
Plasmodium falciparum S-Adenosylmethionine Synthetase Is Essential for Parasite Survival through a Complex Interaction Network with Cytoplasmic and Nuclear Proteins

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Identifiers
2.2. Primary Antibodies
2.3. Parasite Culture
2.4. Generation of Mouse Anti-Pf92 Antisera
2.5. Generation of the PfSAMS-HA-KD Parasite Line
2.6. Generation of the PfSAMS-GFP-BirA Parasite Lines
2.7. Indirect Immunofluorescence Assay
2.8. Subcellular Fractioning
2.9. Western Blotting
2.10. Asexual Blood Stage Replication Assay
2.11. Gametocyte Development Assay
2.12. Preparation of Samples for BioID Analysis
2.13. Proteolytic Digestion
2.14. Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis
2.15. Data Analysis and Label-Free Quantification
2.16. Data Availability
2.17. Statistical Analysis
3. Results
3.1. PfSAMS Is Expressed in the Asexual and Sexual Blood Stages of P. falciparum
3.2. PfSAMS Deficiency Impairs Intraerythrocytic Growth and Gametocyte Development
3.3. BirA-Tagging of PfSAMS Results in Protein Biotinylation in Blood Stage Parasites
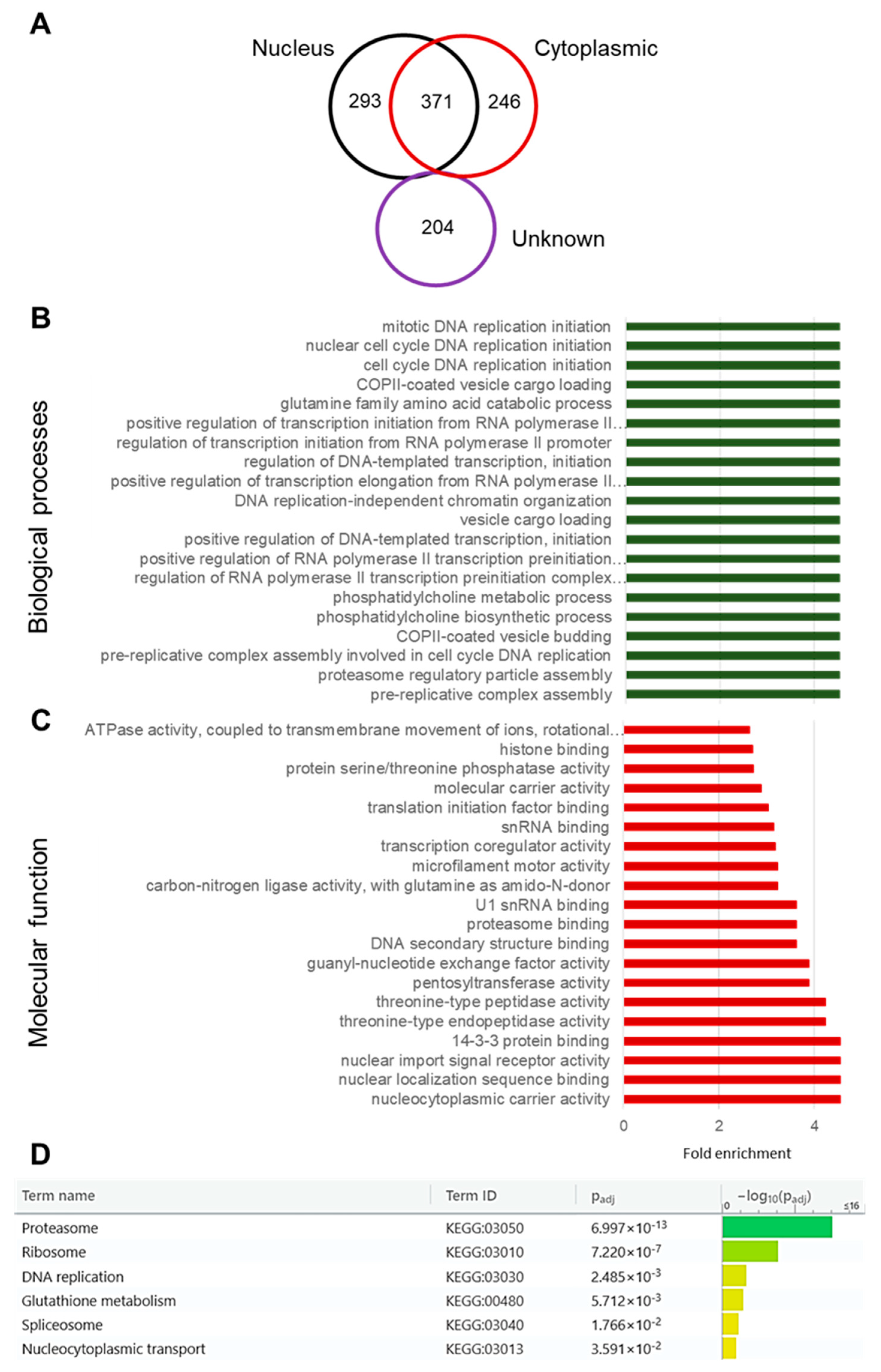
3.4. The PfSAMS Interactome Comprises Nuclear and Cytosolic Proteins Involved in Vital Cellular Functions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, S.C. S-Adenosylmethionine. Int. J. Biochem. Cell Biol. 2000, 32, 391–395. [Google Scholar] [CrossRef]
- Minois, N.; Carmona-Gutierrez, D.; Madeo, F. Polyamines in aging and disease. Aging 2011, 3, 716–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.; Wu, Q.; Li, J.; Sun, S.; Sun, S. S-adenosylmethionine: A metabolite critical to the regulation of autophagy. Cell Prolif. 2020, 53, e12891. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Sun, W.M.; Hwang, J.J.; Stain, S.C.; Lu, S.C. Changes in S-adenosylmethionine synthetase in human liver cancer: Molecular characterization and significance. Hepatology 1996, 24, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Mao, Z.; Hwang, J.J.; Lu, S.C. Differential expression of methionine adenosyltransferase genes influences the rate of growth of human hepatocellular carcinoma cells. Cancer Res. 1998, 58, 1444–1450. [Google Scholar] [PubMed]
- Frau, M.; Feo, F.; Pascale, R.M. Pleiotropic effects of methionine adenosyltransferases deregulation as determinants of liver cancer progression and prognosis. J. Hepatol. 2013, 59, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, C.L.; Kaiser, S.E.; Bolaños, B.; Nowlin, D.; Grantner, R.; Karlicek-Bryant, S.; Feng, J.L.; Jenkinson, S.; Freeman-Cook, K.; Dann, S.G.; et al. Targeting S-adenosylmethionine biosynthesis with a novel allosteric inhibitor of Mat2A. Nat. Chem. Biol. 2017, 13, 785–792. [Google Scholar] [CrossRef]
- Taylor, J.C.; Bock, C.W.; Takusagawa, F.; Markham, G.D. Discovery of novel types of inhibitors of S-adenosylmethionine synthesis by virtual screening. J. Med. Chem. 2009, 52, 5967–5973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Word Malaria Report 2021; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Pretzel, J.; Gehr, M.; Eisenkolb, M.; Wang, L.; Fritz-Wolf, K.; Rahlfs, S.; Becker, K.; Jortzik, E. Characterization and redox regulation of Plasmodium falciparum methionine adenosyltransferase. J. Biochem. 2016, 160, 355–367. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Casandra, D.; Mayho, M.; et al. Uncovering the essential genes of the human malaria parasite Plasmodium falciparum by saturation mutagenesis. Science 2018, 360, eaap7847. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Rubio, J.J.; Riviere, L.; Scherf, A. Shared epigenetic mechanisms control virulence factors in protozoan parasites. Curr. Opin. Microbiol. 2007, 10, 560–568. [Google Scholar] [PubMed]
- Lopez-Rubio, J.-J.; Mancio-Silva, L.; Scherf, A. Genome-wide analysis of heterochromatin associates clonally variant gene regulation with perinuclear repressive centers in malaria parasites. Cell Host Microbe 2009, 5, 179–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petter, M.; Lee, C.C.; Byrne, T.J.; Boysen, K.E.; Volz, J.; Ralph, S.A.; Cowman, A.F.; Brown, G.V.; Duffy, M.F. Expression of P. falciparum var genes involves exchange of the histone variant H2A.Z at the promoter. PLoS Pathog. 2011, 7, e1001292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llinás, M.; Deitsch, K.W.; Voss, T.S. Plasmodium gene regulation: Far more to factor in. Trends Parasitol. 2008, 24, 551–556. [Google Scholar] [CrossRef]
- Cui, L.; Miao, J. Chromatin-mediated epigenetic regulation in the malaria parasite Plasmodium falciparum. Eukaryot. Cell 2010, 9, 1138–1149. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.F.; Selvarajah, S.A.; Josling, G.A.; Petter, M. Epigenetic regulation of the Plasmodium falciparum genome. Brief. Funct. Genomics 2014, 13, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Voss, T.S.; Bozdech, Z.; Bártfai, R. Epigenetic memory takes center stage in the survival strategy of malaria parasites. Curr. Opin. Microbiol. 2014, 20, 88–95. [Google Scholar] [CrossRef]
- Duraisingh, M.T.; Horn, D. Epigenetic Regulation of Virulence Gene Expression in Parasitic Protozoa. Cell Host Microbe 2016, 19, 629–640. [Google Scholar]
- Pérez-Toledo, K.; Rojas-Meza, A.P.; Mancio-Silva, L.; Hernández-Cuevas, N.A.; Delgadillo, D.M.; Vargas, M.; Martínez-Calvillo, S.; Scherf, A.; Hernandez-Rivas, R. Plasmodium falciparum heterochromatin protein 1 binds to tri-methylated histone 3 lysine 9 and is linked to mutually exclusive expression of var genes. Nucleic Acids Res. 2009, 37, 2596–2606. [Google Scholar] [CrossRef]
- Salcedo-Amaya, A.M.; van Driel, M.A.; Alako, B.T.; Trelle, M.B.; van den Elzen, A.M.G.; Cohen, A.M.; Janssen-Megens, E.M.; van de Vegte-Bolmer, M.; Selzer, R.R.; Iniguez, A.L.; et al. Dynamic histone H3 epigenome marking during the intraerythrocytic cycle of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2009, 106, 9655–9660. [Google Scholar] [CrossRef] [Green Version]
- Duraisingh, M.T.; Voss, T.S.; Marty, A.J.; Duffy, M.F.; Good, R.T.; Thompson, J.K.; Freitas-Junior, L.H.; Scherf, A.; Crabb, B.S.; Cowman, A.F. Heterochromatin silencing and locus repositioning linked to regulation of virulence genes in Plasmodium falciparum. Cell 2005, 121, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas-Junior, L.H.; Hernandez-Rivas, R.; Ralph, S.A.; Montiel-Condado, D.; Ruvalcaba-Salazar, O.K.; Rojas-Meza, A.P.; Mâncio-Silva, L.; Leal-Silvestre, R.J.; Gontijo, A.M.; Shorte, S.; et al. Telomeric heterochromatin propagation and histone acetylation control mutually exclusive expression of antigenic variation genes in malaria parasites. Cell 2005, 121, 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonkin, C.J.; Carret, C.K.; Duraisingh, M.T.; Voss, T.S.; Ralph, S.A.; Hommel, M.; Duffy, M.F.; Da Silva, L.M.; Scherf, A.; Ivens, A.; et al. Sir2 paralogues cooperate to regulate virulence genes and antigenic variation in Plasmodium falciparum. PLoS Biol. 2009, 7, e84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Mu, J.; Zhang, Q.; Ni, T.; Srinivasan, P.; Rayavara, K.; Yang, W.; Turner, L.; Lavstsen, T.; Theander, T.G.; et al. PfSETvs methylation of histone H3K36 represses virulence genes in Plasmodium falciparum. Nature 2013, 499, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Coleman, B.I.; Skillman, K.M.; Jiang, R.H.Y.; Childs, L.M.; Altenhofen, L.M.; Ganter, M.; Leung, Y.; Goldowitz, I.; Kafsack, B.F.C.; Marti, M.; et al. A Plasmodium falciparum histone deacetylase regulates antigenic variation and gametocyte conversion. Cell Host Microbe 2014, 16, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Josling, G.A.; Llinás, M. Sexual development in Plasmodium parasites: Knowing when it’s time to commit. Nat. Rev. Microbiol. 2015, 13, 573–587. [Google Scholar] [CrossRef]
- Josling, G.A.; Williamson, K.C.; Llinás, M. Regulation of Sexual Commitment and Gametocytogenesis in Malaria Parasites. Annu. Rev. Microbiol. 2018, 72, 501–519. [Google Scholar] [CrossRef]
- Abel, S.; Le Roch, K.G. The role of epigenetics and chromatin structure in transcriptional regulation in malaria parasites. Brief. Funct. Genomics 2019, 18, 302–313. [Google Scholar] [CrossRef]
- Brancucci, N.M.B.; Gerdt, J.P.; Wang, C.; de Niz, M.; Philip, N.; Adapa, S.R.; Zhang, M.; Hitz, E.; Niederwieser, I.; Boltryk, S.D.; et al. Lysophosphatidylcholine Regulates Sexual Stage Differentiation in the Human Malaria Parasite Plasmodium falciparum. Cell 2017, 171, 1532–1544.e15. [Google Scholar] [CrossRef] [Green Version]
- Wein, S.; Ghezal, S.; Buré, C.; Maynadier, M.; Périgaud, C.; Vial, H.J.; Lefebvre-Tournier, I.; Wengelnik, K.; Cerdan, R. Contribution of the precursors and interplay of the pathways in the phospholipid metabolism of the malaria parasite. J. Lipid Res. 2018, 59, 1461–1471. [Google Scholar] [CrossRef] [Green Version]
- Flammersfeld, A.; Panyot, A.; Yamaryo-Botté, Y.; Aurass, P.; Przyborski, J.M.; Flieger, A.; Botté, C.; Pradel, G. A patatin-like phospholipase functions during gametocyte induction in the malaria parasite Plasmodium falciparum. Cell. Microbiol. 2020, 22, e13146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaji, S.; Babu, M.M.; Iyer, L.M.; Aravind, L. Discovery of the principal specific transcription factors of Apicomplexa and their implication for the evolution of the AP2-integrase DNA binding domains. Nucleic Acids Res. 2005, 33, 3994–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flueck, C.; Bartfai, R.; Volz, J.; Niederwieser, I.; Salcedo-Amaya, A.M.; Alako, B.T.F.; Ehlgen, F.; Ralph, S.A.; Cowman, A.F.; Bozdech, Z.; et al. Plasmodium falciparum heterochromatin protein 1 marks genomic loci linked to phenotypic variation of exported virulence factors. PLoS Pathog. 2009, 5, e1000569. [Google Scholar] [CrossRef] [Green Version]
- Brancucci, N.M.B.; Bertschi, N.L.; Zhu, L.; Niederwieser, I.; Chin, W.H.; Wampfler, R.; Freymond, C.; Rottmann, M.; Felger, I.; Bozdech, Z.; et al. Heterochromatin protein 1 secures survival and transmission of malaria parasites. Cell Host Microbe 2014, 16, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kafsack, B.F.C.; Rovira-Graells, N.; Clark, T.G.; Bancells, C.; Crowley, V.M.; Campino, S.G.; Williams, A.E.; Drought, L.G.; Kwiatkowski, D.P.; Baker, D.A.; et al. A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 2014, 507, 248–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, A.; Hughes, K.R.; Modrzynska, K.K.; Otto, T.D.; Pfander, C.; Dickens, N.J.; Religa, A.A.; Bushell, E.; Graham, A.L.; Cameron, R.; et al. A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 2014, 507, 253–257. [Google Scholar] [CrossRef]
- Filarsky, M.; Fraschka, S.A.; Niederwieser, I.; Brancucci, N.M.B.; Carrington, E.; Carrió, E.; Moes, S.; Jenoe, P.; Bártfai, R.; Voss, T.S. GDV1 induces sexual commitment of malaria parasites by antagonizing HP1-dependent gene silencing. Science 2018, 359, 1259–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usui, M.; Prajapati, S.K.; Ayanful-Torgby, R.; Acquah, F.K.; Cudjoe, E.; Kakaney, C.; Amponsah, J.A.; Obboh, E.K.; Reddy, D.K.; Barbeau, M.C.; et al. Plasmodium falciparum sexual differentiation in malaria patients is associated with host factors and GDV1-dependent genes. Nat. Commun. 2019, 10, 2140. [Google Scholar] [CrossRef] [Green Version]
- Rea, E.; Le Roch, K.G.; Tewari, R. Sex in Plasmodium falciparum: Silence Play between GDV1 and HP1. Trends Parasitol. 2018, 34, 450–452. [Google Scholar] [CrossRef]
- Poran, A.; Nötzel, C.; Aly, O.; Mencia-Trinchant, N.; Harris, C.T.; Guzman, M.L.; Hassane, D.C.; Elemento, O.; Kafsack, B.F.C. Single-cell RNA sequencing reveals a signature of sexual commitment in malaria parasites. Nature 2017, 551, 95–99. [Google Scholar] [CrossRef]
- Yuda, M.; Kaneko, I.; Iwanaga, S.; Murata, Y.; Kato, T. Female-specific gene regulation in malaria parasites by an AP2-family transcription factor. Mol. Microbiol. 2020, 113, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Flammersfeld, A.; Lang, C.; Flieger, A.; Pradel, G. Phospholipases during membrane dynamics in malaria parasites. Int. J. Med. Microbiol. IJMM 2018, 308, 129–141. [Google Scholar] [CrossRef]
- Llinás, M. Less Lipid, More Commitment. Cell 2017, 171, 1474–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.T.; Tong, X.; Campelo, R.; Vanheer, L.N.; Marreiros, I.M.; Nahiyaan, N.; Zuzarte-Luís, V.A.; Deitsch, K.W.; Mota, M.M.; Rhee, K.Y.; et al. Metabolic competition between lipid metabolism and histone methylation regulates sexual differentiation in human malaria parasites. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ponts, N.; Fu, L.; Harris, E.Y.; Zhang, J.; Chung, D.-W.D.; Cervantes, M.C.; Prudhomme, J.; Atanasova-Penichon, V.; Zehraoui, E.; Bunnik, E.M.; et al. Genome-wide mapping of DNA methylation in the human malaria parasite Plasmodium falciparum. Cell Host Microbe 2013, 14, 696–706. [Google Scholar] [CrossRef] [Green Version]
- Kaur, I.; Zeeshan, M.; Saini, E.; Kaushik, A.; Mohmmed, A.; Gupta, D.; Malhotra, P. Widespread occurrence of lysine methylation in Plasmodium falciparum proteins at asexual blood stages. Sci. Rep. 2016, 6, 35432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgarten, S.; Bryant, J.M.; Sinha, A.; Reyser, T.; Preiser, P.R.; Dedon, P.C.; Scherf, A. Transcriptome-wide dynamics of extensive m6A mRNA methylation during Plasmodium falciparum blood-stage development. Nat. Microbiol. 2019, 4, 2246–2259. [Google Scholar] [CrossRef]
- Liu, M.; Guo, G.; Qian, P.; Mu, J.; Lu, B.; He, X.; Fan, Y.; Shang, X.; Yang, G.; Shen, S.; et al. 5-methylcytosine modification by Plasmodium NSUN2 stabilizes mRNA and mediates the development of gametocytes. Proc. Natl. Acad. Sci. USA 2022, 119, e2110713119. [Google Scholar] [CrossRef]
- Thomas, T.; Thomas, T.J. Polyamines in cell growth and cell death: Molecular mechanisms and therapeutic applications. Cell Mol. Life Sci. CMLS 2001, 58, 244–258. [Google Scholar] [CrossRef]
- Ramya, T.N.C.; Surolia, N.; Surolia, A. Polyamine synthesis and salvage pathways in the malaria parasite Plasmodium falciparum. Biochem. Biophys. Res. Commun. 2006, 348, 579–584. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Golenser, J.; Spira, D.T.; Bachrach, U. Polyamine levels and the activity of their biosynthetic enzymes in human erythrocytes infected with the malarial parasite, Plasmodium falciparum. Biochem. J. 1984, 222, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Das Gupta, R.; Krause-Ihle, T.; Bergmann, B.; Müller, I.B.; Khomutov, A.R.; Müller, S.; Walter, R.D.; Lüersen, K. 3-Aminooxy-1-aminopropane and derivatives have an antiproliferative effect on cultured Plasmodium falciparum by decreasing intracellular polyamine concentrations. Antimicrob. Agents Chemother. 2005, 49, 2857–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Bissati, K.; Redel, H.; Ting, L.-M.; Lykins, J.D.; McPhillie, M.J.; Upadhya, R.; Woster, P.M.; Yarlett, N.; Kim, K.; Weiss, L.M. Novel Synthetic Polyamines Have Potent Antimalarial Activities in vitro and in vivo by Decreasing Intracellular Spermidine and Spermine Concentrations. Front. Cell. Infect. Microbiol. 2019, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Aurrecoechea, C.; Brestelli, J.; Brunk, B.P.; Dommer, J.; Fischer, S.; Gajria, B.; Gao, X.; Gingle, A.; Grant, G.; Harb, O.S.; et al. PlasmoDB: A functional genomic database for malaria parasites. Nucleic Acids Res. 2009, 37, D539–D543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, N.; Scholz, S.M.; Moreira, C.K.; Templeton, T.J.; Kuehn, A.; Dude, M.-A.; Pradel, G. Sexual stage adhesion proteins form multi-protein complexes in the malaria parasite Plasmodium falciparum. J. Biol. Chem. 2009, 284, 14537–14546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, K.C.; Keister, D.B.; Muratova, O.; Kaslow, D.C. Recombinant Pfs230, a Plasmodium falciparum gametocyte protein, induces antisera that reduce the infectivity of Plasmodium falciparum to mosquitoes. Mol. Biochem. Parasitol. 1995, 75, 33–42. [Google Scholar] [CrossRef]
- Weißbach, T.; Golzmann, A.; Bennink, S.; Pradel, G.; Julius Ngwa, C. Transcript and protein expression analysis of proteases in the blood stages of Plasmodium falciparum. Exp. Parasitol. 2017, 180, 33–44. [Google Scholar] [CrossRef] [PubMed]
- von Bohl, A.; Kuehn, A.; Simon, N.; Ngongang, V.N.; Spehr, M.; Baumeister, S.; Przyborski, J.M.; Fischer, R.; Pradel, G. A WD40-repeat protein unique to malaria parasites associates with adhesion protein complexes and is crucial for blood stage progeny. Malar. J. 2015, 14, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, C.C.; Glushakova, S.; Scheuermayer, M.; Repnik, U.; Garg, S.; Schaack, D.; Kachman, M.M.; Weißbach, T.; Zimmerberg, J.; Dandekar, T.; et al. Perforin-like protein PPLP2 permeabilizes the red blood cell membrane during egress of Plasmodium falciparum gametocytes. Cell. Microbiol. 2014, 16, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Ngwa, C.J.; Kiesow, M.J.; Papst, O.; Orchard, L.M.; Filarsky, M.; Rosinski, A.N.; Voss, T.S.; Llinás, M.; Pradel, G. Transcriptional Profiling Defines Histone Acetylation as a Regulator of Gene Expression during Human-to-Mosquito Transmission of the Malaria Parasite Plasmodium falciparum. Front. Cell. Infect. Microbiol. 2017, 7, 320. [Google Scholar] [CrossRef] [Green Version]
- Birnbaum, J.; Flemming, S.; Reichard, N.; Soares, A.B.; Mesén-Ramírez, P.; Jonscher, E.; Bergmann, B.; Spielmann, T. A genetic system to study Plasmodium falciparum protein function. Nat. Methods 2017, 14, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Khosh-Naucke, M.; Becker, J.; Mesén-Ramírez, P.; Kiani, P.; Birnbaum, J.; Fröhlke, U.; Jonscher, E.; Schlüter, H.; Spielmann, T. Identification of novel parasitophorous vacuole proteins in P. falciparum parasites using BioID. Int. J. Med. Microbiol. IJMM 2018, 308, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Treeck, M.; Struck, N.S.; Haase, S.; Langer, C.; Herrmann, S.; Healer, J.; Cowman, A.F.; Gilberger, T.W. A conserved region in the EBL proteins is implicated in microneme targeting of the malaria parasite Plasmodium falciparum. J. Biol. Chem. 2006, 281, 31995–32003. [Google Scholar] [CrossRef]
- Peterson, M.G.; Marshall, V.M.; Smythe, J.A.; Crewther, P.E.; Lew, A.; Silva, A.; Anders, R.F.; Kemp, D.J. Integral membrane protein located in the apical complex of Plasmodium falciparum. Mol. Cell. Biol. 1989, 9, 3151–3154. [Google Scholar]
- Pradel, G.; Hayton, K.; Aravind, L.; Iyer, L.M.; Abrahamsen, M.S.; Bonawitz, A.; Mejia, C.; Templeton, T.J. A multidomain adhesion protein family expressed in Plasmodium falciparum is essential for transmission to the mosquito. J. Exp. Med. 2004, 199, 1533–1544. [Google Scholar] [CrossRef]
- Schneweis, S.; Maier, W.A.; Seitz, H.M. Haemolysis of infected erythrocytes—A trigger for formation of Plasmodium falciparum gametocytes? Parasitol. Res. 1991, 77, 458–460. [Google Scholar] [CrossRef]
- Hughes, C.S.; Foehr, S.; Garfield, D.A.; Furlong, E.E.; Steinmetz, L.M.; Krijgsveld, J. Ultrasensitive proteome analysis using paramagnetic bead technology. Mol. Syst. Biol. 2014, 10, 757. [Google Scholar] [CrossRef]
- Sielaff, M.; Kuharev, J.; Bohn, T.; Hahlbrock, J.; Bopp, T.; Tenzer, S.; Distler, U. Evaluation of FASP, SP3, and iST Protocols for Proteomic Sample Preparation in the Low Microgram Range. J. Proteome Res. 2017, 16, 4060–4072. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteomics MCP 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizcaíno, J.A.; Côté, R.G.; Csordas, A.; Dianes, J.A.; Fabregat, A.; Foster, J.M.; Griss, J.; Alpi, E.; Birim, M.; Contell, J.; et al. The PRoteomics IDEntifications (PRIDE) database and associated tools: Status in 2013. Nucleic Acids Res. 2013, 41, D1063–D1069. [Google Scholar] [CrossRef] [PubMed]
- Prommana, P.; Uthaipibull, C.; Wongsombat, C.; Kamchonwongpaisan, S.; Yuthavong, Y.; Knuepfer, E.; Holder, A.A.; Shaw, P.J. Inducible knockdown of Plasmodium gene expression using the glmS ribozyme. PLoS ONE 2013, 8, e73783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, K.J.; In Kim, D.; Raida, M.; Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Sharma, B.; Gupta, A.; Dhar, S.K. Identification of a novel trafficking pathway exporting a replication protein, Orc2 to nucleus via classical secretory pathway in Plasmodium falciparum. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Fan, Q.; Cui, L.; Miao, J. Histone lysine methyltransferases and demethylases in Plasmodium falciparum. Int. J. Parasitol. 2008, 38, 1083–1097. [Google Scholar] [CrossRef] [Green Version]
- Volz, J.; Carvalho, T.G.; Ralph, S.A.; Gilson, P.; Thompson, J.; Tonkin, C.J.; Langer, C.; Crabb, B.S.; Cowman, A.F. Potential epigenetic regulatory proteins localise to distinct nuclear sub-compartments in Plasmodium falciparum. Int. J. Parasitol. 2010, 40, 109–121. [Google Scholar] [CrossRef]
- Fan, Q.; Miao, J.; Cui, L.; Cui, L. Characterization of PRMT1 from Plasmodium falciparum. Biochem. J. 2009, 421, 107–118. [Google Scholar] [CrossRef]
- Reytor, E.; Pérez-Miguelsanz, J.; Alvarez, L.; Pérez-Sala, D.; Pajares, M.A. Conformational signals in the C-terminal domain of methionine adenosyltransferase I/III determine its nucleocytoplasmic distribution. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 3347–3360. [Google Scholar] [CrossRef] [Green Version]
- Coetzee, N.; Sidoli, S.; van Biljon, R.; Painter, H.; Llinás, M.; Garcia, B.A.; Birkholtz, L.-M. Quantitative chromatin proteomics reveals a dynamic histone post-translational modification landscape that defines asexual and sexual Plasmodium falciparum parasites. Sci. Rep. 2017, 7, 607. [Google Scholar] [CrossRef]
- Ngwa, C.J.; Kiesow, M.J.; Orchard, L.M.; Farrukh, A.; Llinás, M.; Pradel, G. The G9a Histone Methyltransferase Inhibitor BIX-01294 Modulates Gene Expression during Plasmodium falciparum Gametocyte Development and Transmission. Int. J. Mol. Sci. 2019, 20, 5087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connacher, J.; von Grüning, H.; Birkholtz, L. Histone Modification Landscapes as a Roadmap for Malaria Parasite Development. Front. Cell Dev. Biol. 2022, 10, 848797. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.L.; de Silva, E.K.; Olszewski, K.L.; Elemento, O.; Llinás, M. Identification and genome-wide prediction of DNA binding specificities for the ApiAP2 family of regulators from the malaria parasite. PLoS Pathog. 2010, 6, e1001165. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.M.; Macpherson, C.R.; Claes, A.; Scheidig-Benatar, C.; Sakamoto, H.; Yam, X.Y.; Preiser, P.; Goel, S.; Wahlgren, M.; Sismeiro, O.; et al. An ApiAP2 member regulates expression of clonally variant genes of the human malaria parasite Plasmodium falciparum. Sci. Rep. 2017, 7, 14042. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, S.; Kaneko, I.; Kato, T.; Yuda, M. Identification of an AP2-family protein that is critical for malaria liver stage development. PLoS ONE 2012, 7, e47557. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Cui, H.; Jiang, Y.; Yang, Z.; Wang, X.; Gao, H.; Liu, C.; Zhang, S.; Su, X.; et al. Systematic CRISPR-Cas9-Mediated Modifications of Plasmodium yoelii ApiAP2 Genes Reveal Functional Insights into Parasite Development. mBio 2017, 8, e01986-17. [Google Scholar] [CrossRef] [Green Version]
- Katoh, Y.; Ikura, T.; Hoshikawa, Y.; Tashiro, S.; Ito, T.; Ohta, M.; Kera, Y.; Noda, T.; Igarashi, K. Methionine adenosyltransferase II serves as a transcriptional corepressor of Maf oncoprotein. Mol. Cell 2011, 41, 554–566. [Google Scholar] [CrossRef]
- Portillo, F.; Vázquez, J.; Pajares, M.A. Protein-protein interactions involving enzymes of the mammalian methionine and homocysteine metabolism. Biochimie 2020, 173, 33–47. [Google Scholar] [CrossRef]
- Kors, S.; Geijtenbeek, K.; Reits, E.; Schipper-Krom, S. Regulation of Proteasome Activity by (Post-)transcriptional Mechanisms. Front. Mol. Biosci. 2019, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Orozco, J.M.; Saxton, R.A.; Condon, K.J.; Liu, G.Y.; Krawczyk, P.A.; Scaria, S.M.; Harper, J.W.; Gygi, S.P.; Sabatini, D.M. SAMTOR is an S-adenosylmethionine sensor for the mTORC1 pathway. Science 2017, 358, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Serfontein, J.; Nisbet, R.E.R.; Howe, C.J.; de Vries, P.J. Evolution of the TSC1/TSC2-TOR signaling pathway. Sci. Signal. 2010, 3, ra49. [Google Scholar] [CrossRef]
- van Dam, T.J.P.; Zwartkruis, F.J.T.; Bos, J.L.; Snel, B. Evolution of the TOR pathway. J. Mol. Evol. 2011, 73, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, K.J.; Jacobs-Lorena, M. Plasmodium falciparum Maf1 Confers Survival upon Amino Acid Starvation. mBio 2017, 8, e02317-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennink, S.; Pradel, G. The molecular machinery of translational control in malaria parasites. Mol. Microbiol. 2019, 112, 1658–1673. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, K.; Arai, T.; Suzuki, T. Depletion of S-adenosylmethionine impacts on ribosome biogenesis through hypomodification of a single rRNA methylation. Nucleic Acids Res. 2019, 47, 4226–4239. [Google Scholar] [CrossRef]
- Yan, F.; LaMarre, J.M.; Röhrich, R.; Wiesner, J.; Jomaa, H.; Mankin, A.S.; Fujimori, D.G. RlmN and Cfr are radical SAM enzymes involved in methylation of ribosomal RNA. J. Am. Chem. Soc. 2010, 132, 3953–3964. [Google Scholar] [CrossRef] [Green Version]
- Stojković, V.; Fujimori, D.G. Radical SAM-Mediated Methylation of Ribosomal RNA. Methods Enzymol. 2015, 560, 355–376. [Google Scholar]
- Hart, R.J.; Ghaffar, A.; Abdalal, S.; Perrin, B.; Aly, A.S.I. Plasmodium AdoMetDC/ODC bifunctional enzyme is essential for male sexual stage development and mosquito transmission. Biol. Open 2016, 5, 1022–1029. [Google Scholar] [CrossRef] [Green Version]
- Assaraf, Y.G.; Golenser, J.; Spira, D.T.; Messer, G.; Bachrach, U. Cytostatic effect of DL-alpha-difluoromethylornithine against Plasmodium falciparum and its reversal by diamines and spermidine. Parasitol. Res. 1987, 73, 313–318. [Google Scholar] [CrossRef]
- Bitonti, A.J.; McCann, P.P.; Sjoerdsma, A. Plasmodium falciparum and Plasmodium berghei: Effects of ornithine decarboxylase inhibitors on erythrocytic schizogony. Exp. Parasitol. 1987, 64, 237–243. [Google Scholar] [CrossRef]
- Wright, P.S.; Byers, T.L.; Cross-Doersen, D.E.; McCann, P.P.; Bitonti, A.J. Irreversible inhibition of S-adenosylmethionine decarboxylase in Plasmodium falciparum-infected erythrocytes: Growth inhibition in vitro. Biochem. Pharmacol. 1991, 41, 1713–1718. [Google Scholar] [CrossRef]
- van Brummelen, A.C.; Olszewski, K.L.; Wilinski, D.; Llinás, M.; Louw, A.I.; Birkholtz, L.-M. Co-inhibition of Plasmodium falciparum S-adenosylmethionine decarboxylase/ornithine decarboxylase reveals perturbation-specific compensatory mechanisms by transcriptome, proteome, and metabolome analyses. J. Biol. Chem. 2009, 284, 4635–4646. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musabyimana, J.P.; Distler, U.; Sassmannshausen, J.; Berks, C.; Manti, J.; Bennink, S.; Blaschke, L.; Burda, P.-C.; Flammersfeld, A.; Tenzer, S.; et al. Plasmodium falciparum S-Adenosylmethionine Synthetase Is Essential for Parasite Survival through a Complex Interaction Network with Cytoplasmic and Nuclear Proteins. Microorganisms 2022, 10, 1419. https://doi.org/10.3390/microorganisms10071419
Musabyimana JP, Distler U, Sassmannshausen J, Berks C, Manti J, Bennink S, Blaschke L, Burda P-C, Flammersfeld A, Tenzer S, et al. Plasmodium falciparum S-Adenosylmethionine Synthetase Is Essential for Parasite Survival through a Complex Interaction Network with Cytoplasmic and Nuclear Proteins. Microorganisms. 2022; 10(7):1419. https://doi.org/10.3390/microorganisms10071419
Chicago/Turabian StyleMusabyimana, Jean Pierre, Ute Distler, Juliane Sassmannshausen, Christina Berks, Janice Manti, Sandra Bennink, Lea Blaschke, Paul-Christian Burda, Ansgar Flammersfeld, Stefan Tenzer, and et al. 2022. "Plasmodium falciparum S-Adenosylmethionine Synthetase Is Essential for Parasite Survival through a Complex Interaction Network with Cytoplasmic and Nuclear Proteins" Microorganisms 10, no. 7: 1419. https://doi.org/10.3390/microorganisms10071419