
First Report of Two Jaculus Rodents as Potential Reservoir Hosts of Leishmania Parasites in Tunisia

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rodent Trapping and Identification
2.2. Detection of Infection by Leishmania Parasites
2.2.1. Clinical Examination
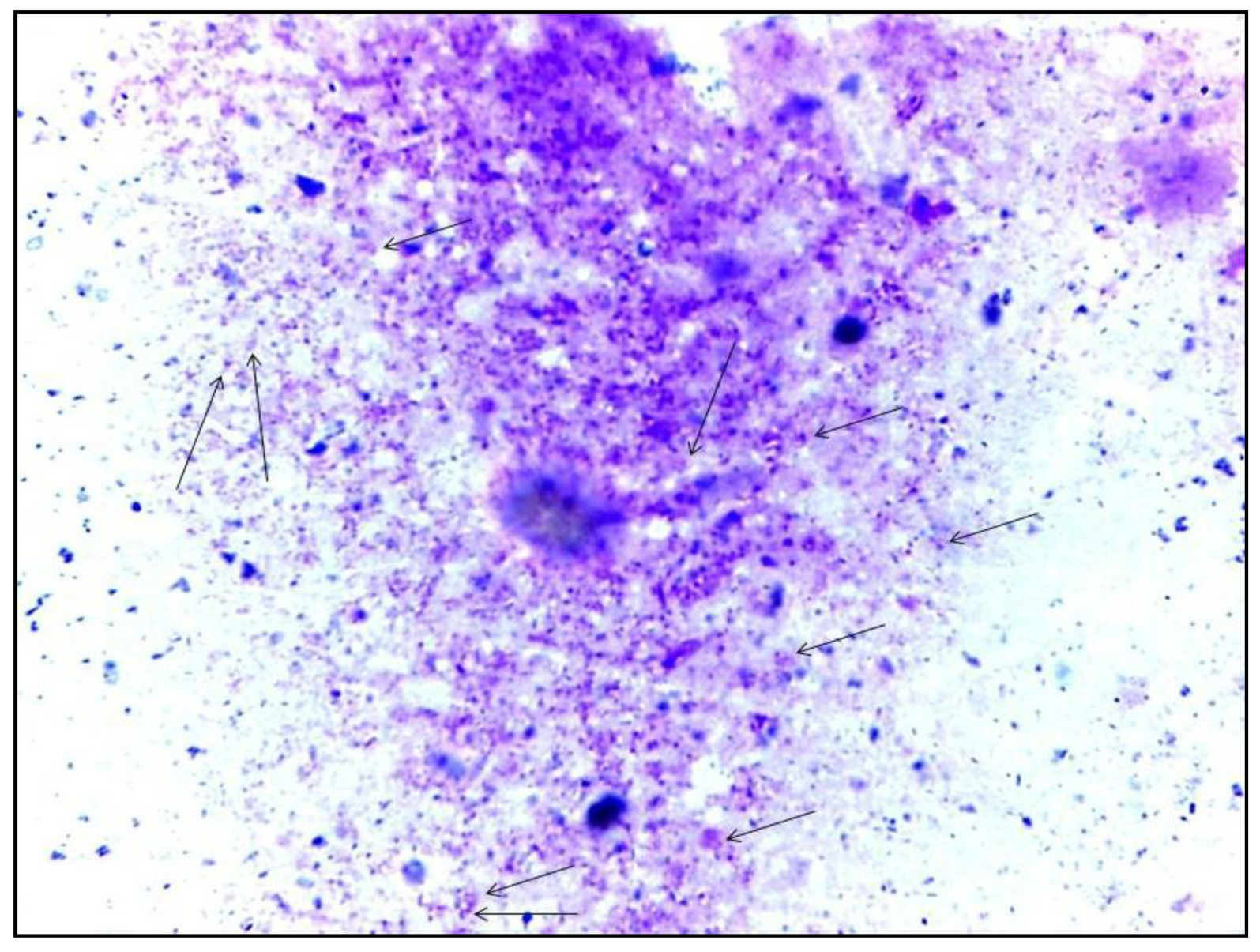
2.2.2. Parasitological Diagnosis
2.2.3. Molecular Screening
2.2.4. Leishmania Parasite Typing
2.2.5. Collection of Publicly Available Sequences and Phylogenetic Analyses
2.3. Statistical Analysis
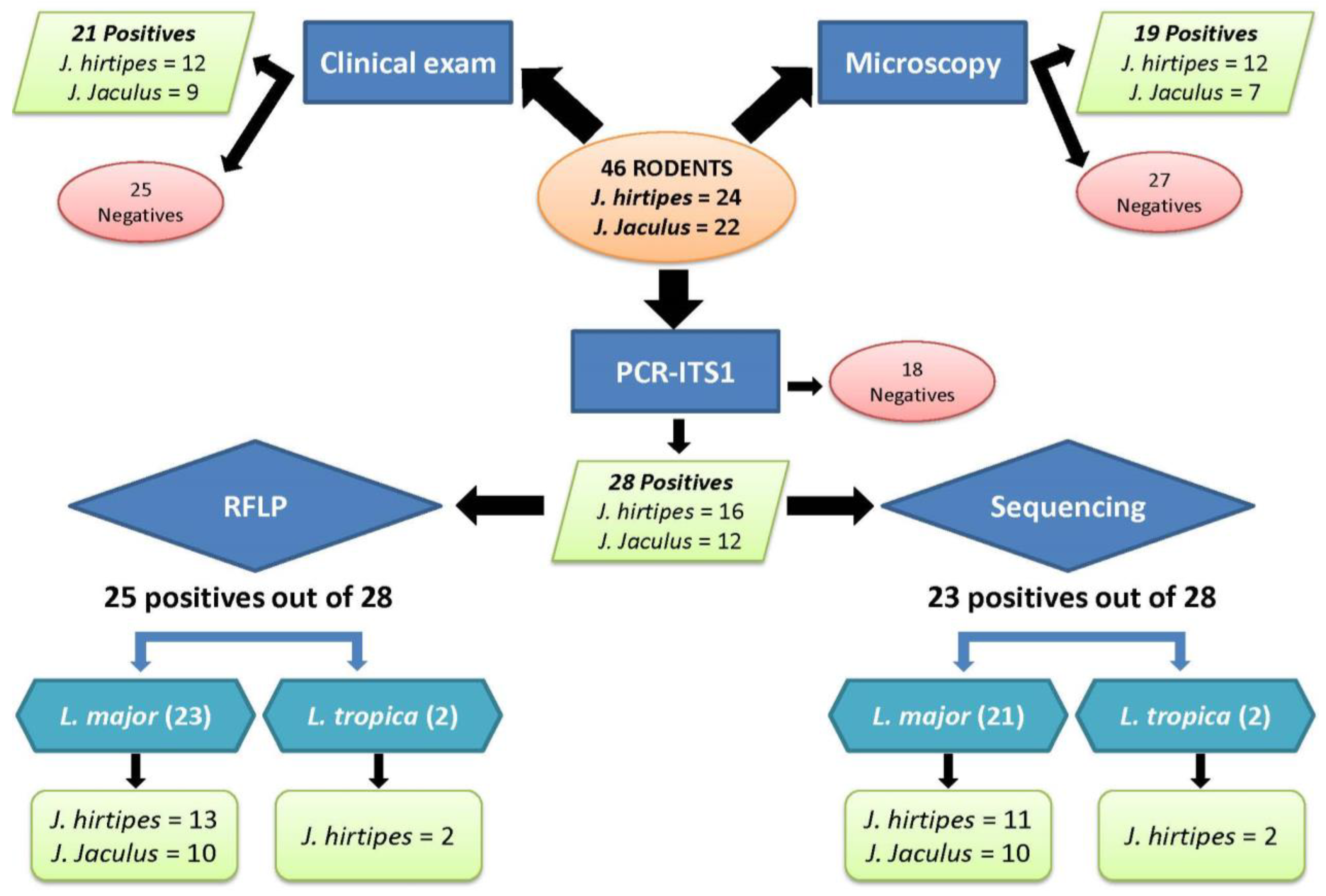
3. Results
3.1. Rodent Identification
3.2. Leishmania Infection
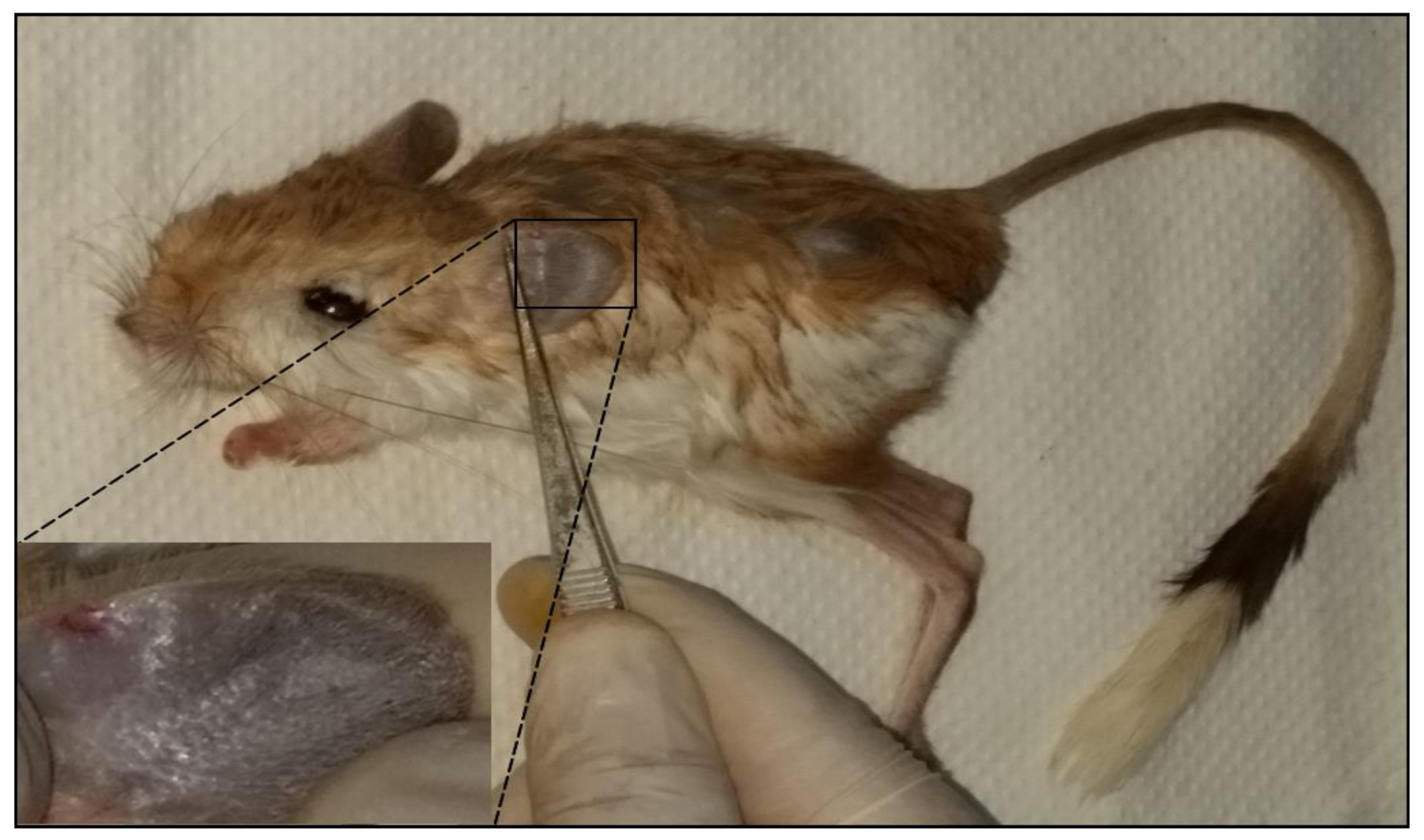
3.3. Clinical Manifestations
3.4. Leishmania Infection Prevalence
3.4.1. Parasitological Methods
3.4.2. Molecular Methods
3.4.3. Combined Methods
3.5. Leishmania Parasite Typing
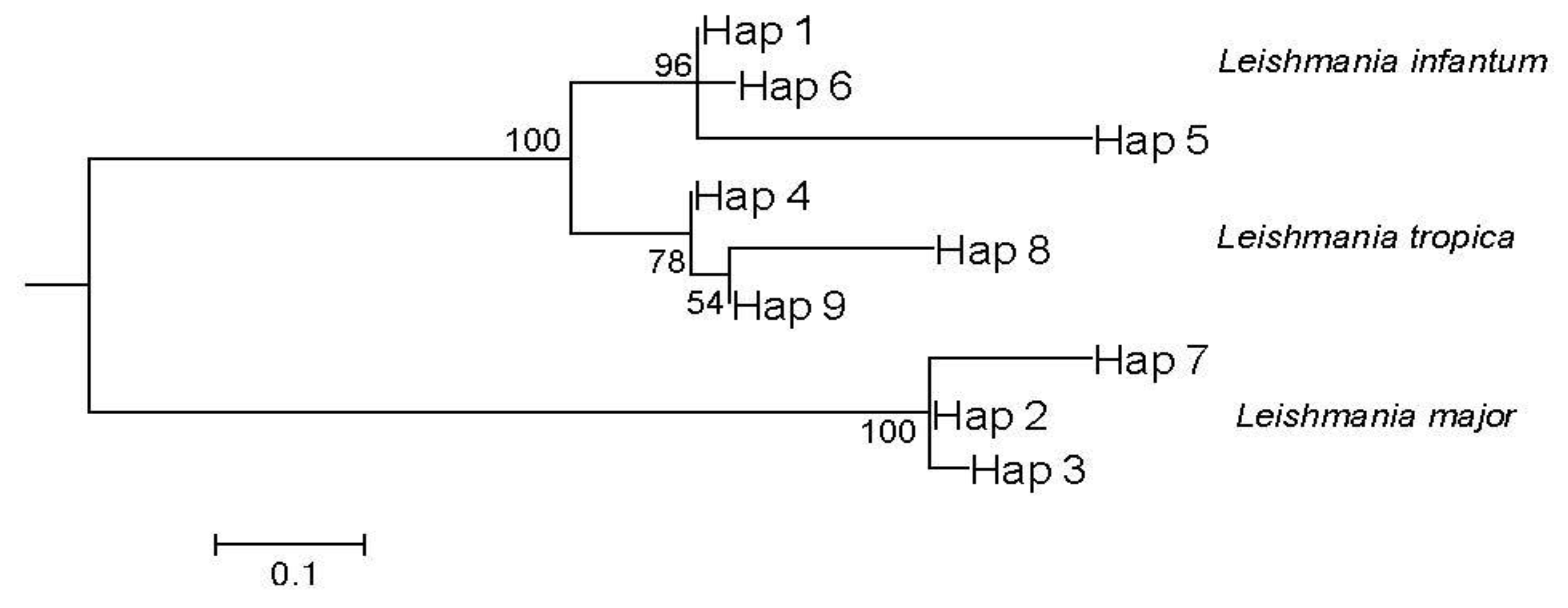
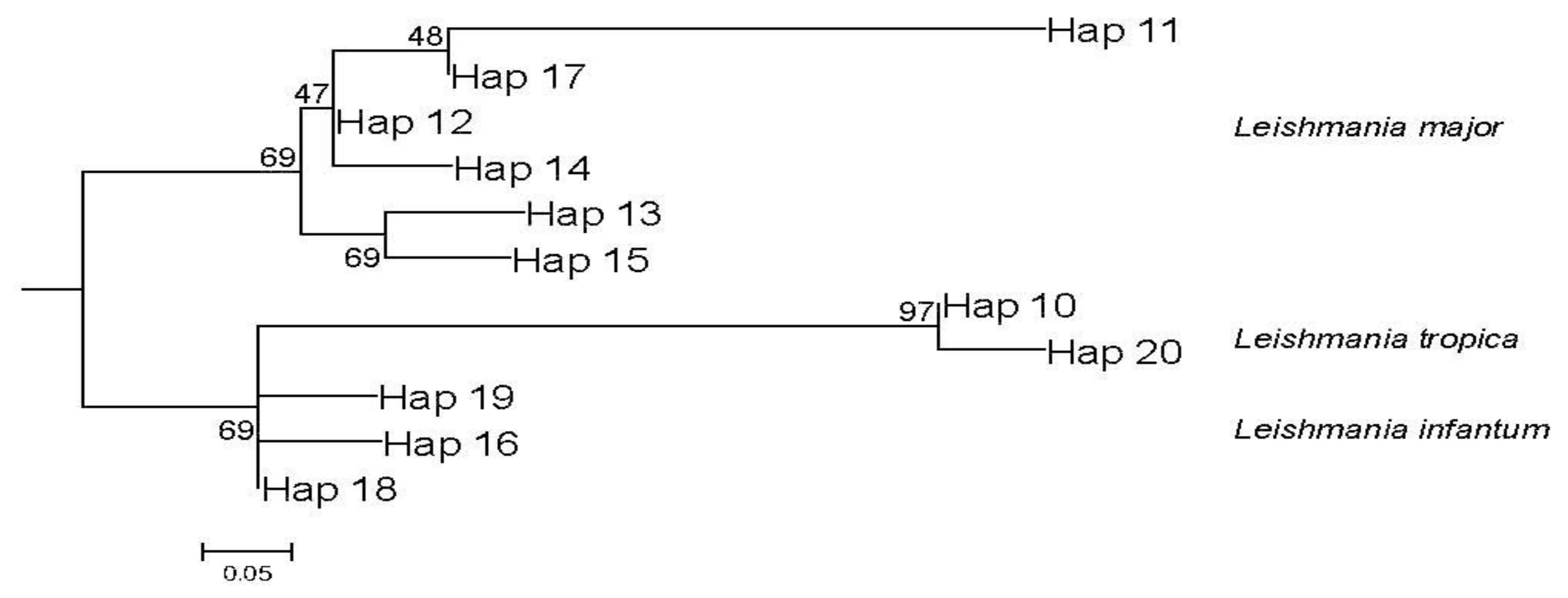
3.6. Leishmania Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klaus, S.N.; Frankenburg, S.; Ingber, A. Epidemiology of cutaneous leishmaniasis. Clin. Dermatol. 1999, 17, 257–260. [Google Scholar] [CrossRef]
- Aoun, K.; Bouratbine, A. Cutaneous leishmaniasis in North Africa: A review. Parasite 2014, 21, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoun, K.; Amri, F.; Chouihi, E.; Haouas, N.; Bedoui, K.; Benikhlef, R.; Ghrab, J.; Babba, H.; Chahed, M.K.; Harrat, Z.; et al. Epidemiology of Leishmania (L.) infantum, L. major and L. killicki in Tunisia: Results and analysis of the identification of 226 human and canine isolates. Bull. Soc. Pathol. Exot. 2008, 101, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Masmoudi, A.; Ayadi, N.; Boudaya, S.; Meziou, T.J.; Mseddi, M.; Marrekchi, S.; Bouassida, S.; Turki, H.; Zahaf, A. Clinical polymorphism of cutaneous leishmaniasis in centre and south of Tunisia. Bull. Soc. Pathol. Exot. 2007, 100, 36–40. [Google Scholar]
- Ben Ismail, R.; Gradoni, L.; Gramiccia, M.; Bettini, S.; Ben Rachid, M.S.; Garraoui, A. Epidemic cutaneous leishmaniasis in Tunisia: Biochemical characterization of parasites. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 669–670. [Google Scholar] [CrossRef]
- Benikhlef, R.; Harrat, Z.; Toudjine, M.; Djerbouh, A.; Bendali-Braham, S.; Belkaid, M. Detection of Leishmania infantum MON-24 in the dog. Med. Trop. 2004, 64, 381–383. [Google Scholar]
- Ghawar, W.; Snoussi, M.A.; Hamida, N.B.; Boukthir, A.; Yazidi, R.; Chaabane, S.; Chemkhi, J.; Zaatour, A.; Salah, A.B. First report of natural infection of least weasel (Mustela nivalis Linnaeus, 1776) with Leishmania major in Tunisia. Vector Borne Zoonotic Dis. 2011, 11, 1507–1509. [Google Scholar] [CrossRef]
- Bousslimi, N.; Ben-Ayed, S.; Ben-Abda, I.; Aoun, K.; Bouratbine, A. Natural infection of North African gundi (Ctenodactylus gundi) by Leishmania tropica in the focus of cutaneous leishmaniasis, Southeast Tunisia. Am. J. Trop. Med. Hyg. 2012, 86, 962–965. [Google Scholar] [CrossRef] [Green Version]
- Ghawar, W.; Bettaieb, J.; Salem, S.; Snoussi, M.A.; Jaouadi, K.; Yazidi, R.; Ben-Salah, A. Natural infection of Ctenodactylus gundi by Leishmania major in Tunisia. Acta Trop. 2018, 177, 89–93. [Google Scholar] [CrossRef]
- Chemkhi, J.; Souguir, H.; Ali, I.B.; Driss, M.; Guizani, I.; Guerbouj, S. Natural infection of Algerian hedgehog, Atelerix algirus (Lereboullet 1842) with Leishmania parasites in Tunisia. Acta Trop. 2015, 150, 42–51. [Google Scholar] [CrossRef]
- Gharaibeh, B.M. Systematics, Distribution, and Zoogeography of Mammals of Tunisia; Texas Tech University: Lubbock, TX, USA, 1997. [Google Scholar]
- Ghawar, W.; Chaouch, M.; Ben Abderrazak, S.; Snoussi, M.A.; Salem, S.; Chouchen, S.; Bouaoun, A.; Ben Salah, A.; Bettaieb, J. Evaluation of the Taxonomic Status of Lesser Egyptian Jerboa, Jaculus jaculus: First Description of New Phylogroups in Tunisia. Animals 2022, 12, 758. [Google Scholar] [CrossRef]
- Svobodova, M.; Alten, B.; Zidkova, L.; Dvorak, V.; Hlavackova, J.; Myskova, J.; Seblova, V.; Kasap, O.E.; Belen, A.; Votypka, J.; et al. Cutaneous leishmaniasis caused by Leishmania infantum transmitted by Phlebotomus tobbi. Int. J. Parasit. 2009, 39, 251–256. [Google Scholar] [CrossRef]
- Njiru, Z.K.; Constantine, C.C.; Guya, S.; Crowther, J.; Kiragu, J.M.; Thompson, R.C.; Davila, A.M. The use of ITS1 rDNA PCR in detecting pathogenic African trypanosomes. Parasitol. Res. 2005, 95, 186–192. [Google Scholar] [CrossRef]
- Schonian, G.; Nasereddin, A.; Dinse, N.; Schweynoch, C.; Schallig, H.D.; Presber, W.; Jaffe, C.L. PCR diagnosis and characterization of Leishmania in local and imported clinical samples. Diagn. Microbiol. Infect. Dis. 2003, 47, 349–358. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Ben Othman, S.; Ghawar, W.; Chaouch, M.; Ayari, C.; Chemkhi, J.; Cancino-Faure, B.; Tomas-Perez, M.; Alcover, M.M.; Riera, C.; Ben Salah, A.; et al. First detection of Leishmania DNA in Psammomys obesus and Psammomys vexillaris: Their potential involvement in the epidemiology of leishmaniasis in Tunisia. Infect. Genet. Evol. 2018, 59, 7–15. [Google Scholar] [CrossRef]
- Ben-Ismail, R.; Ben Rachid, M.S.; Gradoni, L.; Gramiccia, M.; Helal, H.; Bach-Hamba, D. Zoonotic cutaneous leishmaniasis in Tunisia: Study of the disease reservoir in the Douara area. Ann. Soc. Belg. Med. Trop. 1987, 67, 335–343. [Google Scholar] [PubMed]
- Ben-Ismail, R.; Khaled, S.; Makni, S.; Ben Rachid, M.S. Anti-leishmanial antibodies during natural infection of Psammomys obesus and Meriones shawi (Rodentia, Gerbillinae) by Leishmania major. Ann. Soc. Belg. Med. Trop. 1989, 69, 35–40. [Google Scholar] [PubMed]
- Davami, M.H.; Motazedian, M.H.; Kalantari, M.; Asgari, Q.; Mohammadpour, I.; Sotoodeh-Jahromi, A.; Solhjoo, K.; Pourahmad, M. Molecular Survey on Detection of Leishmania Infection in Rodent Reservoirs in Jahrom District, Southern Iran. J. Arthropod. Borne Dis. 2014, 8, 139–146. [Google Scholar] [PubMed]
- Echchakery, M.; Chicharro, C.; Boussaa, S.; Nieto, J.; Carrillo, E.; Sheila, O.; Moreno, J.; Boumezzough, A. Molecular detection of Leishmania infantum and Leishmania tropica in rodent species from endemic cutaneous leishmaniasis areas in Morocco. Parasit. Vectors 2017, 10, 454. [Google Scholar] [CrossRef] [Green Version]
- Parvizi, P.; Moradi, G.; Akbari, G.; Farahmand, M.; Ready, P.D.; Piazak, N.; Assmar, M.; Amirkhani, A. PCR detection and sequencing of parasite ITS-rDNA gene from reservoirs host of zoonotic cutaneous leishmaniasis in central Iran. Parasitol. Res. 2008, 103, 1273–1278. [Google Scholar] [CrossRef]
- Faleh, A.B.; Annabi, A.; López, S.; Said, K.; Ribas, A. On the helminth parasites of the genus Jaculus (Rodentia: Dipodidae) in Tunisia: A preliminary survey study. Leb. Sci. J. 2012, 13, 1107. [Google Scholar]
- Hoogstraal, H. The life cycle and incidence of Hepatozoon balfouri (Laveran, 1905) in Egyptian jerboas (Jaculus spp.) and mites (Haemolaelaps aegyptius Keegan, 1956). J. Protozool. 1961, 8, 231–248. [Google Scholar] [CrossRef]
- Morsy, T.A.; el Bahrawy, A.A.; al Dakhil, M.M.; Abdel Mawla, M.M. Babesia infection in rodents trapped in Riyadh Region, Saudi Arabia, with a general discussion. J. Egypt. Soc. Parasitol. 1994, 24, 177–185. [Google Scholar]
- Al-Taqi, M.; Mohammed, A.H. Susceptibility of Kuwaiti rodents and experimental mice to isolates of Leishmania spp. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 133–136. [Google Scholar] [CrossRef]
- Archibald, R. A Preliminary Report on some Further Investigations on Kala Azar in the Sudan. BMJ Mil. Health 1914, 23, 479–495. [Google Scholar]
- Dedet, P.; Ben Osman, F. Sensibilite experimentale de divers rongeurs de Tunisie vis-a-vis de souches autochtones de Leishmania tropica (Wright, 1903) et de Leishmania donovani (Laveran & Mesnil, 1903). Arch. Inst. Pasteur Tunis 1973, 50, 85–94. [Google Scholar]
- Bin, D.S.; Mostafa, O.M.; Abdoon, A.; Al-Quraishy, S.A.; Alqahtani, A.A. Isoenzyme electrophetic characterization of Leishmania major, the causative agent of zoonotic cutaneous Leishmaniasis in North and West Saudi Arabia. J. Egypt. Soc. Parasitol. 2010, 40, 465–478. [Google Scholar]
- Hamadto, H.A.; Al, F.A.; Farrag, A.B.; Abdel Maksoud, M.K.; Morsy, T.A. Zoonotic cutaneous leishmaniasis: Reservoir host and insect vector in north Sinai, Egypt. J. Egypt. Soc. Parasitol. 2007, 37, 843–850. [Google Scholar]
- Morsy, T.A.; Naser, A.M.; el Gibali, M.R.; Anwar, A.M.; el Said, A.M. Studies on zoonotic cutaneous leishmaniasis among a group of temporary workers in North Sinai Governorate, Egypt. J. Egypt. Soc. Parasitol. 1995, 25, 99–106. [Google Scholar]
- Nadim, A.; Faghih, M. The epidemiology of cutaneous leishmaniasis in the Isfahan province of Iran. I. The reservoir. II. The human disease. Trans. R. Soc. Trop. Med. Hyg. 1968, 62, 534–542. [Google Scholar] [CrossRef]
- Rioux, J.A.; Dereure, J.; Khiami, A.; Pratlong, F.; Sirdar, K.; Lambert, M. Ecoepidemiology of leishmaniasis in Syria. 1. Leishmania major Yakimoff and Schokhor (Kinetoplastida-Trypanosomatidae) infestation of Psammomys obesus Cretzschmar (Rodentia-Gerbillidae). Ann. Parasitol. Hum. Comp. 1990, 65, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Fichet-Calvet, E.; Jomaa, I.; Ben Ismail, R.; Ashford, R.W. Leishmania major infection in the fat sand rat Psammomys obesus in Tunisia: Interaction of host and parasite populations. Ann. Trop. Med. Parasitol. 2003, 97, 593–603. [Google Scholar] [CrossRef]
- Ghawar, W.; Toumi, A.; Snoussi, M.A.; Chlif, S.; Zaatour, A.; Boukthir, A.; Hamida, N.B.; Chemkhi, J.; Diouani, M.F.; Ben-Salah, A. Leishmania major infection among Psammomys obesus and Meriones shawi: Reservoirs of zoonotic cutaneous leishmaniasis in Sidi Bouzid (central Tunisia). Vector Borne Zoonotic Dis. 2011, 11, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Schilling, S.; Glaichenhaus, N. T cells that react to the immunodominant Leishmania major LACK antigen prevent early dissemination of the parasite in susceptible BALB/c mice. Infect. Immun. 2001, 69, 1212–1214. [Google Scholar] [CrossRef] [Green Version]
- Pourmohammadi, B.; Mohammadi-Azni, S.; Kalantari, M. Natural infection of Nesokia indica with Leishmania major and Leishmania infantum parasites in Damghan city, Northern Iran. Acta Trop. 2017, 170, 134–139. [Google Scholar] [CrossRef]
- Ajaoud, M.; Es-sette, N.; Hamdi, S.; El-Idrissi, A.L.; Riyad, M.; Lemrani, M. Detection and molecular typing of Leishmania tropica from Phlebotomus sergenti and lesions of cutaneous leishmaniasis in an emerging focus of Morocco. Parasit. Vectors 2013, 6, 217. [Google Scholar] [CrossRef] [Green Version]
- Chicharro, C.; Llanes-Acevedo, I.P.; Garcia, E.; Nieto, J.; Moreno, J.; Cruz, I. Molecular typing of Leishmania infantum isolates from a leishmaniasis outbreak in Madrid, Spain, 2009 to 2012. Eur. Surveill 2013, 18, 20545. [Google Scholar] [CrossRef] [Green Version]
- Ghatee, M.A.; Sharifi, I.; Kuhls, K.; Kanannejad, Z.; Harandi, M.F.; de Almeida, M.E.; Hatam, G.; Mirhendi, H. Heterogeneity of the internal transcribed spacer region in Leishmania tropica isolates from southern Iran. Exp. Parasitol. 2014, 144, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Hajjaran, H.; Mohebali, M.; Mamishi, S.; Vasigheh, F.; Oshaghi, M.A.; Naddaf, S.R.; Teimouri, A.; Edrissian, G.H.; Zarei, Z. Molecular identification and polymorphism determination of cutaneous and visceral leishmaniasis agents isolated from human and animal hosts in Iran. Biomed. Res. Int. 2013, 2013, 789326. [Google Scholar] [CrossRef] [Green Version]
- Jafari, R.; Najafzadeh, N.; Sedaghat, M.M.; Parvizi, P. Molecular characterization of sandflies and Leishmania detection in main vector of zoonotic cutaneous leishmaniasis in Abarkouh district of Yazd province, Iran. Asian Pac. J. Trop. Med. 2013, 6, 792–797. [Google Scholar] [CrossRef] [Green Version]
- Mahdy, M.A.K.; Al-Mekhlafi, H.M.; Al-Mekhlafi, A.M.; Lim, Y.A.L.; Bin Shuaib, N.O.M.; Azazy, A.A.; Mahmud, R. Molecular Characterization of Leishmania Species Isolated from Cutaneous Leishmaniasis in Yemen. PLoS ONE 2010, 5, e12879. [Google Scholar] [CrossRef]
- Salloum, T.; Khalifeh, I.; Tokajian, S. Detection, molecular typing and phylogenetic analysis of Leishmania isolated from cases of leishmaniasis among Syrian refugees in Lebanon. Parasite Epidemiol. Control 2016, 1, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Spotin, A.; Rouhani, S.; Parvizi, P. The associations of Leishmania major and Leishmania tropica aspects by focusing their morphological and molecular features on clinical appearances in Khuzestan Province, Iran. Biomed. Res. Int 2014, 2014, 913510. [Google Scholar] [CrossRef] [Green Version]
- Zahirnia, A.H.; Bordbar, A.; Ebrahimi, S.; Spotin, A.; Mohammadi, S.; Ghafari, S.M.; Ahmadvand, S.; Jabbari, N.; Esmaeili Rastaghi, A.R.; Parvizi, P. Predominance of Leishmania major and rare occurrence of Leishmania tropica with haplotype variability at the center of Iran. Braz. J. Infect. Dis. 2018, 22, 278–287. [Google Scholar] [CrossRef]
- Kariyawasam, U.L.; Selvapandiyan, A.; Rai, K.; Wani, T.H.; Ahuja, K.; Beg, M.A.; Premathilake, H.U.; Bhattarai, N.R.; Siriwardena, Y.D.; Zhong, D.B.; et al. Genetic diversity of Leishmania donovani that causes cutaneous leishmaniasis in Sri Lanka: A cross sectional study with regional comparisons. BMC Infect. Dis 2017, 17, 791. [Google Scholar] [CrossRef] [Green Version]
- Attia, H.; Sghaier, R.M.; Gelanew, T.; Bali, A.; Schweynoch, C.; Guerfali, F.Z.; Mkannez, G.; Chlif, S.; Belhaj-Hamida, N.; Dellagi, K.; et al. Genetic micro-heterogeneity of Leishmania major in emerging foci of zoonotic cutaneous leishmaniasis in Tunisia. Infect. Genet. Evol. 2016, 43, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Harrabi, M.; Bettaieb, J.; Ghawar, W.; Toumi, A.; Zaatour, A.; Yazidi, R.; Chaabane, S.; Chalghaf, B.; Hide, M.; Banuls, A.L.; et al. Spatio-temporal Genetic Structuring of Leishmania major in Tunisia by Microsatellite Analysis. PLoS Negl. Trop. Dis. 2015, 9, e0004017. [Google Scholar] [CrossRef] [PubMed]
- Chaouch, M.; Fathallah-Mili, A.; Driss, M.; Lahmadi, R.; Ayari, C.; Guizani, I.; Ben Said, M.; BenAbderrazak, S. Identification of Tunisian Leishmania spp. by PCR amplification of cysteine proteinase B (cpb) genes and phylogenetic analysis. Acta Trop. 2013, 125, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Chaara, D.; Ravel, C.; Banuls, A.; Haouas, N.; Lami, P.; Talignani, L.; El Baidouri, F.; Jaouadi, K.; Harrat, Z.; Dedet, J.P.; et al. Evolutionary history of Leishmania killicki (synonymous Leishmania tropica) and taxonomic implications. Parasit. Vectors 2015, 8, 198. [Google Scholar] [CrossRef] [Green Version]
- Shafiei, R.; Kalantari, M.; Yousefi, M.; Aspatwar, A.; Arzamani, K.; Bozorgomid, A.; Mirahmadi, H.; Soleimani, A.; Raeghi, S. Bionomics and phylo-molecular analysis of Leishmania species isolated from human lesions using ITS1 genes in northeast of Iran. J. Parasit. Dis. 2021, 45, 754–761. [Google Scholar] [CrossRef]
- El Hamouchi, A.; Ajaoud, M.; Arroub, H.; Charrel, R.; Lemrani, M. Genetic diversity of Leishmania tropica in Morocco: Does the dominance of one haplotype signify its fitness in both predominantly anthropophilic Phlebotomus sergenti and human beings? Transbound. Emerg. Dis. 2019, 66, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Ben Faleh, A.; Granjon, L.; Tatard, C.; Boratynski, Z.; Cosson, J.F.; Said, K. Phylogeography of two cryptic species of African desert jerboas (Dipodidae: Jaculus). Biol. J. Linn. Soc. 2012, 107, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Moutinho, A.F.; Seren, N.; Pauperio, J.; Silva, T.L.; Martinez-Freiria, F.; Sotelo, G.; Faria, R.; Mappes, T.; Alves, P.C.; Brito, J.C.; et al. Evolutionary history of two cryptic species of northern African jerboas. BMC Evol. Biol. 2020, 20, 26. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Jaculus (J.) jaculus n = 22 | J. hirtipes n = 24 | |
|---|---|---|
| Location of capture * n (%) | ||
| − Guermessa | 8 (36.0) | 8 (33.3) |
| − BniMhira | 14 (64.0) | 16 (66.7) |
| Gender * n (%) | ||
| − Males | 4 (18.2) | 6 (25.0) |
| − Females | 18 (81.8) | 18 (75.0) |
| Sets of Sequences | NS | H | S | η | k | Hd (SD) | π (SD) |
|---|---|---|---|---|---|---|---|
| Different hosts (Tunisia) | 73 | 9 | 40 | 44 | 12.44 | 0.707 (0.029) | 0.1575 (0.004) |
| Rodents hosts (all world) | 71 | 11 | 16 | 18 | 2.97 | 0.765 (0.031) | 0.0327 (0.003) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghawar, W.; Chaouch, M.; Ben Salah, A.; Snoussi, M.A.; Salem, S.; Kharroubi, G.; Chouchen, S.; Bouaoun, A.; Laouini, D.; Bettaieb, J.; et al. First Report of Two Jaculus Rodents as Potential Reservoir Hosts of Leishmania Parasites in Tunisia. Microorganisms 2022, 10, 1502. https://doi.org/10.3390/microorganisms10081502
Ghawar W, Chaouch M, Ben Salah A, Snoussi MA, Salem S, Kharroubi G, Chouchen S, Bouaoun A, Laouini D, Bettaieb J, et al. First Report of Two Jaculus Rodents as Potential Reservoir Hosts of Leishmania Parasites in Tunisia. Microorganisms. 2022; 10(8):1502. https://doi.org/10.3390/microorganisms10081502
Chicago/Turabian StyleGhawar, Wissem, Melek Chaouch, Afif Ben Salah, Mohamed Ali Snoussi, Sadok Salem, Ghassen Kharroubi, Said Chouchen, Amor Bouaoun, Dhafer Laouini, Jihene Bettaieb, and et al. 2022. "First Report of Two Jaculus Rodents as Potential Reservoir Hosts of Leishmania Parasites in Tunisia" Microorganisms 10, no. 8: 1502. https://doi.org/10.3390/microorganisms10081502