NIES-Collection. Microbiol. Cult. Collect. 2013, 29, 1–12. [Google Scholar]
- Ettl, H.; Komarek, J. Was Versteht Man Unter Dem Begriff “Coccale Grünalgen”? Arch. Hydrobiol. Suppl. 1982, 60, 345–374. [Google Scholar]
- Friedl, T. Evolution of the Polyphyletic Genus Pleurastrum (Chlorophtya): Inferences from Nuclear—Encoded Ribosomal DNA Sequences and Motile Cell Ultr Astructure. Phycologia 1996, 35, 456–469. [Google Scholar] [CrossRef]
- Katana, A.; Kwiatowski, J.; Spalik, K.; Zakrys, B. Phylogenetic Position of Koliella (Chlorophyta) as Inferred from Nuclear and Chloroplast Small Subunit RDNA. J. Phycol. 2001, 37, 443–451. [Google Scholar] [CrossRef]
- Moro, C.V.; Crouzet, O.; Rasconi, S.; Thouvenot, A.; Coffe, G.; Batisson, I.; Bohatier, J. New Design Strategy for Development of Specific Primer Sets for PCR-Based Detection of Chlorophyceae and Bacillariophyceae in Environmental Samples. Appl. Environ. Microbiol. 2009, 75, 5729–5733. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Affan, M.A.; Jang, J.; Kang, M.H.; Ko, A.R.; Jeon, S.M.; Oh, C.; Heo, S.J.; Lee, Y.H.; Ju, S.J.; et al. Morphological, Molecular, and Biochemical Characterization of Astaxanthin-Producing Green Microalga Haematococcus Sp. KORDI03 (Haematococcaceae, Chlorophyta) Isolated from Korea. J. Microbiol. Biotechnol. 2015, 25, 238–246. [Google Scholar] [CrossRef] [Green Version]
- McManus, H.A.; Lewis, L.A. Molecular Phylogenetic Relationships in the Freshwater Family Hydrodictyaceae (Shaeropleales, Chlorophyceae), with an Emphasis on Pediastrum Duplex. J. Phycol. 2011, 47, 152–163. [Google Scholar] [CrossRef]
- Hoham, R.W.; Bonome, T.A.; Martin, C.W.; Leebens-Mack, J.H. A Combined 18S RDNA and RbcL Phylogenetic Analysis of Chloromonas and Chlamydomonas (Chlorophyceae, Volvocales) Emphasizing Snow and Other Cold-Temperature Habitats. J. Phycol. 2002, 38, 1051–1064. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | Geographic Coordinates | Habitat | Collection Date | |
|---|---|---|---|---|---|
| (N) | (E) | ||||
| Desmodesmus sp. TAU-MAC 1817 | Agkistro Hot Springs | 41°22′04″ | 23°25′40″ | planktic | 17 October 2017 |
| Desmodesmus multivariabilis TAU-MAC 1917 | planktic | ||||
| Desmodesmus sp. TAU-MAC 2017 | planktic | ||||
| Desmodesmus abundans TAU-MAC 2117 | planktic | ||||
| Desmodesmus multivariabilis TAU-MAC 2517 | planktic | ||||
| Desmodesmus sp. TAU-MAC 2617 | planktic | ||||
| Desmodesmus abundans TAU-MAC 2717 | planktic | ||||
| Ava limnothalassea TAU-MAC 2217 | Kalochori Lagoon | 40°27′53″ | 22°51′48″ | planktic | 1 November 2017 |
| Nomia picochloropsia TAU-MAC 3617 | planktic | ||||
| Asterarcys quadricellulare TAU-MAC 3917 | AUTH water pond | 40°37′58″ | 22°57′21″ | planktic | 1 November 2017 |
| Uronema trentonense TAU-MAC 0215 | Agkistro Hot Springs | 41°22′04″ | 23°25′40″ | benthic | 20 October 2015 |
| Desmodesmus subspicatus TAU-MAC 0415 | planktic | ||||
| Akraea chliaropsychia TAU-MAC 0515 | planktic | ||||
| Spongiosarcinopsis limneus TAU-MAC 3310 | Lake Doirani | 41°18′56″ | 22°45′37″ | planktic | 21 August 2010 |
| Desmodesmus sp. TAU-MAC 1010 | Lake Koronia | 40°42′04″ | 23°08′17″ | planktic | 30 August 2010 |
| Lilaea pamvotia TAU-MAC 3510 | Lake Pamvotis | 39°40′51″ | 20°50′30″ | planktic | 1 November 2010 |
| Monoraphidium sp. TAU-MAC 1210 | planktic | ||||
| Desmodesmus sp. TAU-MAC 0910 | Lake Volvi | 40°40′37″ | 23°33′10″ | planktic | 13 July 2010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lortou, U.; Panteris, E.; Gkelis, S. Uncovering New Diversity of Photosynthetic Microorganisms from the Mediterranean Region. Microorganisms 2022, 10, 1571. https://doi.org/10.3390/microorganisms10081571
Lortou U, Panteris E, Gkelis S. Uncovering New Diversity of Photosynthetic Microorganisms from the Mediterranean Region. Microorganisms. 2022; 10(8):1571. https://doi.org/10.3390/microorganisms10081571
Chicago/Turabian StyleLortou, Urania, Emmanuel Panteris, and Spyros Gkelis. 2022. "Uncovering New Diversity of Photosynthetic Microorganisms from the Mediterranean Region" Microorganisms 10, no. 8: 1571. https://doi.org/10.3390/microorganisms10081571