
Role of Dissimilative Pathway of Komagataella phaffii (Pichia pastoris): Formaldehyde Toxicity and Energy Metabolism

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction

2. Materials and Methods
2.1. Strains
2.2. Cell Cultures and Formaldehyde Pulses
2.3. Parameter Calculation
2.4. Metabolic Flux Analysis
2.5. Analytical Methods
2.6. Satatistical Analysis
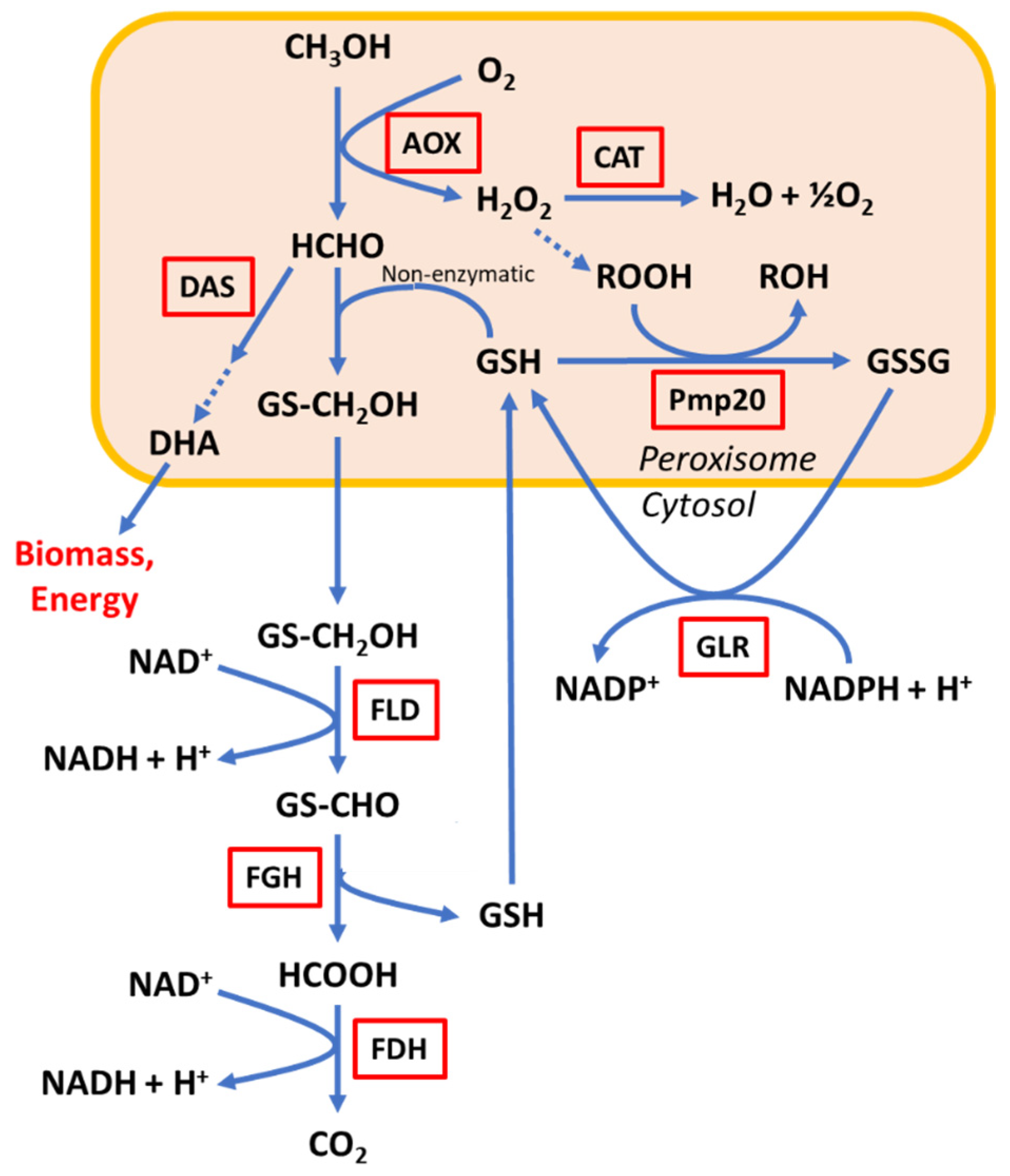
3. Results and Discussion
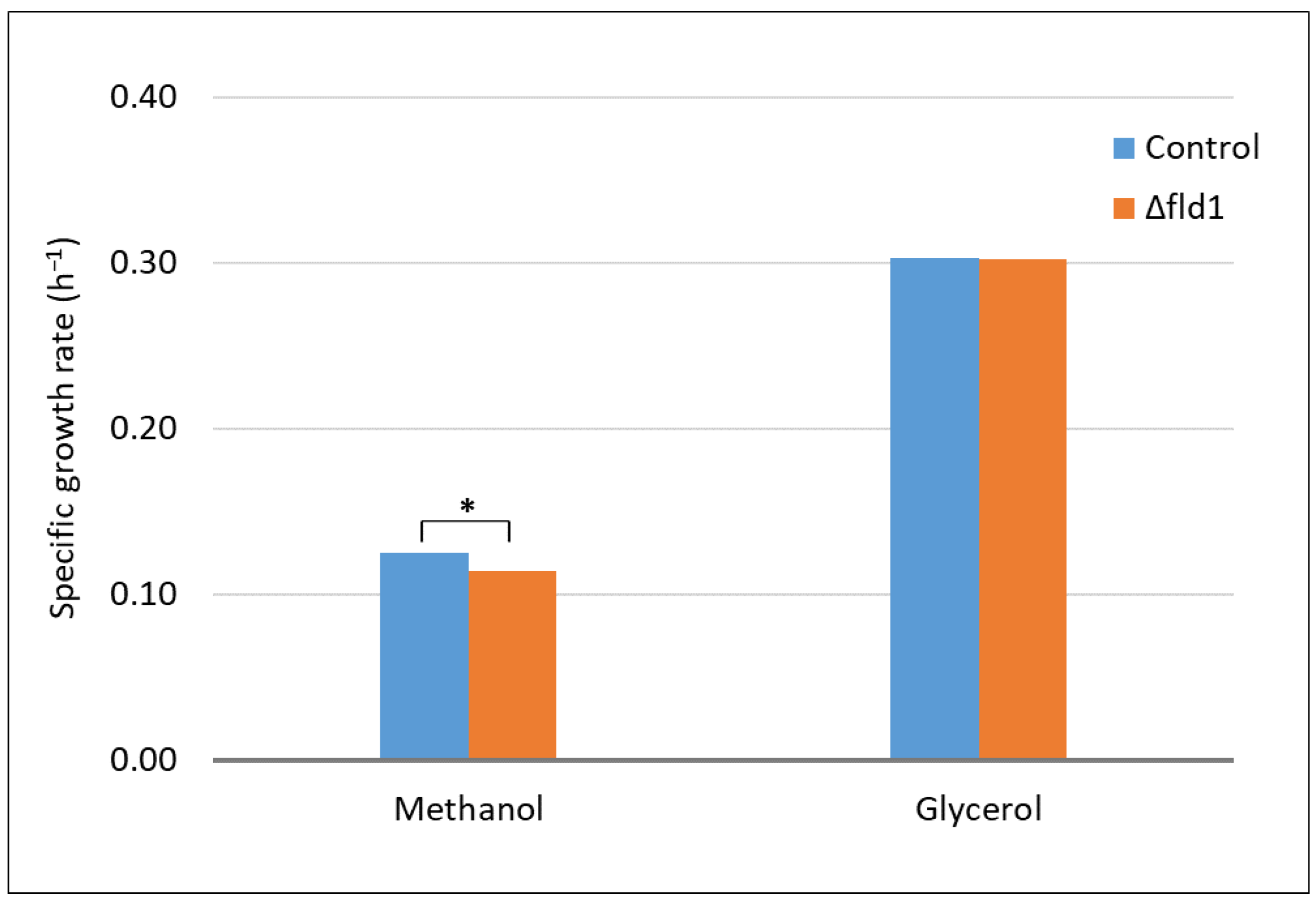
3.1. Effect of FLD1 Gene Deletion on Cell Growth
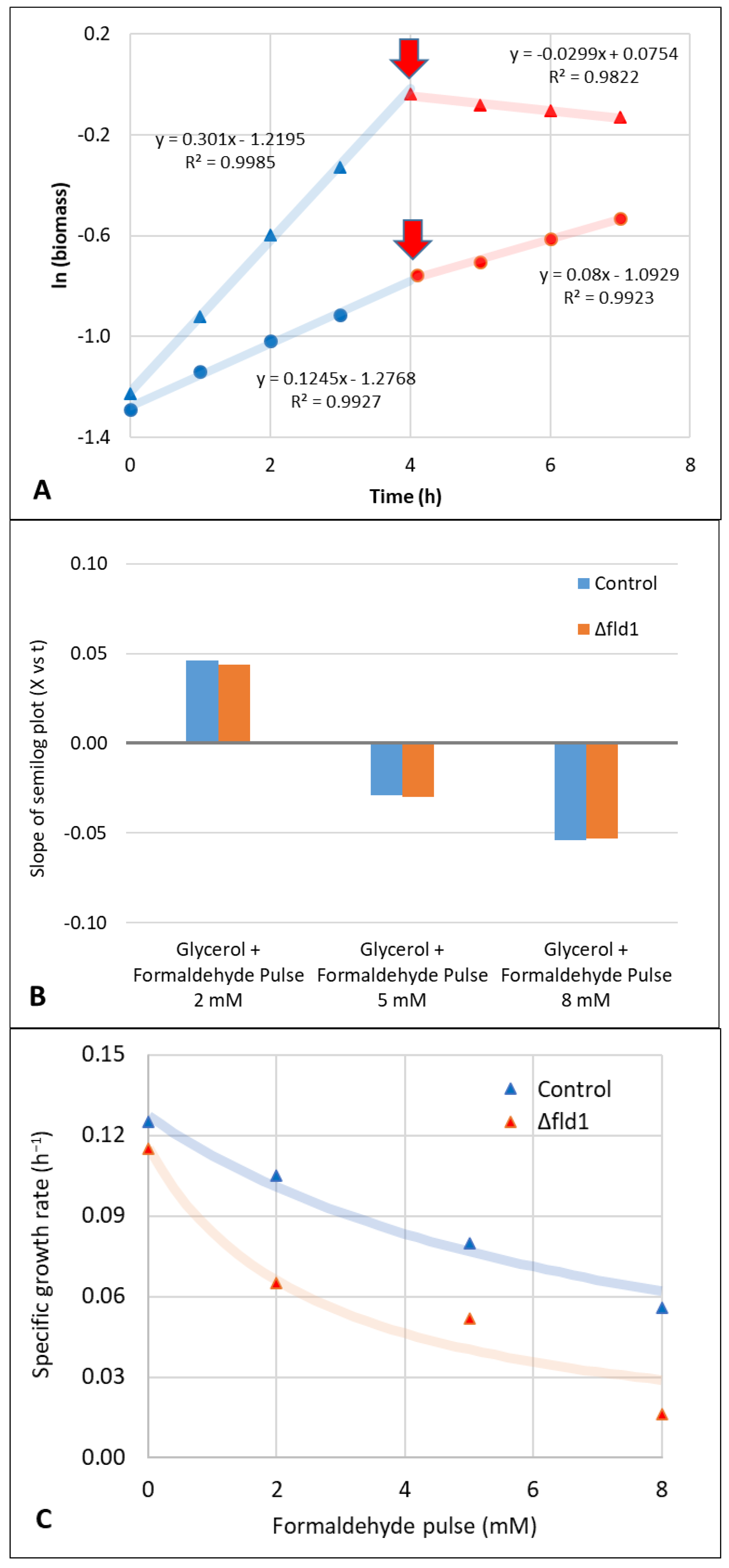
3.2. Evaluation of Formaldehyde Toxicity
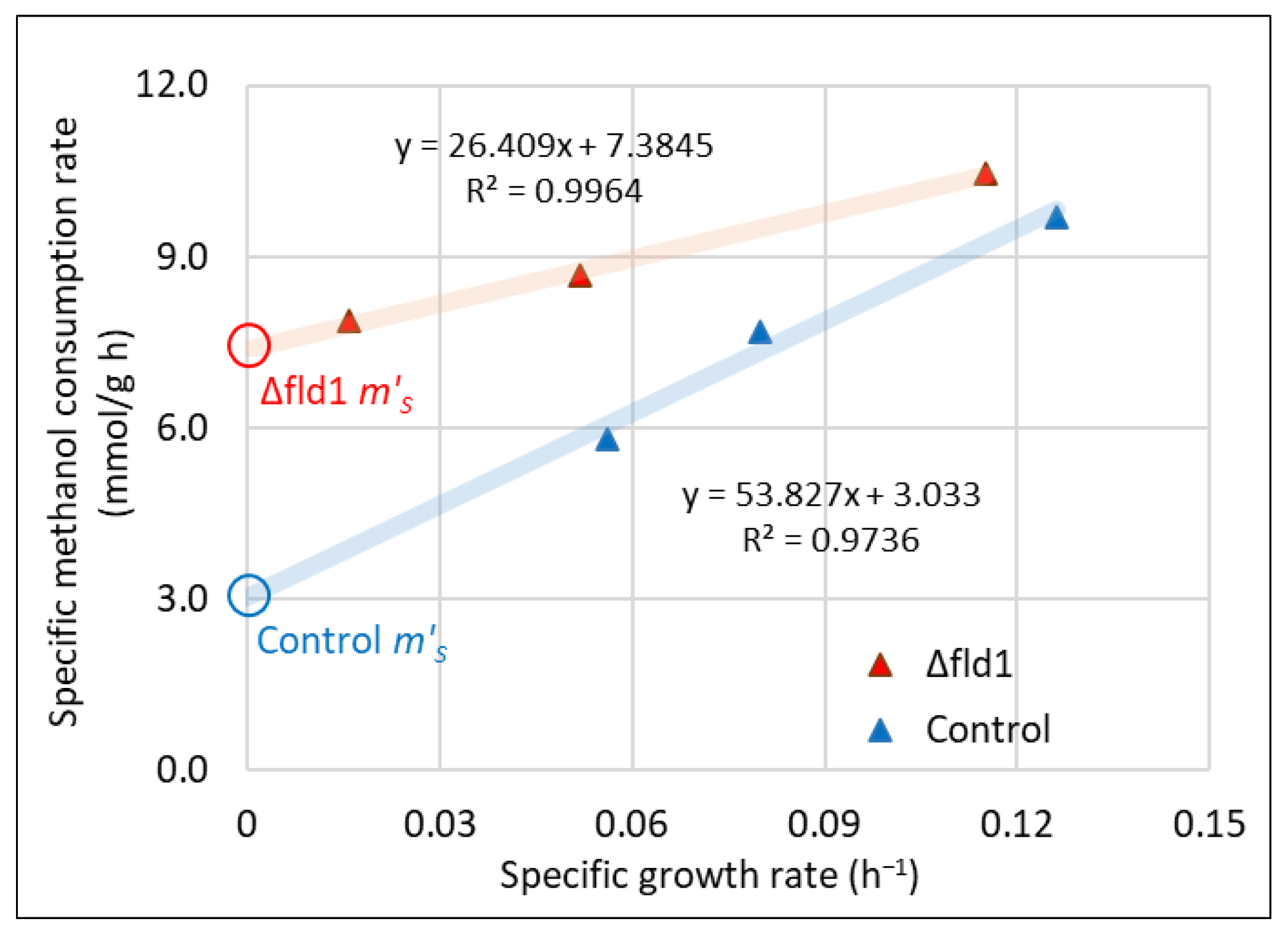
3.3. Maintenance Energy Estimation
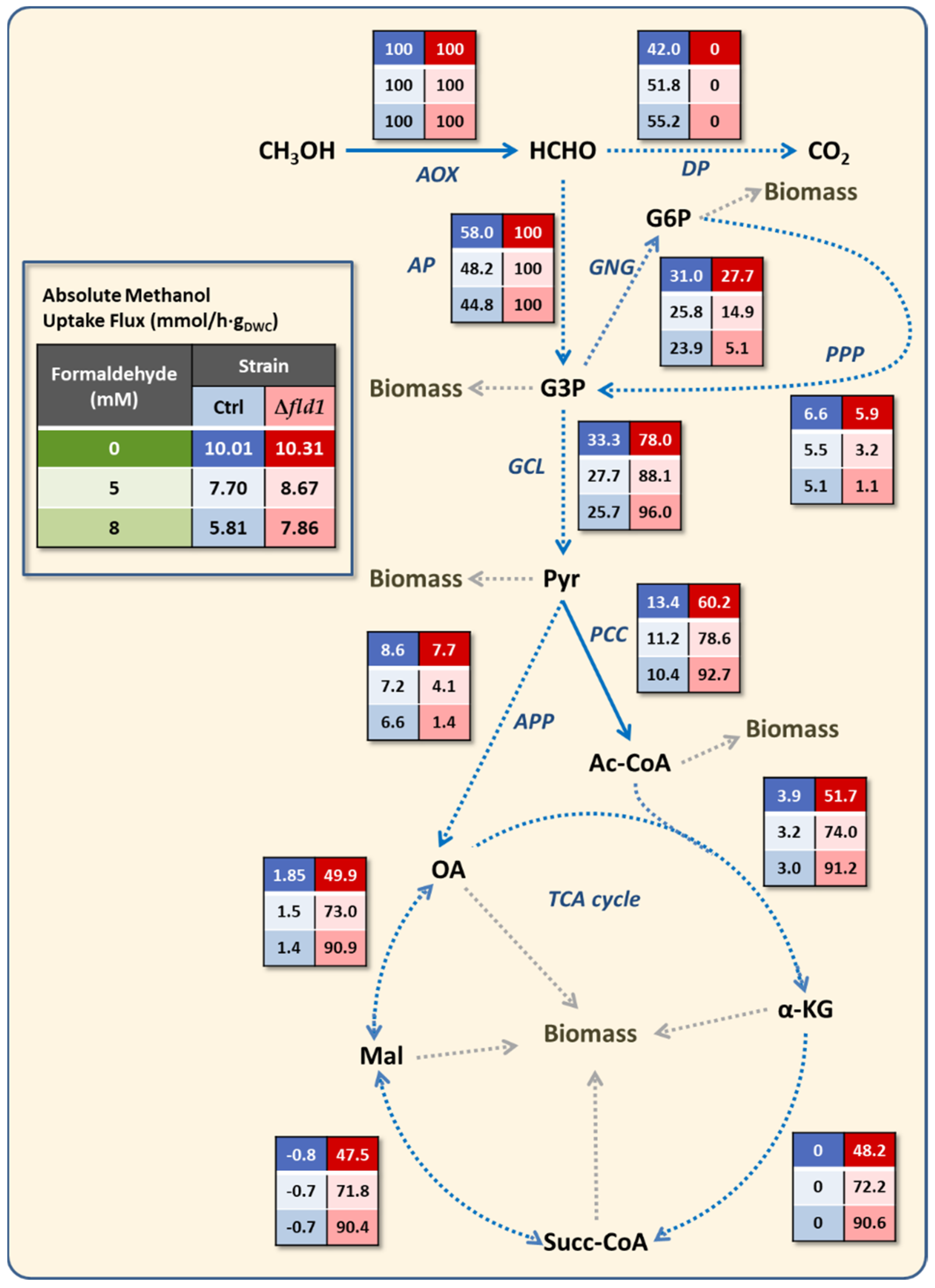
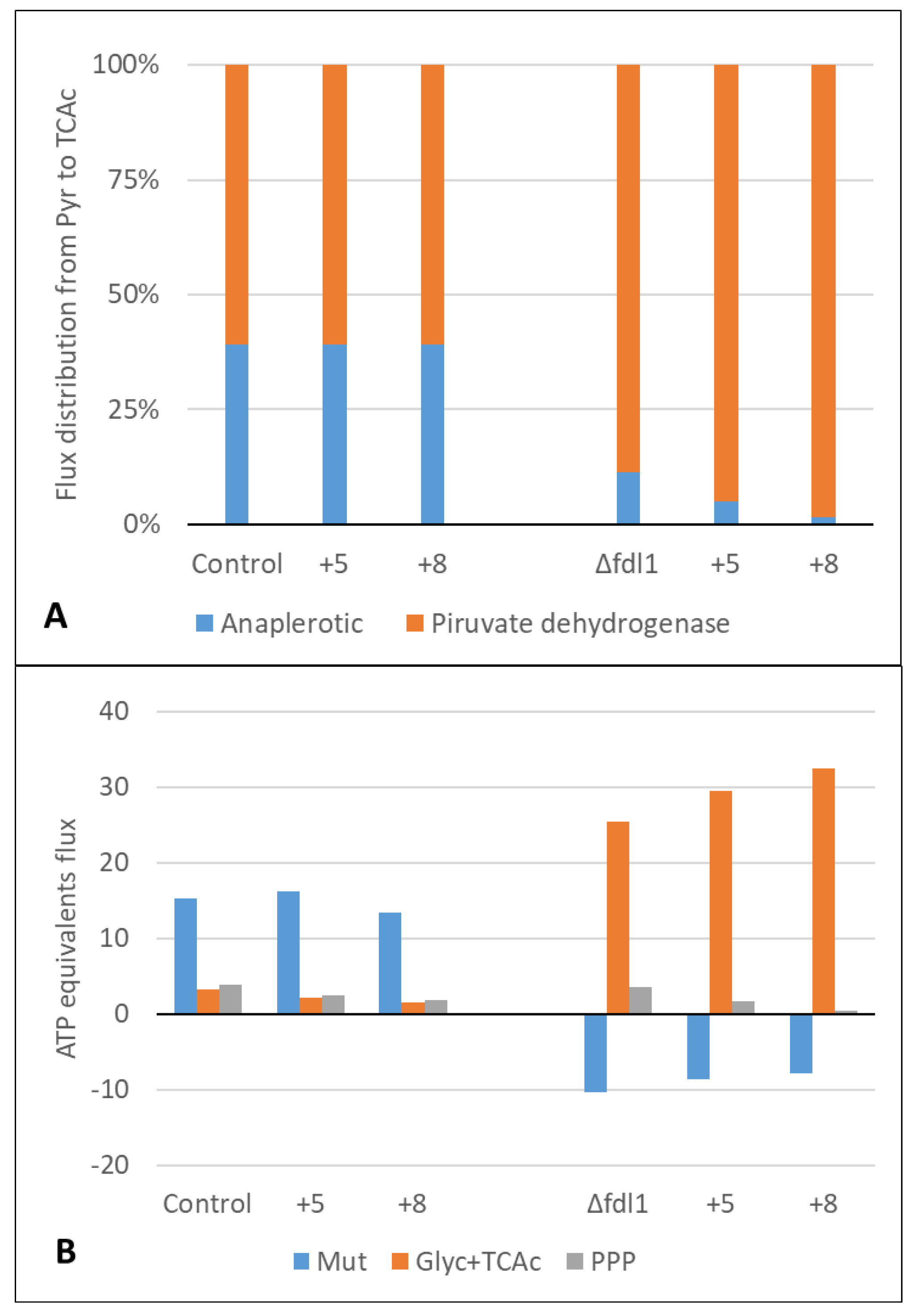
3.4. Metabolic Flux Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bustos, C.; Quezada, J.; Veas, R.; Altamirano, C.; Braun-Galleani, S.; Fickers, P.; Berrios, J. Advances in Cell Engineering of the Komagataella phaffii Platform for Recombinant Protein Production. Metabolites 2022, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.M.V.; Carmo, T.S.; Carvalho, L.S.; Bahia, F.M.; Parachin, N.S. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, S.; Oku, M.; Yurimoto, H.; Sakai, Y. Regulation of Peroxisome Homeostasis by Post-Translational Modification in the Methylotrophic Yeast Komagataella phaffii. Front. Cell Dev. Biol. 2022, 10, 887806. [Google Scholar] [CrossRef] [PubMed]
- North, M.; Gaytán, B.D.; Romero, C., Jr.; de la Rosa, V.Y.; Loguinov, A.; Smith, M.T.; Zhang, L.; Vulpe, C.D. Functional Toxicogenomic Profiling Expands Insight into Modulators of Formaldehyde Toxicity in Yeast. Front. Genet. 2016, 7, 200. [Google Scholar] [CrossRef] [Green Version]
- Vanz, A.; Lünsdorf, H.; Adnan, A.; Nimtz, M.; Gurramkonda, C.; Khanna, N.; Rinas, U. Physiological response of Pichia pastoris GS115 to methanol-induced high level production of the Hepatitis B surface antigen: Catabolic adaptation, stress responses, and autophagic processes. Microb. Cell Factories 2012, 11, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungo, C.; Marison, I.; von Stockar, U. Regulation of alcohol oxidase of a recombinant Pichia pastoris Mut+ strain in transient continuous cultures. J. Biotechnol. 2007, 130, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, K.; Yamaguchi, S.; Takeuchi, A.; Mizumura, T.; Ozawa, S.; Tomizuka, N.; Hayakawa, T.; Nakagawa, T. Regulation of intracellular formaldehyde toxicity during methanol metabolism of the methylotrophic yeast Pichia methanolica. J. Biosci. Bioeng. 2016, 122, 545–549. [Google Scholar] [CrossRef]
- Rußmayer, H.; Buchetics, M.; Gruber, C.; Valli, M.; Grillitsch, K.; Modarres, G.; Guerrasio, R.; Klavins, K.; Neubauer, S.; Drexler, H.; et al. Systems-level organization of yeast methylotrophic lifestyle. BMC Biol. 2015, 13, 80. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Yurimoto, H.; Sakai, Y.; Kato, N. Physiological role of the glutathione-dependent formaldehyde dehydrogenase in the methylotrophic yeast Candida boidinii. Microbiology 2002, 148, 2697–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, T.; Takigami, E.; Yurimoto, H.; Sakai, Y. Yap1-Regulated Glutathione Redox System Curtails Accumulation of Formaldehyde and Reactive Oxygen Species in Methanol Metabolism of Pichia pastoris. Eukaryot Cell 2009, 8, 540–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fickers, P.; le Dall, M.T.; Gaillardin, C.; Thonart, P.; Nicaud, J.M. New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J. Microbiol. Methods 2003, 55, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Lin-Cereghino, J.; Wong, W.W.; Xiong, S.; Giang, W.; Luong, L.T.; Vu, J.; Johnson, S.D.; Lin-Cereghino, G.P. Condensed protocol for competent cell preparation and transformation of the methylotrophic yeast Pichia pastoris. Biotechniques 2005, 38, 44–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theron, C.W.; Berrios, J.; Steels, S.; Telek, S.; Lecler, R.; Rodriguez, C.; Fickers, P. Expression of recombinant enhanced green fluorescent protein provides insight into foreign gene-expression differences between Mut+ and MutS strains of Pichia pastoris. Yeast 2019, 36, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Looser, V.; Bruhlmann, B.; Bumbak, F.; Stenger, C.; Costa, M.; Camattari, A.; Fotiadis, D.; Kovar, K. Cultivation strategies to enhance productivity of Pichia pastoris: A review. Biotechnol. Adv. 2015, 33, 1177–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canales, C.; Altamirano, C.; Berrios, J. The growth of Pichia pastoris Mut+ on methanol–glycerol mixtures fits to interactive dual-limited kinetics: Model development and application to optimised fed-batch operation for heterologous protein production. Bioprocess Biosyst. Eng. 2018, 41, 1827–1838. [Google Scholar] [CrossRef]
- van Bodegom, P. Microbial Maintenance: A Critical Review on Its Quantification. Microb. Ecol. 2007, 53, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Pirt, S.J. Maintenance energy: A general model for energy-limited and energy-sufficient growth. Arch. Microbiol. 1982, 133, 300–302. [Google Scholar] [CrossRef]
- Velastegui, E.; Theron, C.; Berrios, J.; Fickers, P. Downregulation by organic nitrogen of AOX1 promoter used for controlled expression of foreign genes in the yeast Pichia pastoris. Yeast 2019, 36, 297–304. [Google Scholar] [CrossRef]
- Solà, A.; Jouhten, P.; Maaheimo, H.; Sánchez-Ferrando, F.; Szyperski, T.; Ferrer, P. Metabolic flux profiling of Pichia pastoris grown on glycerol/methanol mixtures in chemostat cultures at low and high dilution rates. Microbiology 2007, 153 Pt 1, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Çelik, E.; Çalık, P.; Oliver, S.G. Metabolic flux analysis for recombinant protein production by Pichia pastoris using dual carbon sources: Effects of methanol feeding rate. Biotechnol. Bioeng. 2010, 105, 317–329. [Google Scholar] [CrossRef]
- Unrean, P. Pathway analysis of Pichia pastoris to elucidate methanol metabolism and its regulation for production of recombinant proteins. Biotechnol. Prog. 2014, 30, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Zepeda, A.B.; Figueroa, C.A.; Abdalla, D.S.; Maranhão, A.Q.; Ulloa, P.H.; Pessoa, A., Jr.; Farías, J.G. HSF-1, HIF-1and HSP90 expression on recombinant Pichia pastoris under fed-batch fermentation. Braz. J. Microbiol. 2014, 45, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordà, J.; de Jesus, S.S.; Peltier, S.; Ferrer, P.; Albiol, J. Metabolic flux analysis of recombinant Pichia pastoris growing on different glycerol/methanol mixtures by iterative fitting of NMR-derived (13)C-labelling data from proteinogenic amino acids. New Biotechnol. 2014, 31, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, M.; Koh, G.; Chung, B.K.S.; Lee, D.-Y. Software applications for flux balance analysis. Brief. Bioinform. 2014, 15, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillaparassamee, O.; Chinwetkitvanich, S.; Kanchanasuta, S.; Pisutpaisal, N.; Champreda, V. Metabolic flux analysis on succinic acid production from crude glycerol by Actinobacillus succinogenes. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, D.Y.; Hong, S.H.; Kim, T.Y.; Yun, H.; Oh, Y.G.; Park, S. MetaFluxNet, a program package for metabolic pathway construction and analysis, and its use in large-scale metabolic flux analysis of Escherichia coli. Genome Inform. 2003, 14, 23–33. [Google Scholar]
- Sánchez, C.E.G.; Sáez, R.G.T. Comparison and analysis of objective functions in flux balance analysis. Biotechnol. Prog. 2014, 30, 985–991. [Google Scholar] [CrossRef]
- Bideaux, C.; Montheard, J.; Cameleyre, X.; Molina-Jouve, C.; Alfenore, S. Metabolic flux analysis model for optimizing xylose conversion into ethanol by the natural C5-fermenting yeast Candida shehatae. Appl. Microbiol. Biotechnol. 2016, 100, 1489–1499. [Google Scholar] [CrossRef]
- Sibirny, A.A.; Ubiyvovk, V.M.; Gonchar, M.V.; Titorenko, V.I.; Voronovsky, A.Y.; Kapultsevich, Y.G.; Bliznik, K.M. Reactions of direct formaldehyde oxidation to CO2 are non-essential for energy supply of yeast methylotrophic growth. Arch. Microbiol. 1990, 154, 566–575. [Google Scholar] [CrossRef]
- Tyurin, O.V.; Kozlov, D.G. Deletion of the FLD gene in methylotrophic yeasts Komagataella phaffii and Komagataella kurtzmanii results in enhanced induction of the AOX1 promoter in response to either methanol or formate. Microbiology 2015, 84, 408–411. [Google Scholar] [CrossRef]
- Shen, S.; Sulter, G.; Jeffries, T.W.; Cregg, J.M. A strong nitrogen source-regulated promoter for controlled expression of foreign genes in the yeast Pichia pastoris. Gene 1998, 216, 93–102. [Google Scholar] [CrossRef]
- Yurimoto, H.; Lee, B.; Yasuda, F.; Sakai, Y.; Kato, N. Alcohol dehydrogenases that catalyse methyl formate synthesis participate in formaldehyde detoxification in the methylotrophic yeast Candida boidinii. Yeast 2004, 21, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, Y.; Murdanoto, A.P.; Sembiring, L.; Tani, Y.; Kato, N. A novel formaldehyde oxidation pathway in methylotrophic yeasts: Methylformate as a possible intermediate. FEMS Microbiol. Lett. 1995, 127, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Tomàs-Gamisans, M.; Ferrer, P.; Albiol, J. Fine-tuning the P. pastoris iMT1026 genome-scale metabolic model for improved prediction of growth on methanol or glycerol as sole carbon sources. Microb. Biotechnol. 2018, 11, 224–237. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.; Huang, M.; Lu, H.; Qian, J.; Lin, W.; Chu, J.; Zhuang, Y.; Zhang, S. Comprehensive reconstruction and evaluation of Pichia pastoris genome-scale metabolic model that accounts for 1243 ORFs. Bioresour. Bioprocess. 2017, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.J.; Hinner, M.J. Getting Across the Cell Membrane: An Overview for Small Molecules, Peptides, and Proteins. Methods Mol. Biol. 2015, 1266, 29–53. [Google Scholar] [CrossRef] [Green Version]
- Barrigon, J.M.; Valero, F.; Montesinos, J.L. A macrokinetic model-based comparative meta-analysis of recombinant protein production by Pichia pastoris under AOX1 promoter. Biotechnol. Bioeng. 2015, 112, 1132–1145. [Google Scholar] [CrossRef]
- Canales, C.; Altamirano, C.; Berrios, J. Effect of dilution rate and methanol-glycerol mixed feeding on heterologous Rhizopus oryzae lipase production with Pichia pastoris Mut+ phenotype in continuous culture. Biotechnol. Prog. 2015, 31, 707–714. [Google Scholar] [CrossRef]
- Fukuoka, H.; Kawase, T.; Oku, M.; Yurimoto, H.; Sakai, Y.; Hayakawa, T.; Nakagawa, T. Peroxisomal Fba2p and Tal2p complementally function in the rearrangement pathway for xylulose 5-phosphate in the methylotrophic yeast Pichia pastoris. J. Biosci. Bioeng. 2019, 128, 33–38. [Google Scholar] [CrossRef]
- Cai, H.-L.; Doi, R.; Shimada, M.; Hayakawa, T.; Nakagawa, T. Metabolic regulation adapting to high methanol environment in the methylotrophic yeast Ogataea methanolica. Microb. Biotechnol. 2021, 14, 1512–1524. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Genotype-Plasmid | Source/Reference |
|---|---|---|
| E. coli DH5α | Δ(lacZYA-argF)U169 recA1 endA1 hsdR17(rK–, mK+) phoA supE44 λ– thi-1 gyrA96 relA1 F– φ80lacZΔM15 | Promega |
| RIE280 | DH5α, vector RIP280 (I-SceI -ZeoR- I-SceI) | Lab stock |
| RIE293 | DH5α, vector RIP293 (PFLD1- I-SceI -TFLD in pJet2.1) | This work |
| RIE293_ZeoR | DH5α, vector RIp293_ZeoR (PFLD1-ZeoR-TFLD) | This work |
| RIY232 | Komagatella phaffii GS115, HIS4 (protothroph) | Lab stock |
| RIY428 | RIY232, FLD1::ZeoR | This work |
| Primer | Primer sequences 5′-3′ | Modification |
| FLD1_PF | TACACAACGGATGTCGCACT | |
| FLD1_PR | CAGGAAACAGCTATGACCCGAACACAACAGGGAAACT | I-SceI |
| FLD1_TF | GTAAAACGACGGCCAGTTGGCAGAGTCTGGAGAGGAT | I-SceI |
| FLD1_TR | GAGATCCCAGGCATTCAGAG | |
| FLD1_verif | GGCACGGTGCTAATGGTAGT |
| Formaldehyde Pulse (mM) | ||||||
|---|---|---|---|---|---|---|
| 0 | 5 | 8 | ||||
| Strain | YXS | qs | YXS | qs | YXS | qs |
| Control | 0.38 ** | 10.01 ** | 0.32 | 7.70 | 0.31 | 5.81 |
| Δfld1 | 0.35 | 10.31 | 0.19 | 8.67 | 0.06 | 7.86 |
| Control | Δfld1 | |||||
|---|---|---|---|---|---|---|
| Formaldehyde Pulse (mM) | 0 * | 5 | 8 | 0 | 5 | 8 |
| Dissimilative pathway | 4.21 | 3.98 | 3.21 | - | - | - |
| Assimilative pathway | 5.80 | 3.72 | 2.60 | 10.31 | 8.67 | 7.86 |
| ATP flux for maintenance | 18.5 | 18.7 | 14.9 | 21.0 | 28.1 | 32.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berrios, J.; Theron, C.W.; Steels, S.; Ponce, B.; Velastegui, E.; Bustos, C.; Altamirano, C.; Fickers, P. Role of Dissimilative Pathway of Komagataella phaffii (Pichia pastoris): Formaldehyde Toxicity and Energy Metabolism. Microorganisms 2022, 10, 1466. https://doi.org/10.3390/microorganisms10071466
Berrios J, Theron CW, Steels S, Ponce B, Velastegui E, Bustos C, Altamirano C, Fickers P. Role of Dissimilative Pathway of Komagataella phaffii (Pichia pastoris): Formaldehyde Toxicity and Energy Metabolism. Microorganisms. 2022; 10(7):1466. https://doi.org/10.3390/microorganisms10071466
Chicago/Turabian StyleBerrios, Julio, Chrispian W. Theron, Sébastien Steels, Belén Ponce, Edgar Velastegui, Cristina Bustos, Claudia Altamirano, and Patrick Fickers. 2022. "Role of Dissimilative Pathway of Komagataella phaffii (Pichia pastoris): Formaldehyde Toxicity and Energy Metabolism" Microorganisms 10, no. 7: 1466. https://doi.org/10.3390/microorganisms10071466