
Increased Prevalence of Salmonella Infantis Isolated from Raw Chicken and Turkey Products in the United States Is Due to a Single Clonal Lineage Carrying the pESI Plasmid



Abstract
:1. Introduction
2. Materials and Methods
3. Results
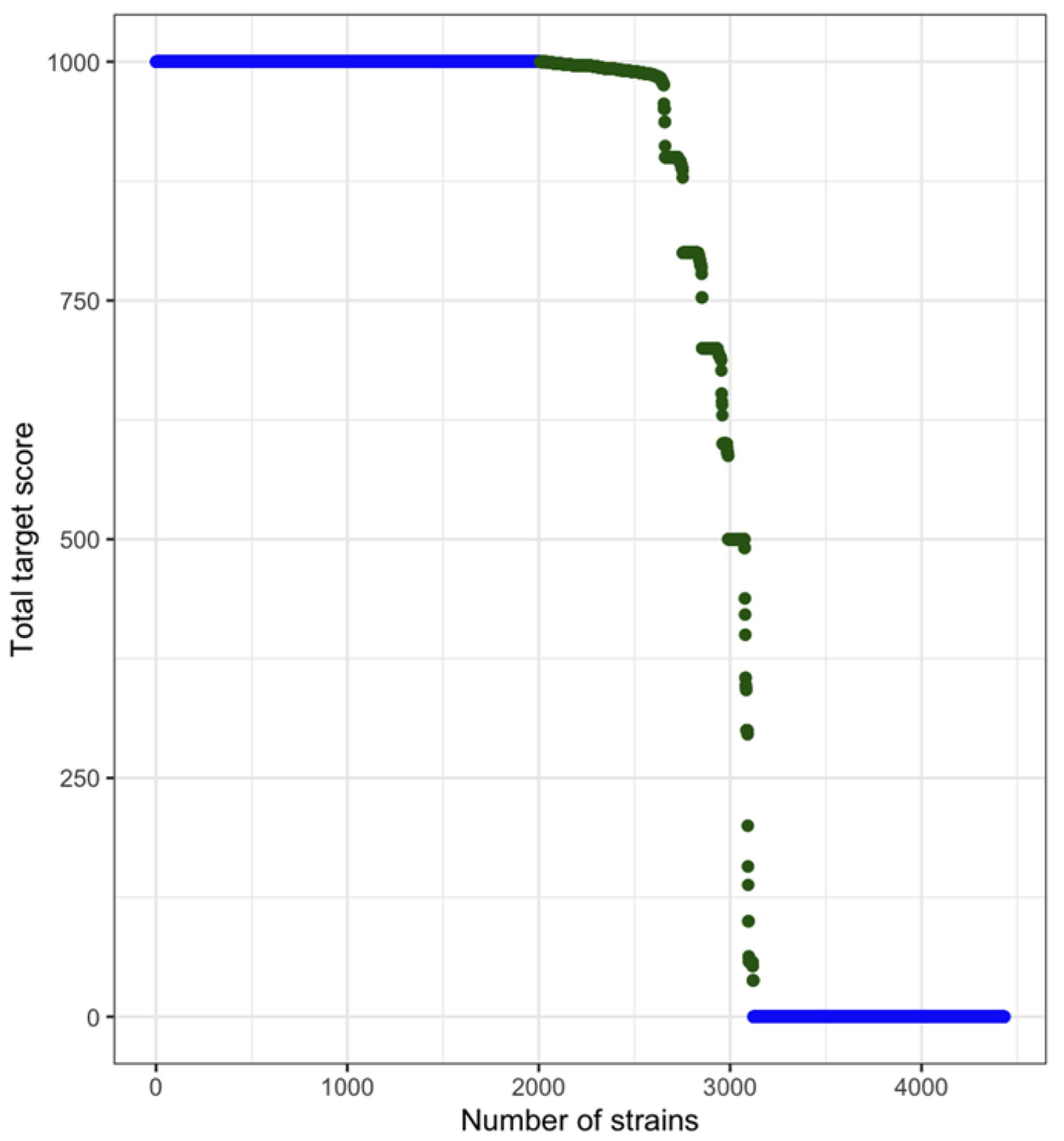
3.1. Curating of Publicly Available Strains from NCBI
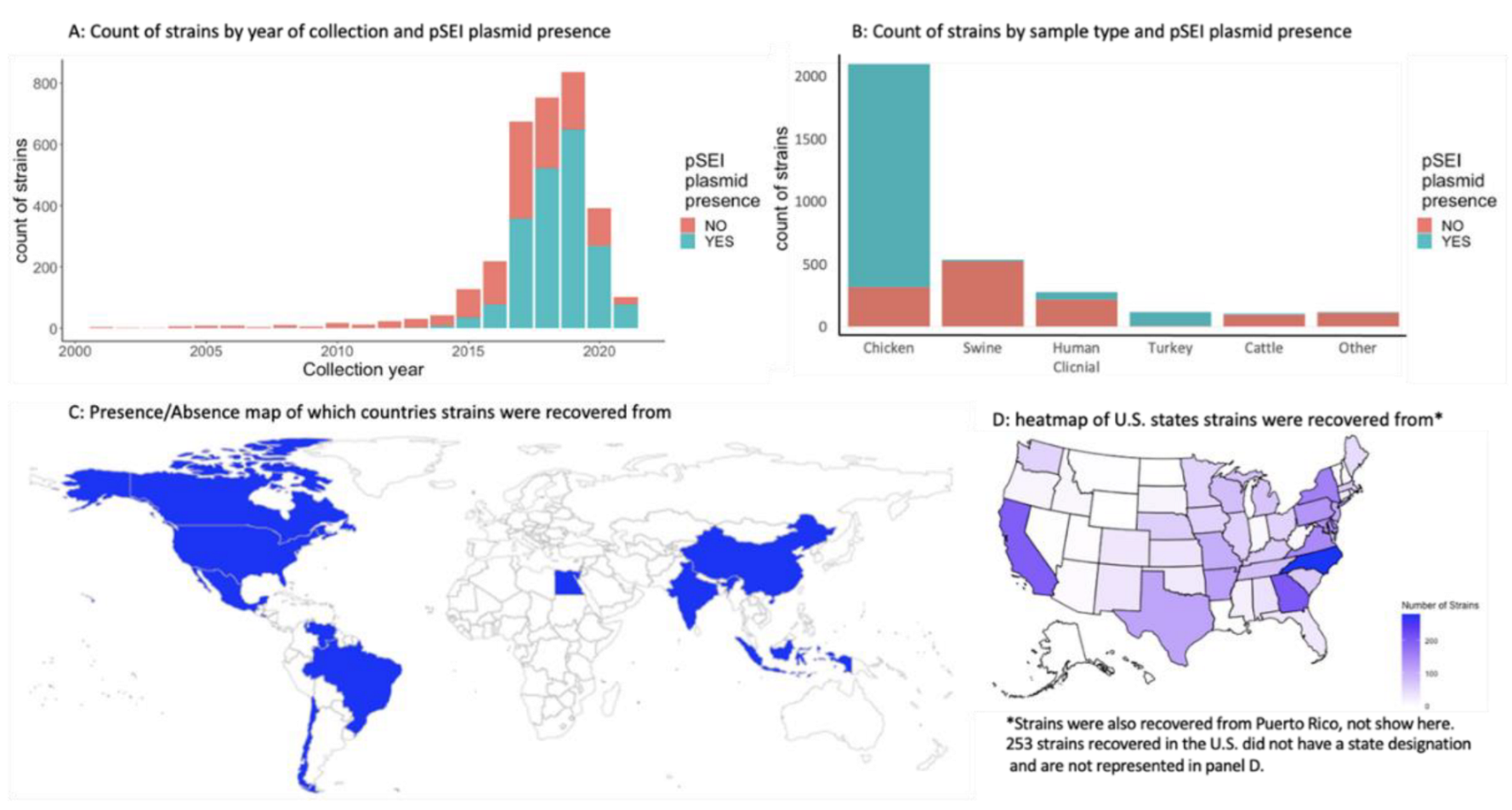
3.2. Metadata Summary
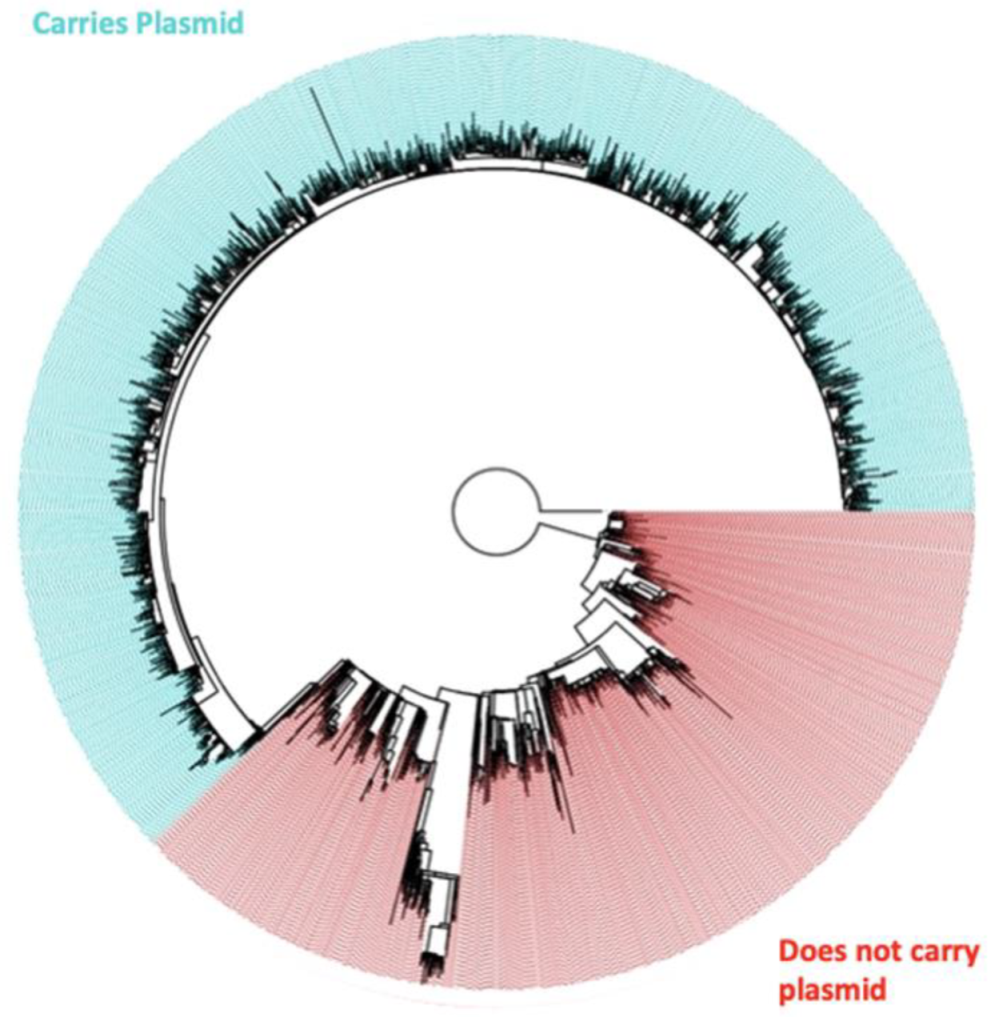
3.3. Description of the Population
3.4. SNP Differences
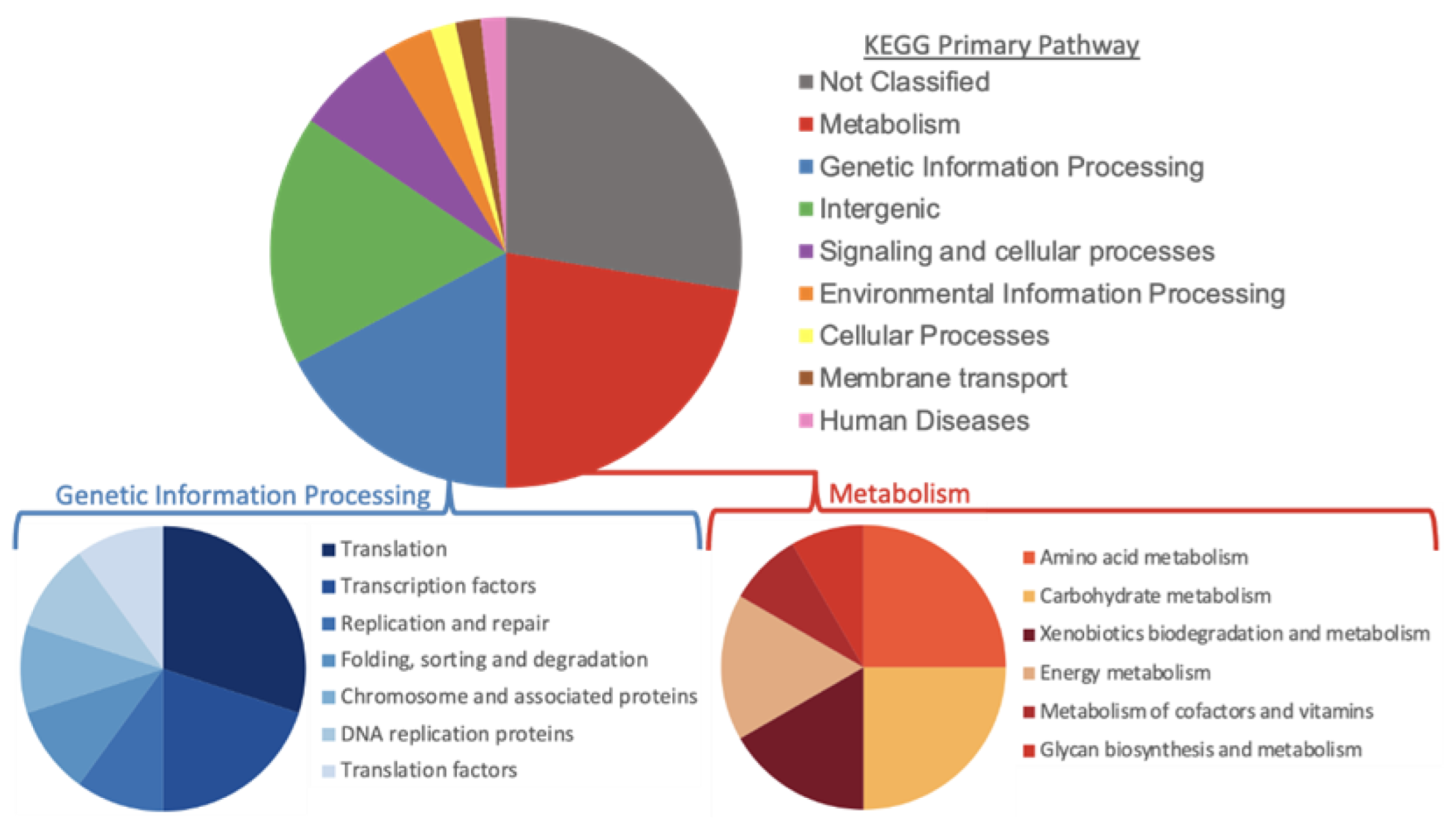
3.5. Pan-Genome Analysis
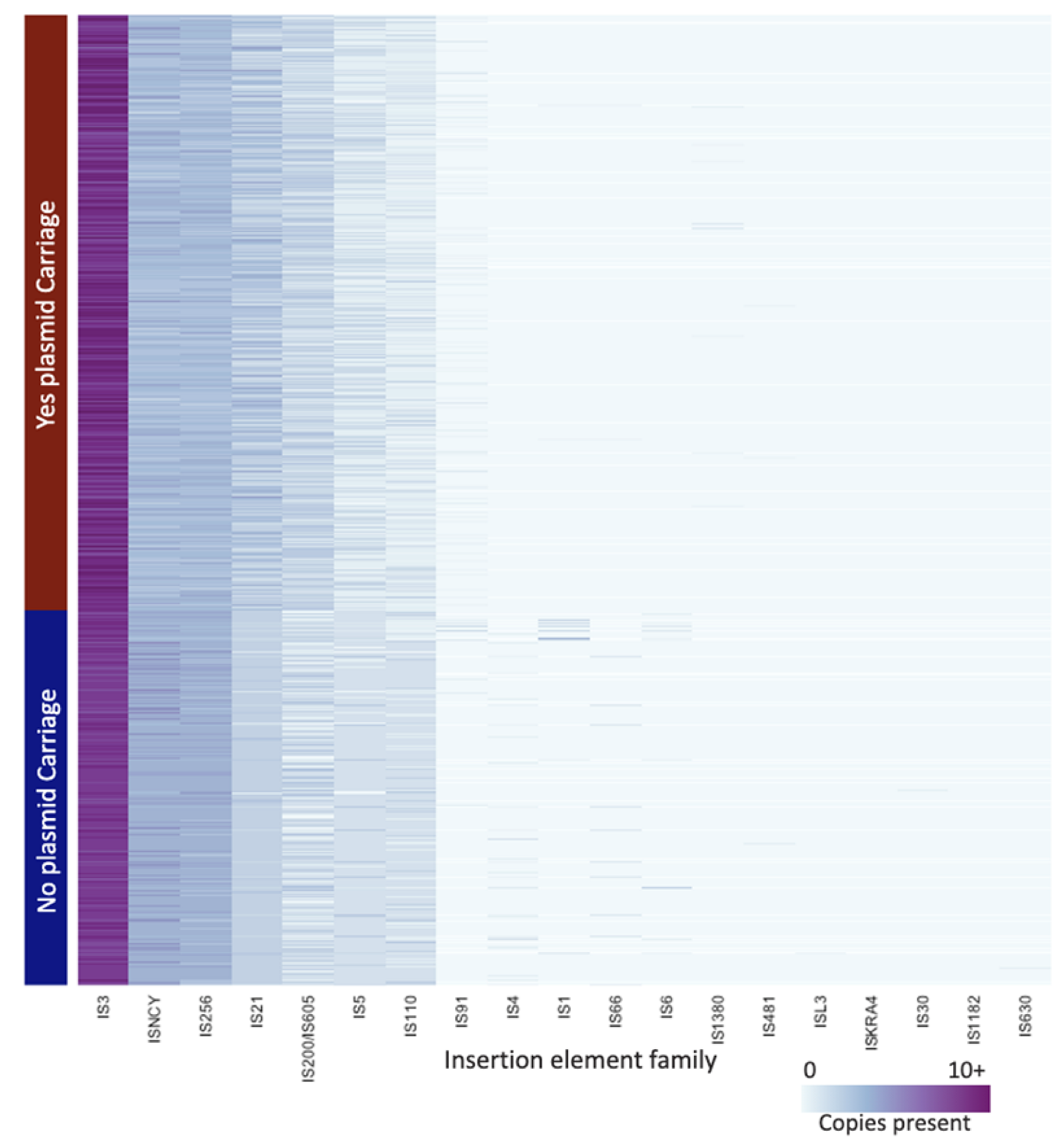
3.6. Insertion Element Family Differences
3.7. Describing the Intermediate Groups
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Tack, D.M.; Ray, L.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Rissman, T.; Jervis, R.; Lathrop, S.; Muse, A.; Duwell, M.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2016–2019. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 509–514. [Google Scholar] [CrossRef]
- CDC. An Atlas of Salmonella in the United States, 1968–2011; Laboratory-Based Disease Surveillance: Atlanta, GA, USA, 2013.
- FSIS Food Safety and Inspection Service. Salmonella Quarterly Reports: FY16. 2016. Available online: https://www.fsis.usda.gov/science-data/data-sets-visualizations/microbiology/microbiological-testing-program-rte-meat-and-7 (accessed on 3 May 2022).
- FSIS Food Safety and Inspection Service. Salmonella Quarterly Reports: FY21. 2021. Available online: https://www.fsis.usda.gov/science-data/data-sets-visualizations/microbiology/microbiological-testing-program-rte-meat-and-7 (accessed on 3 May 2022).
- Brown, A.C.; Chen, J.C.; Watkins, L.K.F.; Campbell, D.; Folster, J.P.; Tate, H.; Wasilenko, J.; Van Tubbergen, C.; Friedman, C.R. CTX-M-65 Extended-Spectrum β-Lactamase–Producing Salmonella enterica Serotype Infantis, United States(1). Emerg. Infect. Dis. 2018, 24, 2284–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, H.; Folster, J.P.; Hsu, C.-H.; Chen, J.; Hoffmann, M.; Li, C.; Morales, C.; Tyson, G.H.; Mukherjee, S.; Brown, A.C.; et al. Comparative Analysis of Extended-Spectrum-β-Lactamase CTX-M-65-Producing Salmonella enterica Serovar Infantis Isolates from Humans, Food Animals, and Retail Chickens in the United States. Antimicrob. Agents Chemother. 2017, 61, e00488-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aviv, G.; Rahav, G.; Gal-Mor, O. Horizontal Transfer of the Salmonella enterica Serovar Infantis Resistance and Virulence Plasmid pESI to the Gut Microbiota of Warm-Blooded Hosts. mBio 2016, 7, e01395-16. [Google Scholar] [CrossRef] [Green Version]
- Aviv, G.; Tsyba, K.; Steck, N.; Salmon-Divon, M.; Cornelius, A.; Rahav, G.; Grassl, A.G.; Gal-Mor, O. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ. Microbiol. 2014, 16, 977–994. [Google Scholar] [CrossRef]
- McMillan, E.A.; Wasilenko, J.L.; Tagg, K.A.; Chen, J.C.; Simmons, M.; Gupta, S.K.; Tillman, G.E.; Folster, J.; Jackson, C.R.; Frye, J.G. Carriage and Gene Content Variability of the pESI-Like Plasmid Associated with Salmonella Infantis Recently Established in United States Poultry Production. Genes 2020, 11, 1516. [Google Scholar] [CrossRef]
- Tyson, G.H.; Li, C.; Harrison, L.B.; Martin, G.; Hsu, C.-H.; Tate, H.; Tran, T.-T.; Strain, E.; Zhao, S. A Multidrug-Resistant Salmonella Infantis Clone is Spreading and Recombining in the United States. Microb. Drug Resist. 2021, 27, 792–799. [Google Scholar] [CrossRef]
- Gymoese, P.; Kiil, K.; Torpdahl, M.; Østerlund, M.T.; Sørensen, G.; Olsen, J.E.; Nielsen, E.M.; Litrup, E. WGS based study of the population structure of Salmonella enterica serovar Infantis. BMC Genom. 2019, 20, 870. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Brynildsrud, O.; Bohlin, J.; Scheffer, L.; Eldholm, V. Rapid scoring of genes in microbial pan-genome-wide association studies with Scoary. Genome Biol. 2016, 17, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Tang, H. ISEScan: Automated identification of insertion sequence elements in prokaryotic genomes. Bioinformatics 2017, 33, 3340–3347. [Google Scholar] [CrossRef] [PubMed]
- Aviv, G.; Elpers, L.; Mikhlin, S.; Cohen, H.; Zilber, S.V.; Grassl, G.A.; Rahav, G.; Hensel, M.; Gal-Mor, O. The plasmid-encoded Ipf and Klf fimbriae display different expression and varying roles in the virulence of Salmonella enterica serovar Infantis in mouse vs. avian hosts. PLoS Pathog. 2017, 13, e1006559. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella Pathogenicity and Host Adaptation in Chicken-Associated Serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, S.; Nagasawa, S.; Hayashi-Nishino, M.; Yamaguchi, A.; Nishino, K. AcrA dependency of the AcrD efflux pump in Salmonella enterica serovar Typhimurium. J. Antibiot. 2011, 64, 433–437. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Roof, D.M.; Roth, J.R. Ethanolamine utilization in Salmonella typhimurium. J. Bacteriol. 1988, 170, 3855–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeter, V.L.; Escalante-Semerena, J.C. Insights into the Relationship between Cobamide Synthase and the Cell Membrane. mBio 2021, 12, e00215-21. [Google Scholar] [CrossRef] [PubMed]
- Hryniewicz, M.M.; Kredich, N.M. The cysP promoter of Salmonella typhimurium: Characterization of two binding sites for CysB protein, studies of in vivo transcription initiation, and demonstration of the anti-inducer effects of thiosulfate. J. Bacteriol. 1991, 173, 5876–5886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, N.A.; Havemann, G.D.; Bobik, T.A. PduP is a coenzyme-a-acylating propionaldehyde dehydrogenase associated with the polyhedral bodies involved in B 12 -dependent 1,2-propanediol degradation by Salmonella enterica serovar Typhimurium LT2. Arch. Microbiol. 2003, 180, 353–361. [Google Scholar] [CrossRef]
- Zeiner, S.A.; Dwyer, B.E.; Clegg, S. FimA, FimF, and FimH Are Necessary for Assembly of Type 1 Fimbriae on Salmonella enterica Serovar Typhimurium. Infect. Immun. 2012, 80, 3289–3296. [Google Scholar] [CrossRef] [Green Version]
- Russell, P.W.; Orndorff, E.P. Lesions in two Escherichia coli type 1 pilus genes alter pilus number and length without affecting receptor binding. J. Bacteriol. 1992, 174, 5923–5935. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Phillippy, A.M.; Deng, K.; Rui, X.; Li, Z.; Tortorello, M.L.; Zhang, W. Transcriptomic Responses of Salmonella enterica Serovars Enteritidis and Typhimurium to Chlorine-Based Oxidative Stress. Appl. Environ. Microbiol. 2010, 76, 5013–5024. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Matsumoto, M.; Hata, M.; Takahashi, M.; Sakae, K. Development of a Rapid PCR Method Using the Insertion Sequence IS 1203 for Genotyping Shiga Toxin-Producing Escherichia coli O157. J. Clin. Microbiol. 2004, 42, 5462–5466. [Google Scholar] [CrossRef] [Green Version]
- van Embden, J.D.; Cave, M.D.; Crawford, J.T.; Dale, J.W.; Eisenach, K.D.; Gicquel, B.; Hermans, P.; Martin, C.; McAdam, R.; Shinnick, T.M. Strain identification of Mycobacterium tuberculosis by DNA fingerprinting: Recommendations for a standardized methodology. J. Clin. Microbiol. 1993, 31, 406–409. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Gourbeyre, E.; Chandler, M. Bacterial insertion sequences: Their genomic impact and diversity. FEMS Microbiol. Rev. 2014, 38, 865–891. [Google Scholar] [CrossRef] [Green Version]
- FDA. NARMS Retail Meat Report: 2014–2015. 2015. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/integrated-reportssummaries (accessed on 3 May 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome Position | Gene Function | Ref. Allele (pESI+) * | Alt. Allele (pESI-) | Strain Groups | |
|---|---|---|---|---|---|
| Intermediate groups that do not carry the pESI-like plasmid | GP-1 1 (n = 7) | ||||
| 944796 | Intergenic | A | C | A | |
| 953613 | AraC family transcriptional regulator | A | G | A | |
| 1146423 | Intergenic | A | C | A | |
| 1176706 | NAD(P)H nitroreductase | A | G | A | |
| 1512725 | Nitroreductase A | A | G | A | |
| 1626589 | Cell division protein ZapC | T | G | T | |
| 1771815 | NADH dehydrogenase | A | G | A | |
| 2361903 | Intergenic | C | T | C | |
| 2451383 | Carbon-nitrogen hydrolase | A | T | A | |
| 2485165 | Penicillin-binding protein 2 | A | G | A | |
| 2552207 | Intergenic | C | T | C | |
| 2577135 | cobS | A | T | A | |
| 3052001 | PTS sugar transporter | T | C | T | |
| 3373891 | Alanine-tRNA ligase | T | C | T | |
| 3557602 | Exodeoxyribonuclease V subunit gamma | G | T | G | |
| 3694291 | ATPase | T | C | T | |
| 3883819 | Intergenic | A | C | A | |
| 4385282 | Intergenic | A | C | A | |
| 4522651 | ATP-dependent protease | C | G | C | |
| Intermediate groups that do carry the pESI-like plasmid | GP + 1 2 (n = 2) | GP + 2 3 (n = 2) | |||
| 1774378 | Peptidoglycan-binding protein LysM | G | A | G | A |
| 2836180 | DNA gyrase subunit A | A | C | C | A |
| 4137324 | Intergenic | A | T | T | T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMillan, E.A.; Weinroth, M.D.; Frye, J.G. Increased Prevalence of Salmonella Infantis Isolated from Raw Chicken and Turkey Products in the United States Is Due to a Single Clonal Lineage Carrying the pESI Plasmid. Microorganisms 2022, 10, 1478. https://doi.org/10.3390/microorganisms10071478
McMillan EA, Weinroth MD, Frye JG. Increased Prevalence of Salmonella Infantis Isolated from Raw Chicken and Turkey Products in the United States Is Due to a Single Clonal Lineage Carrying the pESI Plasmid. Microorganisms. 2022; 10(7):1478. https://doi.org/10.3390/microorganisms10071478
Chicago/Turabian StyleMcMillan, Elizabeth A., Margaret D. Weinroth, and Jonathan G. Frye. 2022. "Increased Prevalence of Salmonella Infantis Isolated from Raw Chicken and Turkey Products in the United States Is Due to a Single Clonal Lineage Carrying the pESI Plasmid" Microorganisms 10, no. 7: 1478. https://doi.org/10.3390/microorganisms10071478