
Yeast Diversity during Spontaneous Fermentations and Oenological Characterisation of Indigenous Saccharomyces cerevisiae for Potential as Wine Starter Cultures

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Yeasts Enumeration, Isolation and Molecular Identification
2.3. Strain Typing of S. cerevisiae Isolates
2.4. Fermentation Performance of Selected S. cerevisiae Strains
2.5. Profiling of Wine Composition
2.6. Statistical Analysis
3. Results and Discussion
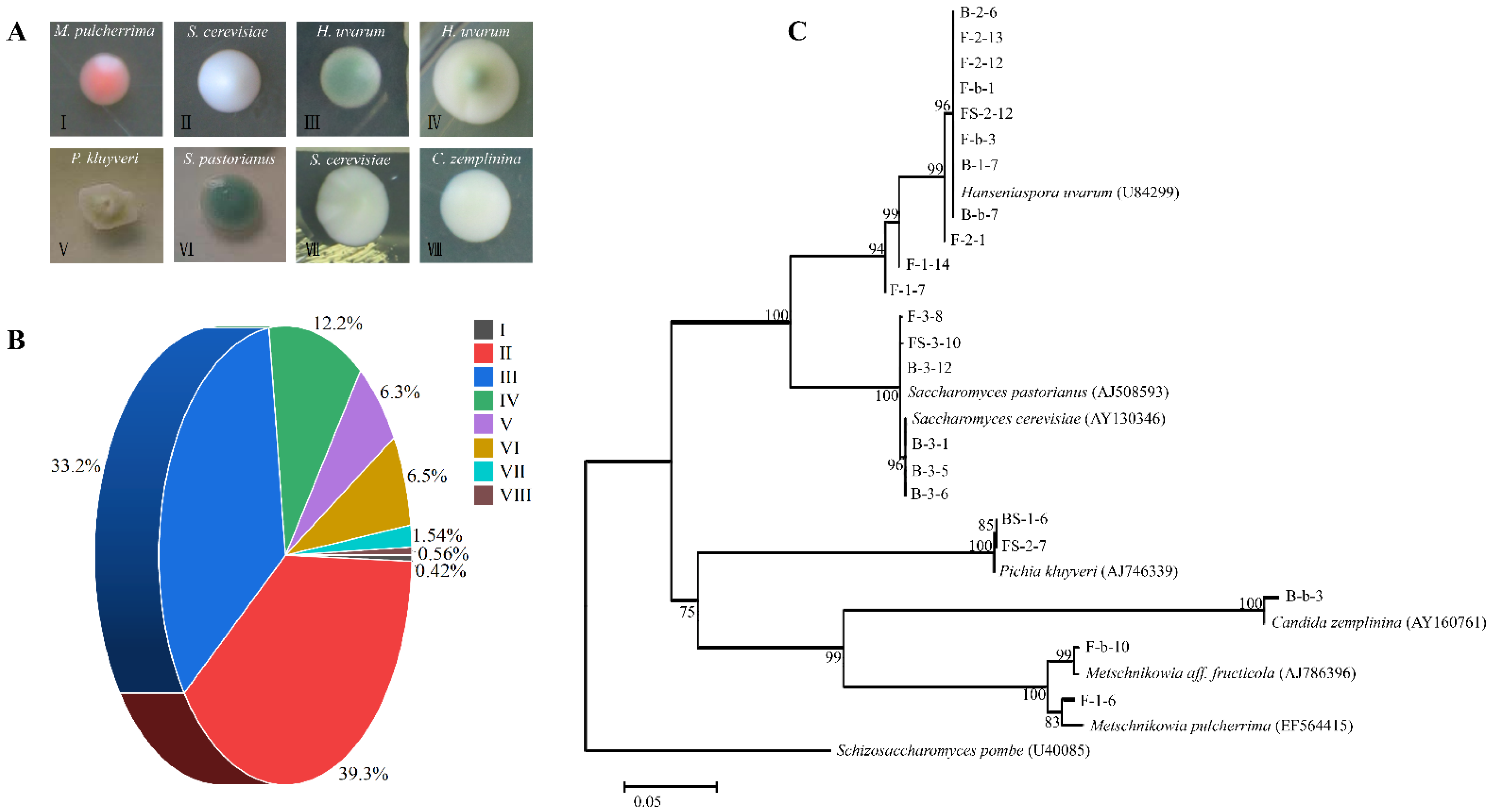
3.1. General Yeast Population Profile
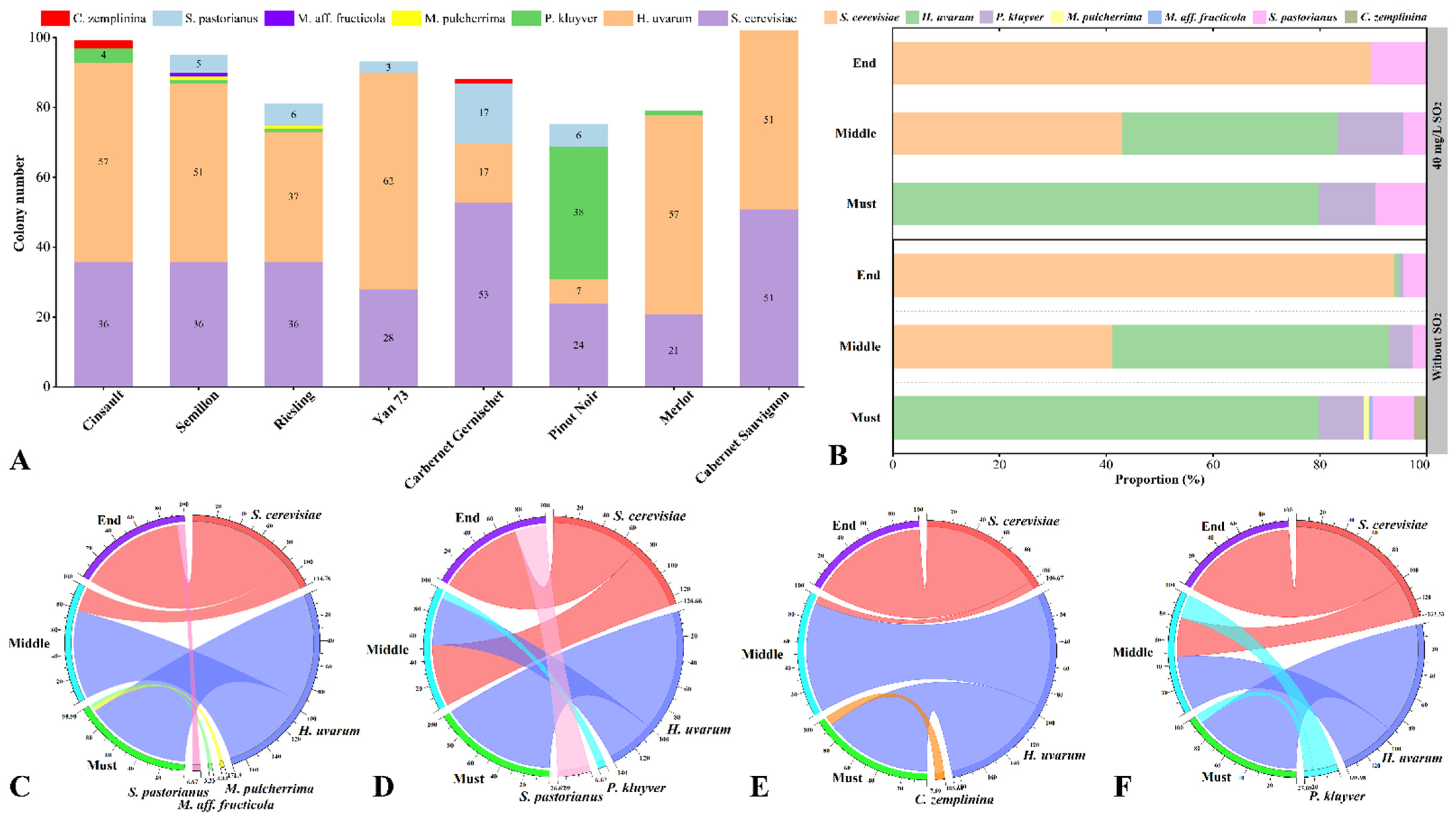
3.2. Yeast Population Dynamics during Spontaneous Fermentation
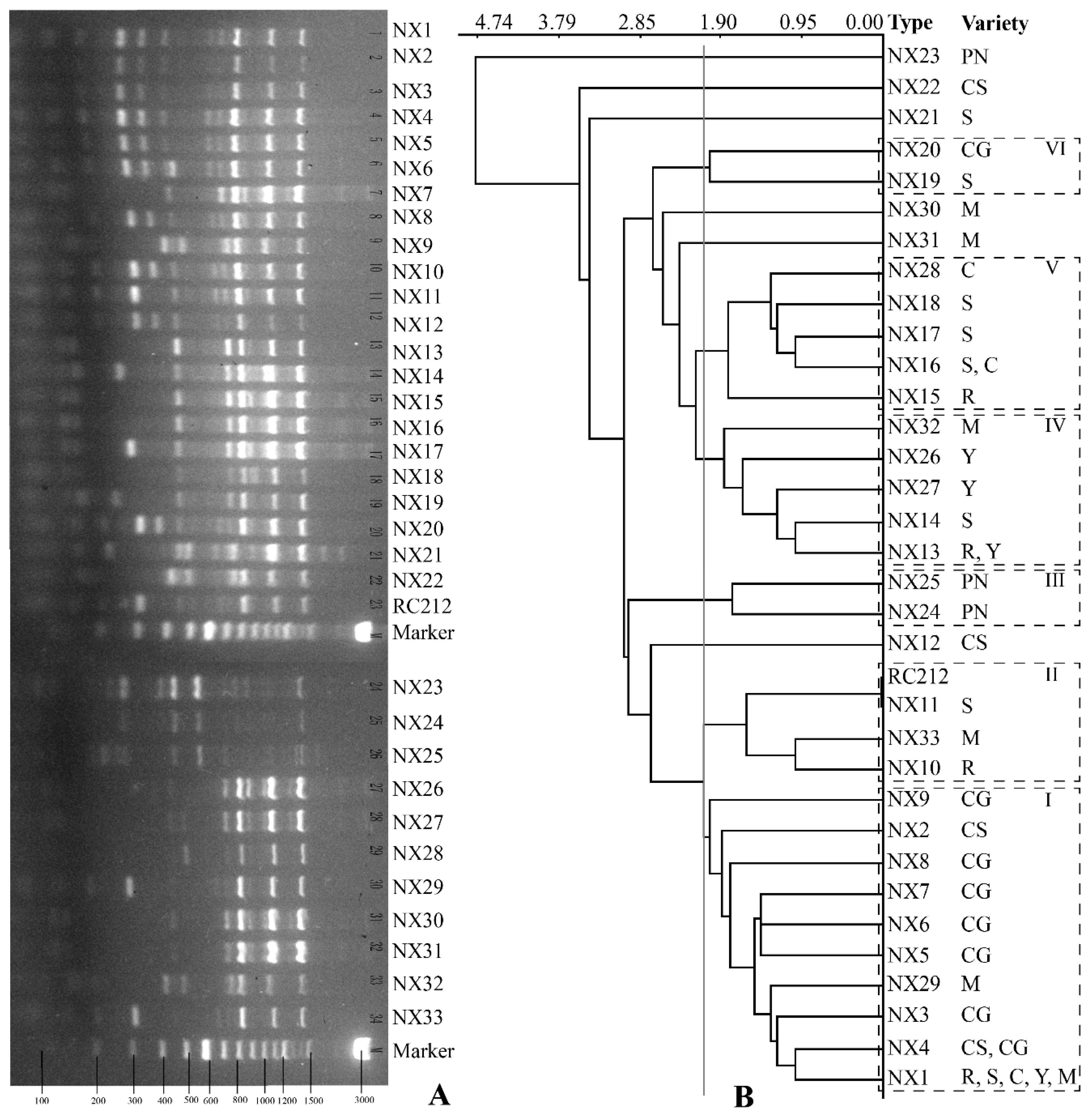
3.3. Genetic Diversity of S. cerevisiae by Interdelta Polymorphism Fingerprinting
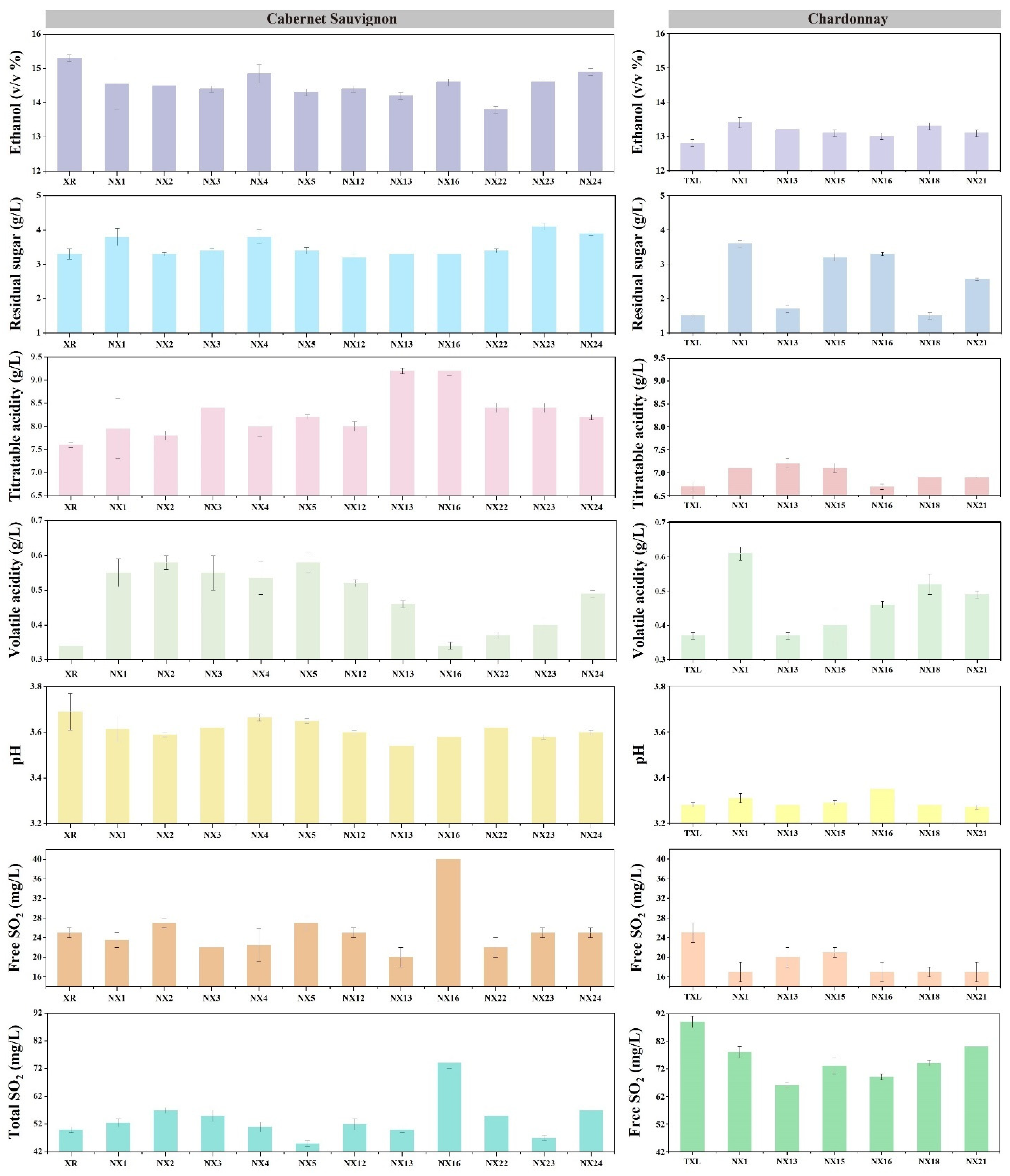
3.4. Vinification Using Indigenous S. cerevisiae Strains with Differentiated Genotypes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
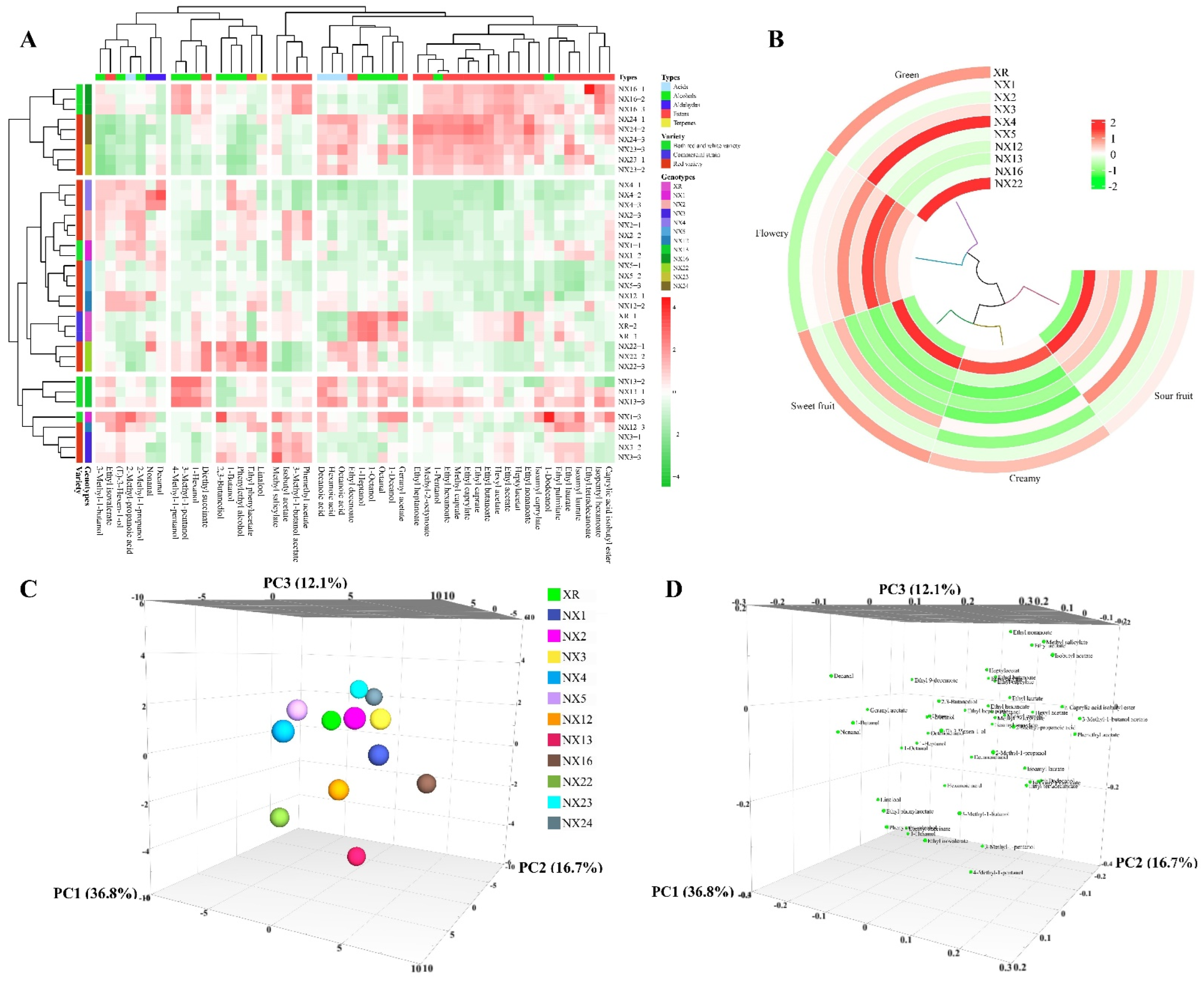{kind=link}
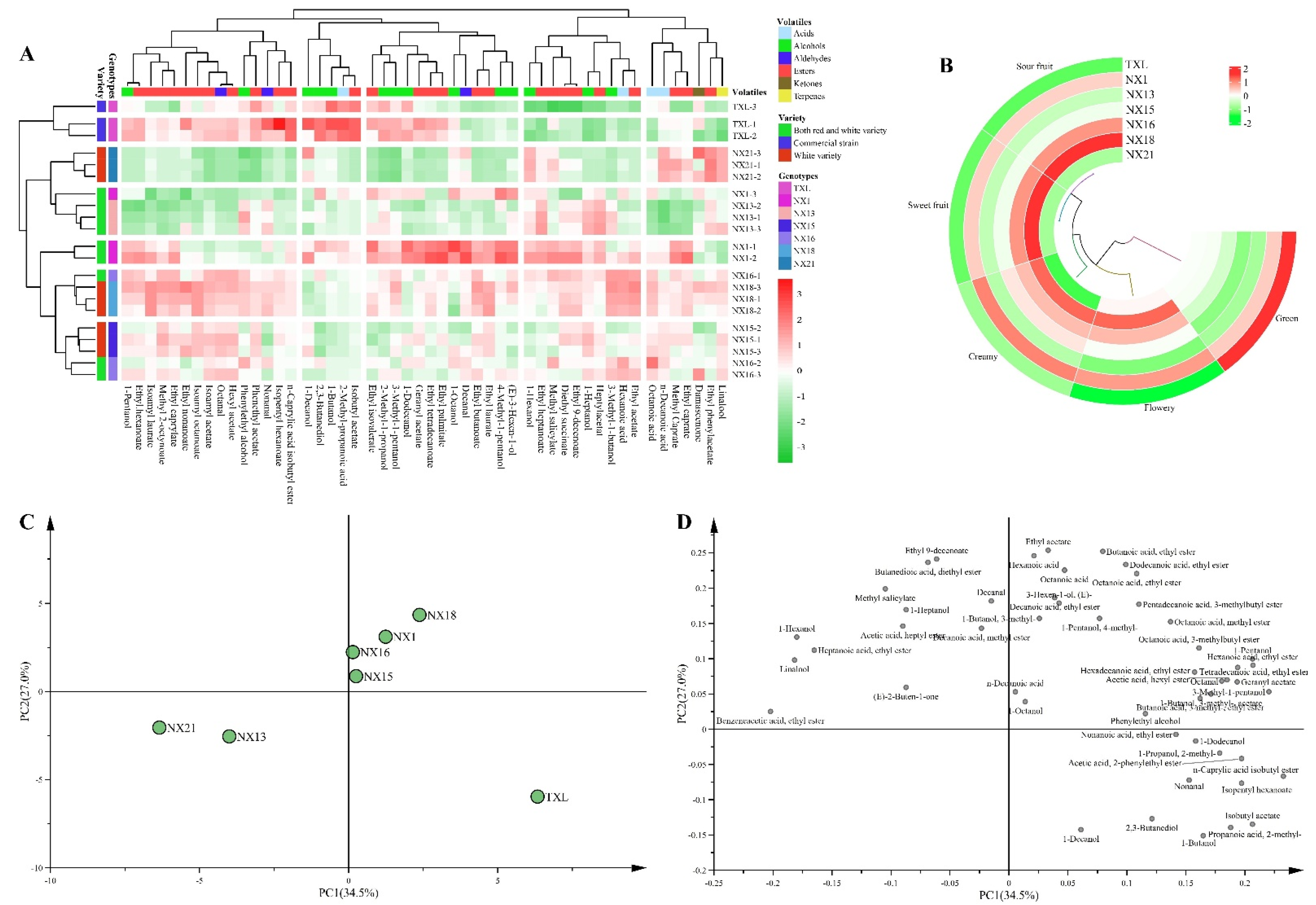{kind=link}
| Compounds | XR | NX1 | NX2 | NX3 | NX4 | NX5 | NX12 | NX13 | NX16 | NX22 | NX23 | NX24 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Higher alcohols | ||||||||||||
| Phenylethyl alcohol | 167,392.81 ± 8101.51 b | 166,138.4 ± 16,137.44 b | 184,165.29 ± 1262.85 b | 163,080.39 ± 5833.96 b | 181,400.58 ± 13,807.16 b | 153,952.72 ± 6061.12 bc | 161,268.44 ± 2111.01 b | 172,947.88 ± 9876.84 b | 170,013.97 ± 5498.8 b | 219,079.07 ± 20,367.73 a | 122,070.6 ± 4850.72 c | 122,826.97 ± 3810.22 c |
| 1-Butanol | 4032.25 ± 55.28 c | 3255.38 ± 191.63 ghi | 3506.21 ± 41.77 efg | 3866.75 ± 148.69 cde | 4516.09 ± 38.29 b | 3612.9 ± 91.64 fg | 3327.66 ± 123.64 hi | 3014.35 ± 104.99 i | 3470.17 ± 63.07 gh | 4524.09 ± 77.16 b | 2634.42 ± 48.23 j | 3126.15 ± 87.79 hi |
| 1-Hexanol | 5317.06 ± 25.64 b | 4570.32 ± 91.29 fg | 4300.47 ± 49.38 ghi | 4243.81 ± 55.95 hi | 4887.98 ± 9.76 ef | 4610.74 ± 60.89 fg | 4961.53 ± 11.53 cd | 5975.9 ± 185.07 a | 4925.24 ± 74.53 de | 5258.73 ± 58.39 bc | 4358.67 ± 89.23 gh | 5012.94 ± 105.5 cd |
| 2,3-Butanediol | 111,590.44 ± 3845.3 ef | 123,330.57 ± 14,588.11 de | 98,891.31 ± 10,380.16 ef | 132,756.81 ± 19,232.17 de | 90,805.68 ± 7400.75 ef | 81,275.53 ± 6401.69 ef | 77,136.82 ± 4803.14 ef | 57,625.82 ± 1910.63 f | 108,578.58 ± 14,651.52 ef | 162,414.27 ± 18,866.1 bc | 96,817.49 ± 12,192.06 ef | 92,397.23 ± 3723.52 ef |
| 3-Methyl-1-butanol | 380,702.13 ± 4759.75 de | 387,432.53 ± 8767 cd | 414,953.11 ± 3126 ab | 387,767.4 ± 5091.16 cd | 416,610.34 ± 2661.54 a | 353,892.33 ± 3751.61 ef | 379,568.41 ± 2894.13 de | 401,987.72 ± 14,710.3 cd | 401,066.72 ± 5778.98 cd | 378,320.26 ± 3211.6 de | 287,741.45 ± 5179.77 g | 305,563.44 ± 6784.64 g |
| Esters | ||||||||||||
| Ethyl acetate | 141,076.1 ± 570.96 b | 122,903.61 ± 2959.39 de | 121,830.71 ± 783.22 fg | 137,144.55 ± 2347.51 b | 113,410.61 ± 606.22 gh | 115,354.37 ± 1619.63 gh | 110,158.64 ± 851.51 h | 114,443.65 ± 3603.46 gh | 155,122.62 ± 2309.6 a | 113,190.06 ± 735.67 gh | 135,861.31 ± 2615.16 bc | 150,842.72 ± 2602.23 a |
| Ethyl butanoate | 334.9 ± 11.72 ef | 259.71 ± 16.41 jk | 222.11 ± 3.17 k | 305.62 ± 11.79 fg | 276.33 ± 5.71 hi | 294.94 ± 13.39 gh | 342.9 ± 8.56 de | 441.1 ± 20.65 b | 401.13 ± 14.64 bc | 550.72 ± 6.63 a | 271.19 ± 7.64 ghij | 369.46 ± 10.9 cd |
| Ethyl caprate | 639.06 ± 0.87 cde | 552.32 ± 6.67 efghi | 533.78 ± 11.62 fghi | 523.65 ± 17.35 ghi | 470.09 ± 16.86 i | 501.33 ± 5.29 hi | 497.1 ± 24.02 hi | 587.38 ± 74.11 defgh | 710.46 ± 16.93 bc | 581.51 ± 15.24 defgh | 751 ± 44.33 ab | 828.31 ± 38.13 a |
| Ethyl caprylate | 2346.69 ± 19.12 cd | 2081.53 ± 30.68 de | 2101.35 ± 7.97 de | 2264.03 ± 66.28 cd | 1895.57 ± 25.46 e | 2097.47 ± 27.7 de | 2250.86 ± 31.97 cde | 2325.33 ± 303.39 cd | 2771.84 ± 39.05 b | 2246.45 ± 36.88 cde | 2796.76 ± 97.31 b | 3243.46 ± 105.34 a |
| Ethyl heptanoate | 14.8 ± 3.21 def | 15.02 ± 4.25 def | 13.97 ± 1.13 efg | 17.25 ± 1.18 cd | 13.15 ± 4.11 fg | 16.14 ± 3.41 cde | 17.01 ± 2.24 cd | 23 ± 2.16 b | 17.74 ± 2.08 c | 16.96 ± 4.31 cd | 24.51 ± 2.65 b | 27.16 ± 0.94 a |
| Ethyl hexanoate | 1399.44 ± 8.17 fg | 1407.03 ± 41.28 fg | 1328.87 ± 24.21 g | 1628.04 ± 28.63 de | 1341.56 ± 5.21 g | 1522.38 ± 40.04 ef | 1506.59 ± 15.63 ef | 1761.7 ± 69.05 cd | 1897.43 ± 22.3 bc | 1585.58 ± 31.68 e | 1994.14 ± 39.93 ab | 2130.84 ± 53.18 a |
| Ethyl isovalerate | 190.24 ± 1.19 cd | 190.76 ± 0.31 cd | 216.75 ± 10.97 bc | 218.99 ± 5.04 bc | 242.63 ± 5.71 ab | 222.18 ± 8.14 bc | 260.02 ± 3.02 a | 246.26 ± 11.62 ab | 222 ± 16.6 bc | 214.27 ± 10.6 bc | 150.54 ± 9.56 e | 155.04 ± 12.96 de |
| Fatty acids | ||||||||||||
| Hexanoic acid | 1787.69 ± 43.28 f | 1959.13 ± 180.21 def | 1836.25 ± 31.86 f | 2041.96 ± 66.68 cdef | 1876.05 ± 97.95 ef | 2054.75 ± 102.65 cdef | 2194.66 ± 14.1 abcde | 2479.7 ± 97.51 a | 2102.49 ± 58.76 bcdef | 2279.8 ± 126.28 abcd | 2251.77 ± 139.77 abcd | 2390.67 ± 88.19 ab |
| Octanoic acid | 1308.31 ± 30.95 ef | 1376.76 ± 40.1 def | 1425.21 ± 18.1 cdef | 1384.72 ± 42.99 def | 1257.54 ± 77.07 f | 1456.28 ± 50.16 bcde | 1515.97 ± 3.99 abcd | 1601.95 ± 73.22 abc | 1369.78 ± 41.93 def | 1598.34 ± 99.93 abc | 1710.75 ± 72.14 a | 1639.71 ± 58.75 ab |
| Carbonyl Compounds | ||||||||||||
| Octanal | 10.09 ± 1.19 ab | 3.49 ± 1.8 ab | 7.23 ± 0.19 ab | 6.17 ± 3.59 ab | 3.43 ± 0.33 ab | 5.99 ± 0.17 ab | 4.85 ± 1.84 ab | 11.35 ± 1.09 ab | 2.06 ± 0.56 b | 3.79 ± 2 ab | 9.37 ± 3.57 ab | 4.16 ± 2.33 ab |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the Plant Microbiome: Looking Back and Future Perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial Terroir for Wine Grapes. Proc. Natl. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H. Wine Yeasts for the Future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Lin, M.; Guo, S.; Yang, S.; Han, X.; Ren, M.; Song, Y.; Du, L.; You, Y.; Zhan, J.; et al. A Fundamental Landscape of Fungal Biogeographical Patterns across the Main Chinese Wine-Producing Regions and the Dominating Shaping Factors. Food Res. Int. 2021, 150, 110736. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, P.; Chen, D.; Howell, K. From the Vineyard to the Winery: How Microbial Ecology Drives Regional Distinctiveness of Wine. Front. Microbiol. 2019, 10, 2679. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of Saccharomyces cerevisiae Yeasts Associated to Spontaneously Fermenting Grapes from an Italian “Heroic Vine-Growing Area”. Food Microbiol. 2012, 31, 159–166. [Google Scholar] [CrossRef]
- Gao, F.; Chen, J.; Xiao, J.; Cheng, W.; Zheng, X.; Wang, B.; Shi, X. Microbial Community Composition on Grape Surface Controlled by Geographical Factors of Different Wine Regions in Xinjiang, China. Food Res. Int. 2019, 122, 348–360. [Google Scholar] [CrossRef]
- Regodón, J.; Pérez, F.; Valdés, M.; De Miguel, C.; Ramırez, M. A Simple and Effective Procedure for Selection of Wine Yeast Strains. Food Microbiol. 1997, 14, 247–254. [Google Scholar] [CrossRef]
- Alexandre, H. Wine Yeast Terroir: Separating the Wheat from the Chaff—For an Open Debate. Microorganisms 2020, 8, 787. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial Biogeography of Wine Grapes is Conditioned by Cultivar, Vintage, and Climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine Fermentation Microbiome: A Landscape from Different Portuguese Wine Appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of Non-Saccharomyces Yeast Strains for Reducing Alcohol Levels in Wine by Sugar Respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renouf, V.; Miot-Sertier, C.; Strehaiano, P.; Lonvaud-Funel, A. The Wine Microbial Consortium: A Real Terroir Characteristic. Oeno One 2006, 40, 209–216. [Google Scholar] [CrossRef]
- Feng, L.; Wang, J.; Ye, D.; Song, Y.; Qin, Y.; Liu, Y. Yeast Population Dynamics During Spontaneous Fermentation of Icewine and Selection of Indigenous Saccharomyces cerevisiae Strains for the Winemaking in Qilian, China. J. Sci. Food Agric. 2020, 100, 5385–5394. [Google Scholar] [CrossRef]
- Longo, E.; Cansado, J.; Agrelo, D.; Villa, T.G. Effect of Climatic Conditions on Yeast Diversity in Grape Musts from Northwest Spain. Am. J. Enol. Vitic. 1991, 42, 141–144. [Google Scholar]
- Pretorius, I.S. Tailoring Wine Yeast for the New Millennium: Novel Approaches to the Ancient Art of Winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Sun, Y.; Qin, Y.; Pei, Y.; Wang, G.; Joseph, C.L.; Bisson, L.F.; Liu, Y. Evaluation of Chinese Saccharomyces cerevisiae Wine Strains from Different Geographical Origins. Am. J. Enol. Vitic. 2017, 68, 73–80. [Google Scholar] [CrossRef] [Green Version]
- González-Alonso, I.; Walker, M.E.; Vallejo-Pascual, M.-E.; Naharro-Carrasco, G.; Jiranek, V. Capturing Yeast Associated with Grapes and Spontaneous Fermentations of the Negro Saurí Minority Variety from an Experimental Vineyard near León. Sci. Rep. 2021, 11, 3748. [Google Scholar] [CrossRef]
- Liu, D.; Legras, J.-L.; Zhang, P.; Chen, D.; Howell, K. Diversity and Dynamics of Fungi during Spontaneous Fermentations and Association with Unique Aroma Profiles in Wine. Int. J. Food Microbiol. 2021, 338, 108983. [Google Scholar] [CrossRef] [PubMed]
- Clowers, K.J.; Heilberger, J.; Piotrowski, J.S.; Will, J.L.; Gasch, A.P. Ecological and Genetic Barriers Differentiate Natural Populations of Saccharomyces cerevisiae. Mol. Biol. Evol. 2015, 32, 2317–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannakou, K.; Cotterrell, M.; Delneri, D. Genomic Adaptation of Saccharomyces species to Industrial Environments. Front. Genet. 2020, 11, 916. [Google Scholar] [CrossRef] [PubMed]
- Legras, J.-L.; Galeote, V.; Bigey, F.; Camarasa, C.; Marsit, S.; Nidelet, T.; Sanchez, I.; Couloux, A.; Guy, J.; Franco-Duarte, R.; et al. Adaptation of S. cerevisiae to Fermented Food Environments Reveals Remarkable Genome Plasticity and the Footprints of Domestication. Mol. Biol. Evol. 2018, 35, 1712–1727. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romano, P. Biotechnological Approach Based on Selected Saccharomyces cerevisiae Starters for Reducing the Use of Sulfur Dioxide in Wine. Microorganisms 2020, 8, 738. [Google Scholar] [CrossRef]
- Morgan, S.C.; Scholl, C.M.; Benson, N.L.; Stone, M.L.; Durall, D.M. Sulfur Dioxide Addition at Crush Alters Saccharomyces cerevisiae strain Composition in Spontaneous Fermentations at Two Canadian Wineries. Int. J. Food Microbiol. 2017, 244, 96–102. [Google Scholar] [CrossRef]
- Scacco, A.; Oliva, D.; Di Maio, S.; Polizzotto, G.; Genna, G.; Tripodi, G.; Lanza, C.M.; Verzera, A. Indigenous Saccharomyces cerevisiae strains and Their Influence on the Quality of Cataratto, Inzolia and Grillo White Wines. Food Res. Int. 2012, 46, 1–9. [Google Scholar] [CrossRef]
- Suzzi, G.; Arfelli, G.; Schirone, M.; Corsetti, A.; Perpetuini, G.; Tofalo, R. Effect of Grape Indigenous Saccharomyces cerevisiae strains on Montepulciano D’abruzzo Red Wine Quality. Food Res. Int. 2012, 46, 22–29. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional Microbial Signatures Positively Correlate with Differential Wine Phenotypes: Evidence for a Microbial Aspect to Terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef] [Green Version]
- Drumonde-Neves, J.; Franco-Duarte, R.; Vieira, E.; Mendes, I.; Lima, T.; Schuller, D.; Pais, C. Differentiation of Saccharomyces cerevisiae Populations from Vineyards of the Azores Archipelago: Geography Vs Ecology. Food Microbiol. 2018, 74, 151–162. [Google Scholar] [CrossRef]
- Bagheri, B.; Philipp, C.; Horacek, M.; Bauer, F.; Setati, M. Microbial Diversity in Grape Musts from Austrian and South African Grape Varieties and Regions. BIO Web Conf. 2019, 12, 02028. [Google Scholar] [CrossRef]
- Capece, A.; Granchi, L.; Guerrini, S.; Mangani, S.; Romaniello, R.; Vincenzini, M.; Romano, P. Diversity of Saccharomyces cerevisiae Strains Isolated from Two Italian Wine-Producing Regions. Front. Microbiol. 2016, 7, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Ding, Y.; Gao, F.; Zhang, L.; Wang, L.; Li, H.; Wang, H. Community Succession of the Grape Epidermis Microbes of Cabernet Sauvignon (Vitis Vinifera L.) from Different Regions in China During Fruit Development. Int. J. Food Microbiol. 2022, 362, 109475. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, W.; Huang, X.; Xu, Y. Investigation of Yeast Population Diversity and Dynamics in Spontaneous Fermentation of Vidal Blanc Icewine by Traditional Culture-Dependent and High-Throughput Sequencing Methods. Food Res. Int. 2018, 112, 66–77. [Google Scholar] [CrossRef]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to Profile Native Flora Fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- Feng, L.; Jia, H.; Wang, J.; Qin, Y.; Liu, Y.; Song, Y. Selection of Indigenous Saccharomyces cerevisiae strains for Winemaking in Northwest China. Am. J. Enol. Vitic. 2019, 70, 115–126. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.; Yi, H.; Wang, B.; Xiao, J.; Zhou, X.; Jiankun, X.; Jiang, L.; Shi, X. Microbial Community Composition and Its Role in Volatile Compound Formation During the Spontaneous Fermentation of Ice Wine Made from Vidal Grapes. Process Biochem. 2020, 92, 365–377. [Google Scholar] [CrossRef]
- OIV. Compendium of International Methods of Analysis of Wines and Musts; International Organisation of Vine and Wine Paris: Paris, France, 2020. [Google Scholar]
- Lan, Y.-B.; Qian, X.; Yang, Z.-J.; Xiang, X.-F.; Yang, W.-X.; Liu, T.; Zhu, B.-Q.; Pan, Q.-H.; Duan, C.-Q. Striking Changes in Volatile Profiles at Sub-Zero Temperatures during Over-ripening of ‘Beibinghong’ grapes in Northeastern China. Food Chem. 2016, 212, 172–182. [Google Scholar] [CrossRef]
- González-Álvarez, M.; Noguerol-Pato, R.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J. Sensory Description of Sweet Wines Obtained by the Winemaking Procedures of Raisining, Botrytisation and Fortification. Food Chem. 2014, 145, 1021–1030. [Google Scholar] [CrossRef]
- Hu, K.; Jin, G.-J.; Xu, Y.-H.; Tao, Y.-S. Wine Aroma Response to Different Participation of Selected Hanseniaspora uvarum in Mixed Fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef]
- Welke, J.E.; Zanus, M.; Lazzarotto, M.; Zini, C.A. Quantitative Analysis of Headspace Volatile Compounds Using Comprehensive Two-Dimensional Gas Chromatography and Their Contribution to the Aroma of Chardonnay Wine. Food Res. Int. 2014, 59, 85–99. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Chen, Y.; Li, J.; Shi, X.; Deng, L.; Wang, B. Evaluation of the Effect of Auxiliary Starter Yeasts with Enzyme Activities on Kazak Cheese Quality and Flavor. Front. Microbiol. 2020, 11, 614208. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Mendoza, L.M.; Neef, A.; Vignolo, G.; Belloch, C. Yeast Diversity during the Fermentation of Andean Chicha: A Comparison of High-Throughput Sequencing and Culture-Dependent Approaches. Food Microbiol. 2017, 67, 1–10. [Google Scholar] [CrossRef]
- Lin, M.M.-H.; Boss, P.K.; Walker, M.E.; Sumby, K.M.; Grbin, P.R.; Jiranek, V. Evaluation of Indigenous Non-Saccharomyces Yeasts Isolated from a South Australian Vineyard for Their Potential as Wine Starter Cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef] [PubMed]
- De-La-Fuente-Blanco, A.; Sáenz-Navajas, M.-P.; Ferreira, V. On the Effects of Higher Alcohols on Red Wine Aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-T.; Hsieh, C.-W.; Lo, Y.-C.; Liou, B.-K.; Lin, H.-W.; Hou, C.-Y.; Cheng, K.-C. Isolation and Identification of Aroma-Producing Non-Saccharomyces Yeast Strains and the Enological Characteristic Comparison in Wine Making. LWT-Food Sci. Technol. 2022, 154, 112653. [Google Scholar] [CrossRef]
- Cureau, N.; Threlfall, R.; Carbonero, F.; Howard, L.; Lavefve, L. Fungal Diversity and Dynamics during Grape Wine Fermentations with Different Sulfite Levels and Yeast Inoculations. Am. J. Enol. Vitic. 2021, 72, 240–256. [Google Scholar] [CrossRef]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef]
- Chalvantzi, I.; Banilas, G.; Tassou, C.; Nisiotou, A. Patterns of Genetic Diversity and the Invasion of Commercial Starters in Saccharomyces cerevisiae Vineyard Populations of Santorini Island. Foods 2020, 9, 561. [Google Scholar] [CrossRef]
- Viel, A.; Legras, J.-L.; Nadai, C.; Carlot, M.; Lombardi, A.; Crespan, M.; Migliaro, D.; Giacomini, A.; Corich, V. The Geographic Distribution of Saccharomyces cerevisiae Isolates within Three Italian Neighboring Winemaking Regions Reveals Strong Differences in Yeast Abundance, Genetic Diversity and Industrial Strain Dissemination. Front. Microbiol. 2017, 8, 1595. [Google Scholar] [CrossRef] [Green Version]
- Goddard, M.R.; Anfang, N.; Tang, R.; Gardner, R.C.; Jun, C. A Distinct Population of Saccharomyces cerevisiae in New Zealand: Evidence for Local Dispersal by Insects and Human-Aided Global Dispersal in Oak Barrels. Environ. Microbiol. 2010, 12, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.S.; Cozzolino, D.; Bartowsky, E.J.; Fleet, G.H.; Henschke, P.A. Metabolic Profiling as a Tool for Revealing Saccharomyces Interactions During Wine Fermentation. FEMS Yeast Res. 2006, 6, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea Thermotolerans on Chemical Composition and Sensory Profiles of Merlot Wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.M.; Walker, M.E.; Boss, P.K.; Jiranek, V. The Effect of Grape Juice Dilution and Complex Nutrient Addition on Oenological Fermentation and Wine Chemical Composition. J. Food Compos. Anal. 2022, 105, 104241. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial Modulation of Aromatic Esters in Wine: Current Knowledge and Future Prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.-M.; Van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]






| Type | Riesling | Semillon | Cabernet Sauvignon | Cabernet Gernischet | Cinsault | Pinot Noir | Yan73 | Merlot | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NS | S | NS | S | NS | S | NS | S | NS | S | NS | S | NS | S | NS | S | |||||||||||||||
| M | E | M | E | M | E | M | E | M | E | M | E | M | E | I | M | E | M | E | M | E | M | E | E | M | E | E | E | M | E | |
| NX1 | 3 (38) | 8 (62) | 7 (88) | 7 | 1 (17) | 2 (17) | 1(14) | 5(46) | - | - | - | - | - | - | - | - | - | - | 5 | 13 | - | - | - | 1 | - | - | 17 (65) | 1 | 16 (88) | |
| NX2 | - | - | - | - | - | - | - | - | 1 (12) | 2 (17) | 2 (14) | 1 (7) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| NX3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 7 (50) | 6 (55) | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| NX4 | - | - | - | - | - | - | - | - | 7 (88) | 7 (58) | 11 (79) | 10 (72) | 7 (70) | 7 (50) | 3 (75) | 7 (50) | 3 (27) | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX5 | - | - | - | - | - | - | - | - | - | - | - | - | 1 (10) | 5 (36) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX6 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (7) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (9) | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX8 | - | - | - | - | - | - | - | - | - | - | - | - | 2 (20) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX9 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (7) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX10 | - | - | 1 (12) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX11 | - | - | - | - | - | 1 (8) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX12 | - | - | - | - | - | - | - | - | - | 2 (17) | 1 (7) | 2 (14) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX13 | 5 (62) | 4 (30) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 11 (84) | 11 (92) | - | - | - |
| NX14 | - | - | - | - | - | - | 1 (14) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX15 | - | 1 (8) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX16 | - | - | - | - | 5 (83) | 7 (58) | 4 (58) | 2 | - | - | - | - | - | - | - | - | - | 1 | 2(50) | - | - | - | - | - | - | - | - | - | - | - |
| NX17 | - | - | - | - | - | 1 (8) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX18 | - | - | - | - | - | - | - | 4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX19 | - | - | - | - | - | 1 (8) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (25) | - | 1 (9) | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX21 | - | - | - | - | - | - | 1 (14) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX22 | - | - | - | - | - | - | - | - | - | 1 (8) | - | 1 (7) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| NX23 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (50) | 11 (92) | 9 | - | - | - | - | - | - |
| NX24 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (50) | - | - | - | - | - | - | - | - |
| NX25 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (8) | - | - | - | - | - | - | - |
| NX26 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (8) | - | - | - | - |
| NX27 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (8) | 1 (8) | - | - | - |
| NX28 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 (50) | - | - | - | - | - | - | - | - | - | - | - |
| NX29 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 (8) | - | - |
| NX30 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 (8) | - | - |
| NX31 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 (11) | - | 1 (6) |
| NX32 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 (8) | - | - |
| NX33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 (6) |
| Total | 8 | 13 | 8 | 7 | 6 | 12 | 7 | 11 | 8 | 12 | 14 | 14 | 10 | 14 | 4 | 14 | 11 | 1 | 4 | 5 | 13 | 2 | 12 | 9 | 1 | 13 | 12 | 26 | 1 | 18 |
| 290 | ||||||||||||||||||||||||||||||
| Compounds | TXL | NX1 | NX13 | NX15 | NX16 | NX18 | NX21 |
|---|---|---|---|---|---|---|---|
| Higher alcohols | |||||||
| 1-Hexanol | 1971.76 ± 147.92 f | 2842.83 ± 146.66 ab | 2651.52 ± 74.29 abc | 2309.15 ± 72.57 e | 2628.71 ± 85.34 abcd | 2570.46 ± 37.14 bcde | 2918.71 ± 13.84 a |
| 1-Butanol | 2226.23 ± 191.1 a | 1294.55 ± 193.17 b | 878.94 ± 74.01 cde | 788.7 ± 53.8 e | 994.74 ± 26.87 bcde | 1129.32 ± 15.8 bcd | 1099 ± 48.95 bcde |
| Phenylethyl alcohol | 33,452.18 ± 217.87 bc | 30,548.64 ± 920.91 bcde | 34,624.57 ± 859.49 b | 31,241.83 ± 746.26 bcd | 34,075.19 ± 1382.96 b | 34,578.42 ± 73.79 b | 27,445.21 ± 781.72 def |
| 2,3-Butanediol | 62,208.64 ± 7896.55 a | 52,996.87 ± 6709.23 abc | 42,002.45 ± 2192.48 abcd | 29,209.82 ± 1752.84 d | 44,376.28 ± 6013.81 abcd | 37,672.03 ± 1337.7 bcd | 41,197.97 ± 876.81 abcd |
| Esters | |||||||
| Ethyl butanoate | 1437.82 ± 51.97 de | 1909.82 ± 107.85 b | 1540.11 ± 56.39 cd | 1751.26 ± 72.09 bc | 1802.22 ± 100.98 b | 1965.09 ± 67.34 ab | 1384.46 ± 4.52 de |
| Ethyl heptanoate | 2.22 ± 0.2 h | 5.93 ± 0.59 ab | 6.34 ± 0.29 a | 4.17 ± 0.11 cde | 4.65 ± 0.41 cd | 4.38 ± 0.24 cd | 4.91 ± 0.02 bc |
| Ethyl hexanoate | 3314.12 ± 207.96 ab | 3309.02 ± 261.28 ab | 2450.15 ± 113.84 e | 3044.31 ± 115.22 bcd | 3109.39 ± 214.3 bc | 3236.18 ± 139.05 b | 2705.37 ± 0.34 cde |
| Ethyl acetate | 124,257.25 ± 5737.59 efgh | 154,587.96 ± 11,783.89 bcd | 137,846.7 ± 4915.97 de | 144,759.19 ± 5168.89 cde | 163,631.1 ± 7155.4 bc | 175,981.76 ± 4627.54 ab | 132,947.74 ± 934.09 defg |
| Ethyl caprylate | 5949.03 ± 668.51 bcd | 6576.43 ± 951.39 bc | 5287.53 ± 169.86 cde | 6646.79 ± 157.89 bc | 6695.31 ± 402.05 bc | 7301.32 ± 195.72 ab | 5847.88 ± 10.79 bcd |
| Ethyl caprate | 2509.32 ± 368.15 bcde | 3258.82 ± 550.69 ab | 2031.23 ± 42.4 de | 2915.93 ± 86.19 abcd | 2761.74 ± 167.02 abcd | 3141.74 ± 15.44 ab | 2951.21 ± 162.31 abc |
| Fatty acids | |||||||
| Octanoic acid | 5089.21 ± 373.88 fghi | 5782.67 ± 160.38 cdefg | 4734.89 ± 72.96 hi | 5864.91 ± 159.48 bcdef | 6352.34 ± 414.9 abcd | 6214.38 ± 93.05 abcde | 5480.99 ± 169.41 efghi |
| Hexanoic acid | 3699.21 ± 17.38 e | 4526.67 ± 195.94 bc | 4003.64 ± 158.99 de | 4202.46 ± 160.85 cd | 5018.69 ± 100.03 a | 5016.69 ± 69.15 a | 4035 ± 85.38 de |
| Carbonyl Compounds | |||||||
| Octanal | 315.83 ± 16.9 b | 257.69 ± 22.57 c | 240.71 ± 9.58 c | 324.74 ± 10.24 b | 321.02 ± 19.74 b | 322.93 ± 11.01 b | 226.68 ± 0.17 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Jiang, J.; Song, Y.; Zang, X.; Wang, G.; Pei, Y.; Song, Y.; Qin, Y.; Liu, Y. Yeast Diversity during Spontaneous Fermentations and Oenological Characterisation of Indigenous Saccharomyces cerevisiae for Potential as Wine Starter Cultures. Microorganisms 2022, 10, 1455. https://doi.org/10.3390/microorganisms10071455
Chen Y, Jiang J, Song Y, Zang X, Wang G, Pei Y, Song Y, Qin Y, Liu Y. Yeast Diversity during Spontaneous Fermentations and Oenological Characterisation of Indigenous Saccharomyces cerevisiae for Potential as Wine Starter Cultures. Microorganisms. 2022; 10(7):1455. https://doi.org/10.3390/microorganisms10071455
Chicago/Turabian StyleChen, Yu, Jiao Jiang, Yaoyao Song, Xiaomin Zang, Guoping Wang, Yingfang Pei, Yuyang Song, Yi Qin, and Yanlin Liu. 2022. "Yeast Diversity during Spontaneous Fermentations and Oenological Characterisation of Indigenous Saccharomyces cerevisiae for Potential as Wine Starter Cultures" Microorganisms 10, no. 7: 1455. https://doi.org/10.3390/microorganisms10071455