
Warming-Induced Labile Carbon Change Soil Organic Carbon Mineralization and Microbial Abundance in a Northern Peatland

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Soil Sampling
2.2. Laboratory Incubation and CO2 Analysis
2.3. Fungal and Bacterial Abundance Analysis
2.4. Enzyme Activity Analysis
2.5. Soil MBC, DOC, NH4+-N, and NO3−-N Contents Analysis
2.6. Total Carbon (TC) and Total Nitrogen (TN) Contents Analysis
2.7. Calculations and Statistical Analysis
2.8. Statistical Analyses
3. Results
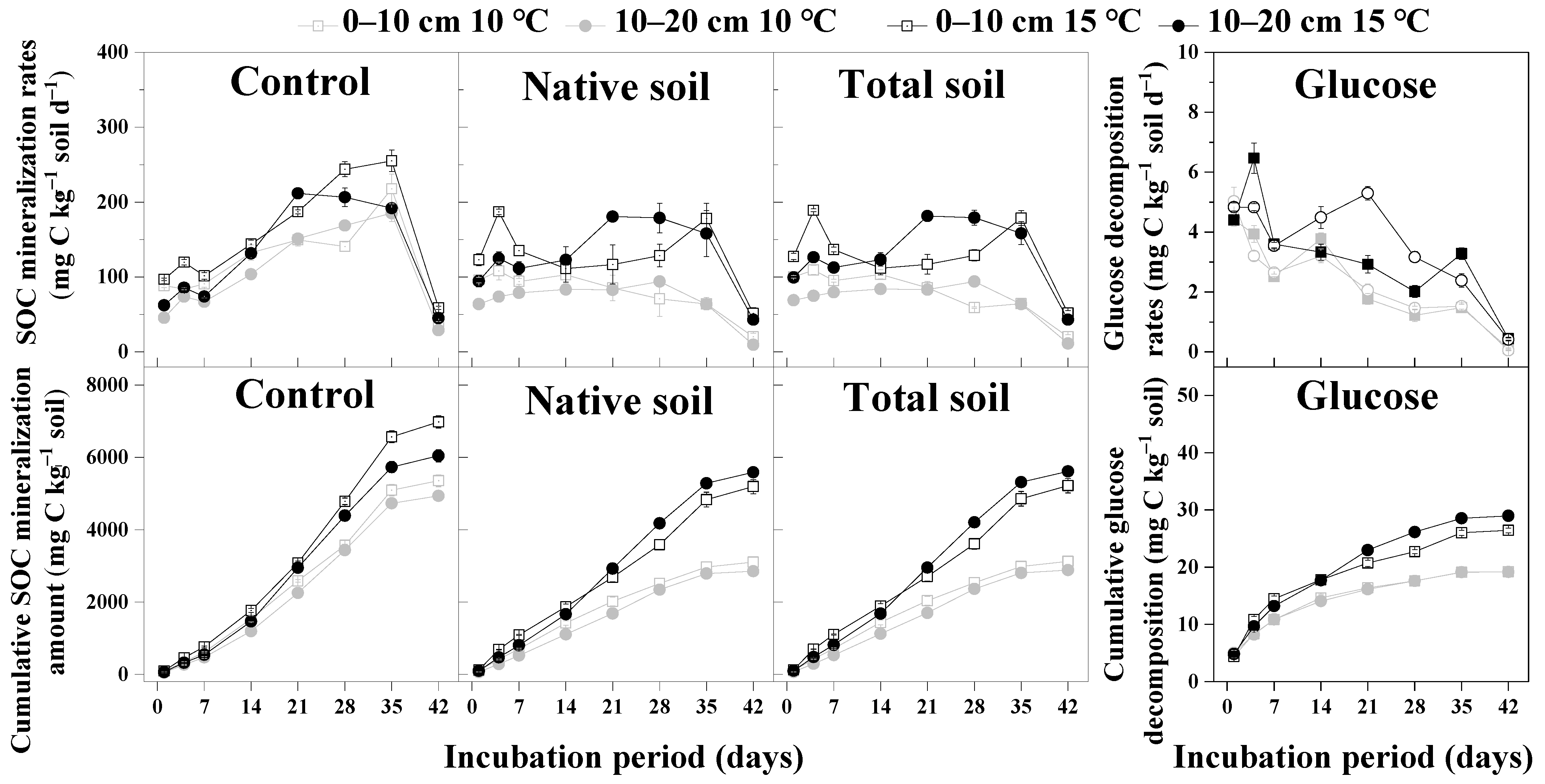
3.1. The Effects of Warming and Glucose Addition on SOC Mineralization
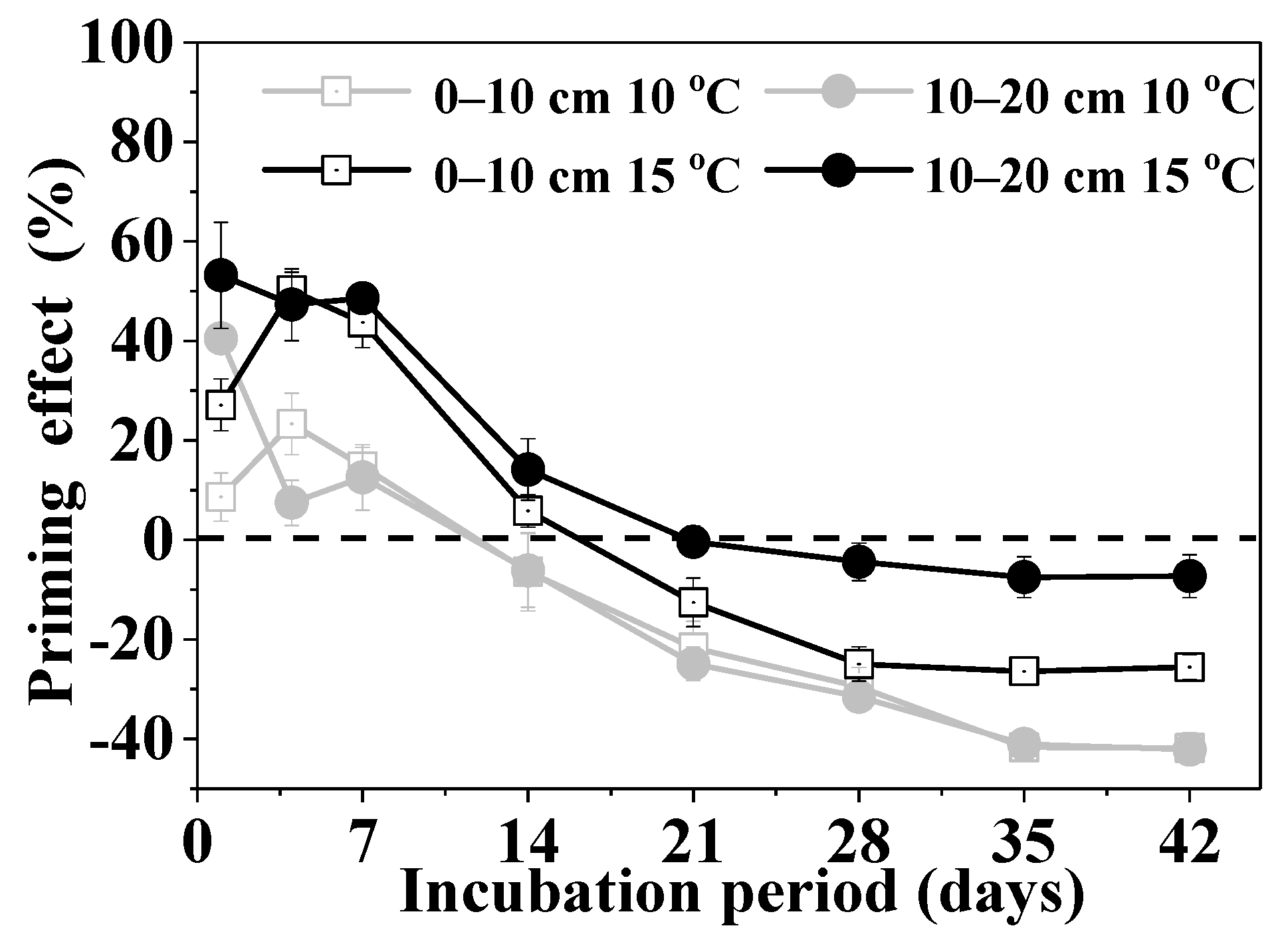
3.2. Priming Effect
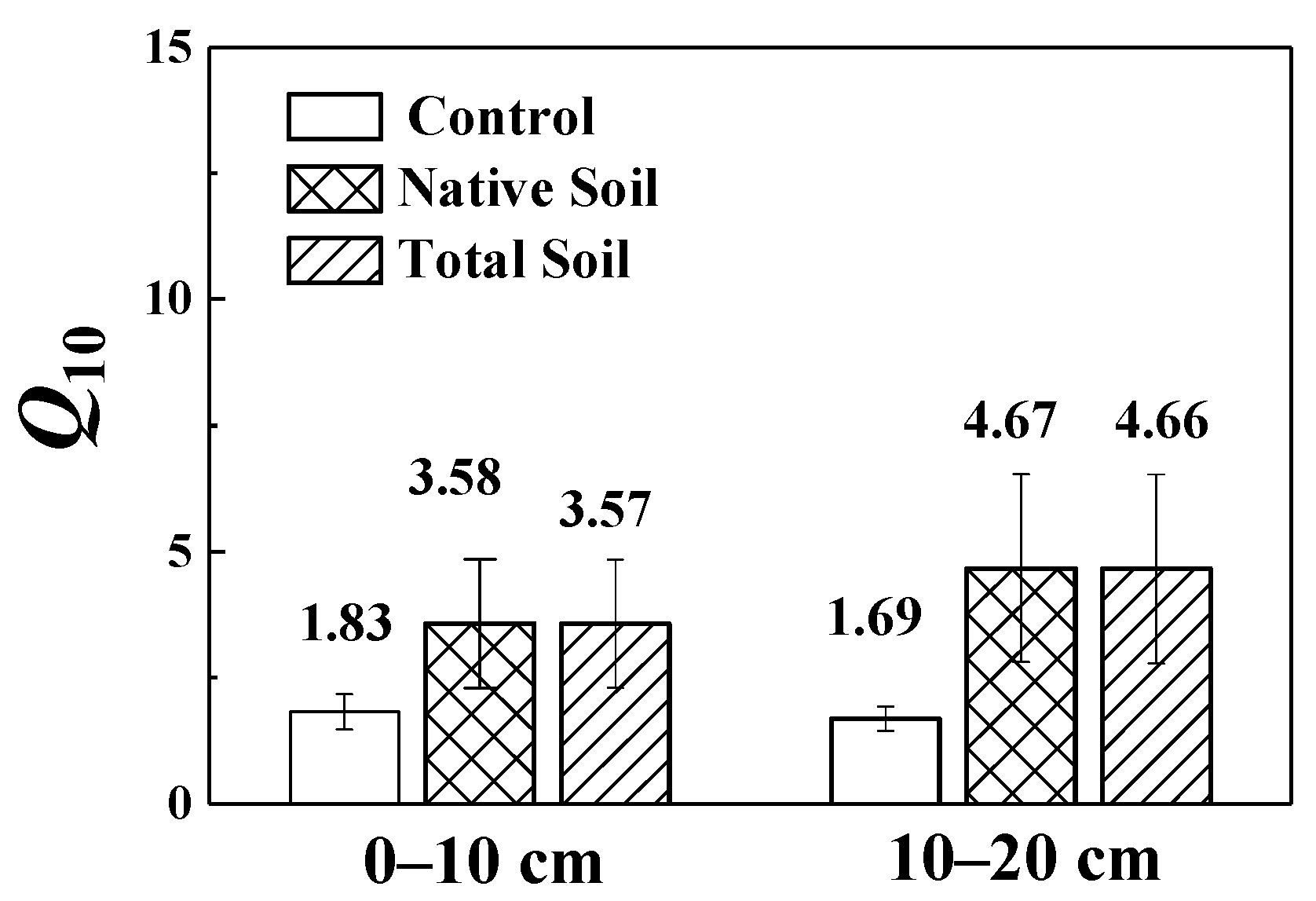
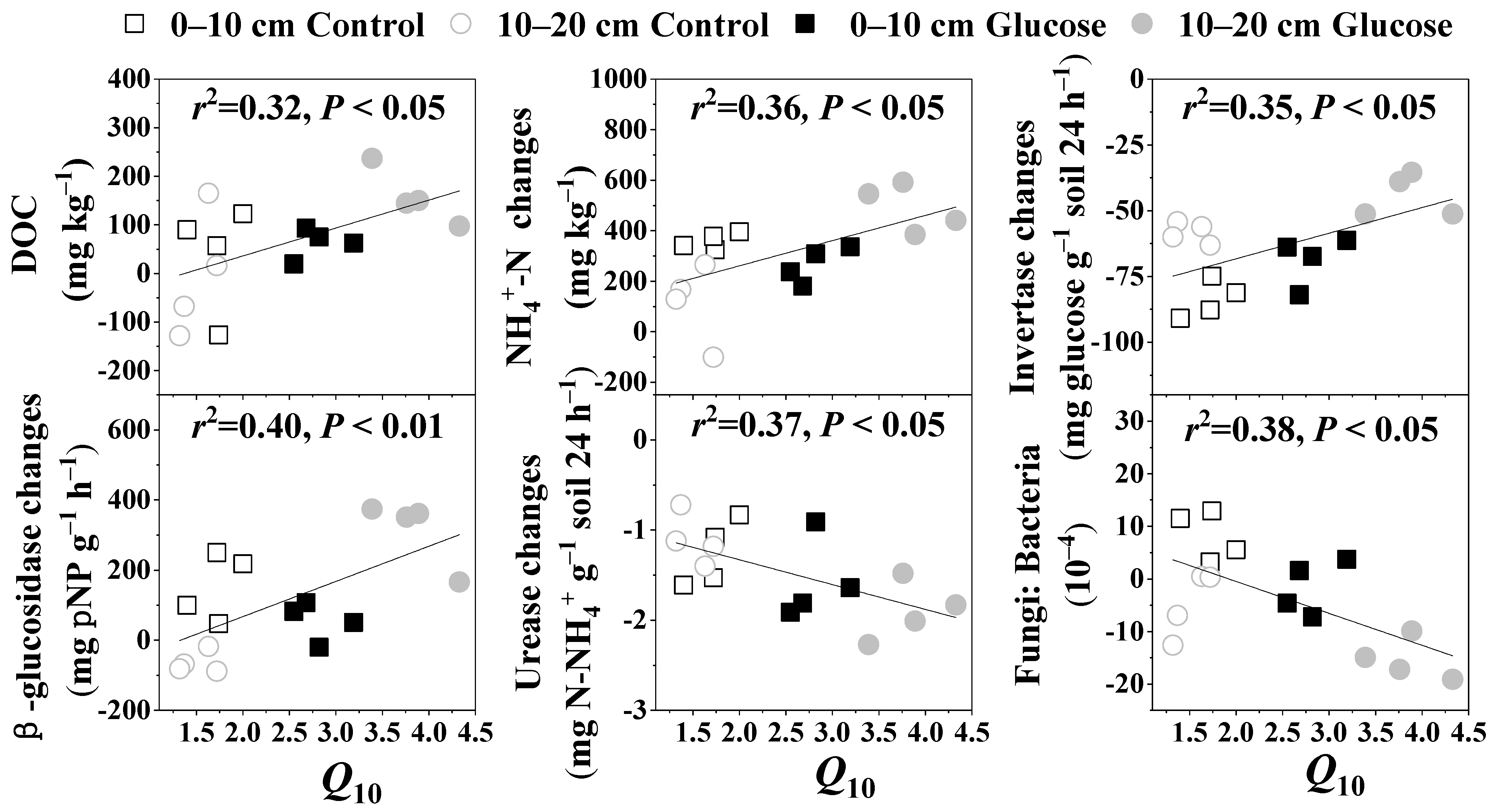
3.3. Temperature Sensitivity of SOC Mineralization
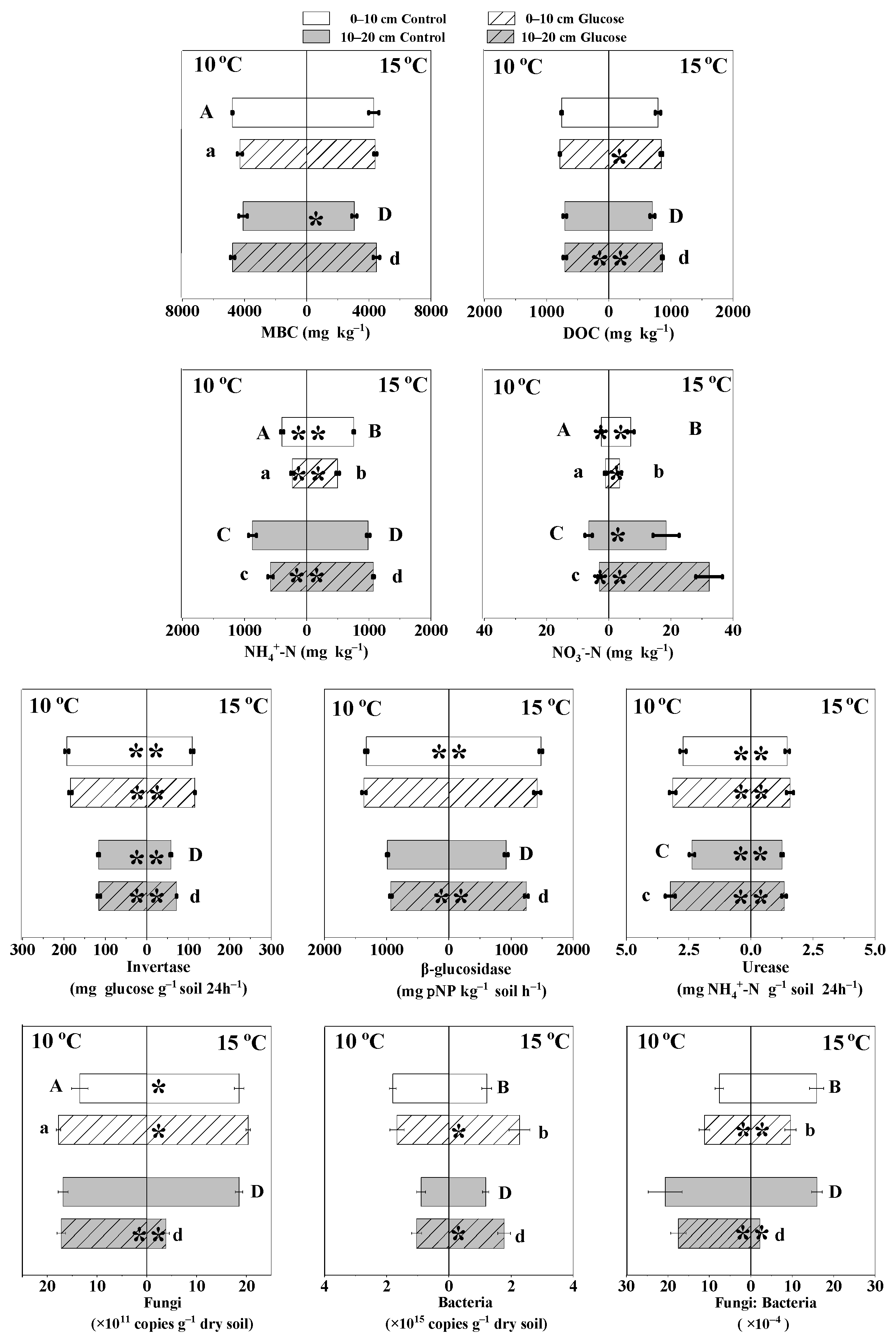
3.4. The Effects of Warming and Glucose Addition on Fungal and Bacterial Abundances
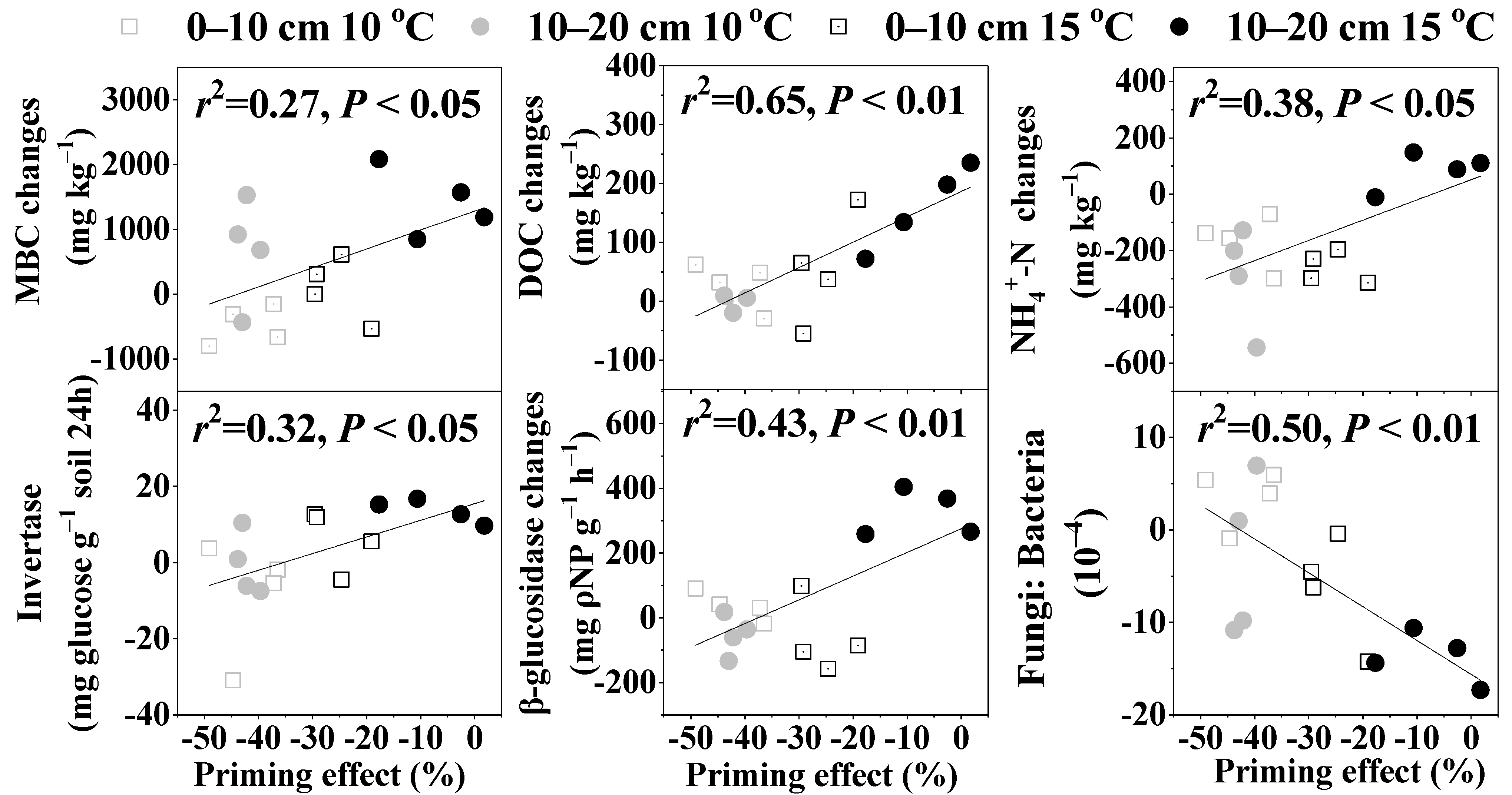
3.5. The Effects of Warming and Glucose Addition on MBC and DOC Contents
3.6. The Effects of Warming and Glucose Addition on NH4+-N and NO3−-N Contents
3.7. The Effects of Warming and Glucose Addition on Invertase, β-glucosidase, and Urease Activities
4. Discussion
4.1. Response of SOC Mineralization to Warming and Its Temperature Sensitivity
4.2. Response of SOC Mineralization to Glucose Addition and Its Microbial Mechanism
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Johansson, T.; Malmer, N.; Crill, P.M.; Friborg, T.; Akerman, J.H.; Mastepanov, M.; Christensen, T.R. Decadal vegetation changes in a northern peatland, greenhouse gas fluxes and net radiative forcing. Glob. Change Biol. 2006, 12, 2352–2369. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Bockheim, J.; Canadell, J.G.; Euskirchen, E.; Field, C.B.; Goryachkin, S.V.; Hagemann, S.; Kuhry, P.; Lafleur, P.M.; Lee, H.; et al. Vulnerability of permafrost carbon to climate change: Implications for the global carbon cycle. Bioscience 2008, 58, 701–714. [Google Scholar] [CrossRef]
- Gorham, E. Northern peatlands-role in the carbon-cycle and probable responses to climatic warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef]
- Alexandrov, G.A.; Brovkin, V.; Kleinen, T.; Yu, Z. The capacity of northern peatlands for long-term carbon sequestration. Biogeosciences 2020, 17, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Liu, Q.Y.; Xu, X.L.; Wang, H.M.; Blagodatskaya, E.; Kuzyakov, Y. Dominant extracellular enzymes in priming of SOM decomposition depend on temperature. Geoderma 2019, 343, 187–195. [Google Scholar] [CrossRef]
- Wu, H.T.; Lu, M.Z.; Lu, X.G.; Guan, Q.; He, X.H. Interactions between earthworms and mesofauna has no significant effect on emissions of CO2 and N2O from soil. Soil Biol. Biochem. 2015, 88, 294–297. [Google Scholar] [CrossRef]
- Wu, H.T.; Lu, X.G.; Wu, D.H.; Song, L.H.; Yan, X.M.; Liu, J. Ant mounds alter spatial and temporal patterns of CO2, CH4 and N2O emissions from a marsh soil. Soil Biol. Biochem. 2013, 57, 884–891. [Google Scholar] [CrossRef]
- Song, Y.Y.; Song, C.C.; Hou, A.X.; Ren, J.S.; Wang, X.W.; Cui, Q.; Wang, M.Q. Effects of temperature and root additions on soil carbon and nitrogen mineralization in a predominantly permafrost peatland. Catena 2018, 165, 381–389. [Google Scholar] [CrossRef]
- Chen, L.Y.; Liu, L.; Qin, S.Q.; Yang, G.B.; Fang, K.; Zhu, B.; Kuzyakov, Y.; Chen, P.D.; Xu, Y.P.; Yang, Y.H. Regulation of priming effect by soil organic matter stability over a broad geographic scale. Nat. Commun. 2019, 10, 5112. [Google Scholar] [CrossRef] [Green Version]
- Perveen, N.; Barot, S.; Maire, V.; Cotrufo, M.F.; Shahzad, T.; Blagodatskaya, E.; Stewart, C.E.; Ding, W.X.; Siddiq, M.R.; Dimassi, B.; et al. Universality of priming effect: An analysis using thirty five soils with contrasted properties sampled from five continents. Soil Biol. Biochem. 2019, 134, 162–171. [Google Scholar] [CrossRef]
- Mau, R.L.; Dijkstra, P.; Schwartz, E.; Koch, B.J.; Hungate, B.A. Warming induced changes in soil carbon and nitrogen influence priming responses in four ecosystems. Appl. Soil Ecol. 2018, 124, 110–116. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Hill, P.W.; Jones, D.L. Root exudate components change litter decomposition in a simulated rhizosphere depending on temperature. Plant Soil 2007, 290, 293–305. [Google Scholar] [CrossRef]
- Liu, Y.; Shahbaz, M.; Ge, T.; Zhu, Z.; Liu, S.; Chen, L.; Wu, X.; Deng, Y.; Lu, S.; Wu, J. Effects of root exudate stoichiometry on CO2 emission from paddy soil. Eur. J. Soil Biol. 2020, 101, 103247. [Google Scholar] [CrossRef]
- Renella, G.; Egamberdiyeva, D.; Landi, L.; Mench, M.; Nannipieri, P. Microbial activity and hydrolase activities during decomposition of root exudates released by an artifificial root surface in Cd-contaminated soils. Soil Biol. Biochem. 2006, 38, 702–708. [Google Scholar] [CrossRef]
- Mcgill, B.J. Trees are rarely most abundant where they grow best. J. Plant Ecol. 2012, 5, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Rusinova-Videva, S.; Pavlova, K.; Georgieva, K. Effect of different carbon sources on biosynthesis of exopolysaccharide from Antarctic Strain Cryptococcus Laurentii AL62. Biotechnol. Biotechnol. Equip. 2011, 25, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Zhu, Z.; Qi, Y.; Zou, D.; Zhang, G.; Zeng, X.; Ge, T.; Wu, J.; Xiao, Z. Effects of different stoichiometric ratios on mineralisation of root exudates and its priming effect in paddy soil. Sci. Total Environ. 2020, 743, 140808. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Huang, Y.; Fu, S.; Wang, J.; Ming, A.; Li, X.; Yao, M.; Li, H. Tree species mixture inhibits soil organic carbon mineralization accompanied by decreased r-selected bacteria. Plant Soil 2018, 431, 203–216. [Google Scholar] [CrossRef]
- Sun, Z.L.; Liu, S.G.; Zhang, T.A.; Zhao, X.C.; Chen, S.; Wang, Q.K. Priming of soil organic carbon decomposition induced by exogenous organic carbon input: A meta-analysis. Plant Soil 2019, 443, 463–471. [Google Scholar] [CrossRef]
- Cheng, W.X. Rhizosphere feedbacks in elevated CO2. Tree Physiol. 1999, 19, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Craine, J.M.; Morrow, C.; Fierer, N. Microbial nitrogen limitation increases decomposition. Ecology 2007, 88, 2105–2113. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar] [CrossRef]
- Hobbie, J.E.; Hobbie, E.A. Microbes in nature are limited by carbon and energy: The starving-survival lifestyle in soil and consequences for estimating microbial rates. Front. Microbiol. 2013, 4, 324. [Google Scholar] [CrossRef] [Green Version]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- De Graaff, M.A.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Dungait, J.A.J.; Kemmitt, S.J.; Michallon, L.; Guo, S.L.; Wen, Q.; Brookes, P.C.; Evershed, R.P. The variable response of soil microorganisms to trace concentrations of low molecular weight organic substrates of increasing complexity. Soil Biol. Biochem. 2013, 64, 57–64. [Google Scholar] [CrossRef]
- Qiu, Q.Y.; Wu, L.F.; Ouyang, Z.; Li, B.B.; Xu, Y.Y. Different effects of plant-derived dissolved organic matter (DOM) and urea on the priming of soil organic carbon. Environ. Sci.-Process. Impacts 2016, 18, 330–341. [Google Scholar] [CrossRef]
- Naylor, D.; McClure, R.; Jansson, J. Trends in Microbial Community Composition and Function by Soil Depth. Microorganisms 2022, 10, 540. [Google Scholar] [CrossRef]
- Keiser, A.D.; Smith, M.; Bell, S.; Hofmockel, K.S. Peatland microbial community response to altered climate tempered by nutrient availability. Soil Biol. Biochem. 2019, 137, 107561. [Google Scholar] [CrossRef]
- Gromova, M.S.; Matvienko, A.I.; Makarov, M.I.; Cheng, C.H.; Menyailo, O.V. Temperature Sensitivity (Q(10)) of Soil Basal Respiration as a Function of Available Carbon Substrate, Temperature, and Moisture. Eurasian Soil Sci. 2020, 53, 377–382. [Google Scholar] [CrossRef]
- Li, X.J.; Xie, J.S.; Zhang, Q.F.; Lyu, M.K.; Xiong, X.L.; Liu, X.F.; Lin, T.C.; Yang, Y.S. Substrate availability and soil microbes drive temperature sensitivity of soil organic carbon mineralization to warming along an elevation gradient in subtropical Asia. Geoderma 2020, 364, 114198. [Google Scholar] [CrossRef]
- Xu, X.H.; Yang, B.S.; Wang, H.; Cao, Y.A.; Li, K.; Gao, S.W. Temperature sensitivity of soil heterotrophic respiration is altered by carbon substrate along the development of Quercus Mongolica forest in northeast China. Appl. Soil Ecol. 2019, 133, 52–61. [Google Scholar] [CrossRef]
- Guan, Z.J.; Luo, Q.; Chen, X.; Feng, X.W.; Tang, Z.X.; Wei, W.; Zheng, Y.R. Saline soil enzyme activities of four plant communities in Sangong River basin of Xinjiang, China. J. Arid Land 2014, 6, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Veres, Z.; Kotroczo, Z.; Fekete, I.; Toth, J.A.; Lajtha, K.; Townsend, K.; Tothmeresz, B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Appl. Soil Ecol. 2015, 92, 18–23. [Google Scholar] [CrossRef]
- Feng, W.T.; Liang, J.Y.; Hale, L.E.; Jung, C.G.; Chen, J.; Zhou, J.Z.; Xu, M.G.; Yuan, M.T.; Wu, L.Y.; Bracho, R.; et al. Enhanced decomposition of stable soil organic carbon and microbial catabolic potentials by long-term field warming. Glob. Change Biol. 2017, 23, 4765–4776. [Google Scholar] [CrossRef]
- Chen, J.; Elsgaard, L.; van Groenigen, K.J.; Olesen, J.E.; Liang, Z.; Jiang, Y.; Lærke, P.E.; Zhang, Y.F.; Luo, Y.Q.; Hungate, B.A.; et al. Soil carbon loss with warming: New evidence from carbon-degrading enzymes. Glob. Change Biol. 2020, 26, 1944–1952. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 11th ed.; USDA—Natural Resources Conservation Service: Washington, DC, USA, 2010. [Google Scholar]
- Song, Y.Y.; Song, C.C.; Ren, J.S.; Tan, W.W.; Jin, S.F.; Jiang, L. Influence of nitrogen additions on litter decomposition, nutrient dynamics, and enzymatic activity of two plant species in a peatland in Northeast China. Sci. Total Environ. 2018, 625, 640–646. [Google Scholar] [CrossRef]
- Wang, J.; Song, C.; Hou, A.; Miao, Y.; Yang, G.; Zhang, J. Effects of freezing-thawing cycle on peatland active organic carbon fractions and enzyme activities in the Da Xing’anling Mountains, Northeast China. Environ. Earth Sci. 2014, 72, 1853–1860. [Google Scholar] [CrossRef]
- Yin, H.; Wheeler, E.; Phillips, R.P. Root-induced changes in nutrient cycling in forests depend on exudation rates. Soil Biol. Biochem. 2014, 78, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.P.; Ni, Y.Y.; Liu, X.J.; Chu, H.Y. Microbes changed their carbon use strategy to regulate the priming effect in an 11-year nitrogen addition experiment in grassland. Sci. Total Environ. 2020, 727, 138645. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Song, C.C.; Ren, J.S.; Ma, X.Y.; Tan, W.W.; Wang, X.W.; Gao, J.L.; Hou, A.X. Short-term response of the soil microbial abundances and enzyme activities to experimental warming in a boreal peatland in Northeast China. Sustainability 2019, 11, 590. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Enzymes. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties; Soil Science Society of America: Madison, WI, USA, 1994. [Google Scholar]
- Yao, H.Y. Soil Microbial Ecology and Its Experimental Techniques; Science Press: Beijing, China, 2006. [Google Scholar]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Ghani, A.; Dexter, M.; Perrott, K.W. Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilisation, grazing and cultivation. Soil Biol. Biochem. 2003, 35, 1231–1243. [Google Scholar] [CrossRef]
- Jin, T.; Shimizu, M.; Marutani, S.; Desyatkin, A.R.; Iizuka, N.; Hata, H.; Hatano, R. Effect of chemical fertilizer and manure application on N2O emission from reed canary grassland in Hokkaido, Japan. Soil Sci. Plant Nutr. 2010, 56, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Leifeld, J.; Fuhrer, J. The temperature response of CO2 production from bulk soils and soil fractions is related to soil organic matter quality. Biogeochemistry 2005, 75, 433–453. [Google Scholar] [CrossRef]
- Zhang, X.W.; Han, X.Z.; Yu, W.T.; Wang, P.; Cheng, W.X. Priming effects on labile and stable soil organic carbon decomposition: Pulse dynamics over two years. PLoS ONE 2017, 12, e0184978. [Google Scholar] [CrossRef] [Green Version]
- Thiessen, S.; Gleixner, G.; Wutzler, T.; Reichstein, M. Both priming and temperature sensitivity of soil organic matter decomposition depend on microbial biomass—An incubation study. Soil Biol. Biochem. 2013, 57, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Zhang, T.; Guo, R.; Cao, H.; Shi, L.; Guo, J.; Sun, W. Response of soil enzyme activity to warming and nitrogen addition in a meadow steppe. Soil Res. 2015, 53, 242–252. [Google Scholar] [CrossRef]
- Liu, Y.; He, N.P.; Xu, L.; Tian, J.; Gao, Y.; Zheng, S.; Wang, Q.; Wen, X.F.; Xu, X.L.; Yakov, K. A new incubation and measurement approach to estimate the temperature response of soil organic matter decomposition. Soil Biol. Biochem. 2019, 138, 107596. [Google Scholar] [CrossRef]
- Fissore, C.; Giardina, C.P.; Kolka, R.K. Reduced substrate supply limits the temperature response of soil organic carbon decomposition. Soil Biol. Biochem. 2013, 67, 306–311. [Google Scholar] [CrossRef]
- Gershenson, A.; Bader, N.E.; Cheng, W.X. Effects of substrate availability on the temperature sensitivity of soil organic matter decomposition. Glob. Change Biol. 2009, 15, 176–183. [Google Scholar] [CrossRef]
- Fang, Y.Y.; Singh, B.P.; Matta, P.; Cowie, A.L.; Van Zwieten, L. Temperature sensitivity and priming of organic matter with different stabilities in a Vertisol with aged biochar. Soil Biol. Biochem. 2017, 115, 346–356. [Google Scholar] [CrossRef]
- Lefevre, R.; Barre, P.; Moyano, F.E.; Christensen, B.T.; Bardoux, G.; Eglin, T.; Girardin, C.; Houot, S.; Katterer, T.; van Oort, F.; et al. Higher temperature sensitivity for stable than for labile soil organic carbon—Evidence from incubations of long-term bare fallow soils. Glob. Change Biol. 2014, 20, 633–640. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Y.; Ruan, H.H.; Luo, Y.Q.; Wang, J.S. Temperature sensitivity increases with soil organic carbon recalcitrance along an elevational gradient in the Wuyi Mountains, China. Soil Biol. Biochem. 2010, 42, 1811–1815. [Google Scholar] [CrossRef]
- Wetterstedt, J.A.M.; Persson, T.; Agren, G.I. Temperature sensitivity and substrate quality in soil organic matter decomposition: Results of an incubation study with three substrates. Glob. Change Biol. 2010, 16, 1806–1819. [Google Scholar] [CrossRef]
- Creamer, C.A.; de Menezes, A.B.; Krull, E.S.; Sanderman, J.; Newton-Walters, R.; Farrell, M. Microbial community structure mediates response of soil C decomposition to litter addition and warming. Soil Biol. Biochem. 2015, 80, 175–188. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.Y.; Liu, C.; Song, C.C.; Wang, X.W.; Wang, L.L. Linking soil organic carbon mineralization with soil microbial and substrate properties under warming in permafrost peatlands of northeastern china. Catena 2021, 203, 105348. [Google Scholar] [CrossRef]
- Bradford, M.A.; Keiser, A.D.; Davies, C.A.; Mersmann, C.A.; Strickland, M.S. Empirical evidence that soil carbon formation from plant inputs is positively related to microbial growth. Biogeochemistry 2013, 113, 271–281. [Google Scholar] [CrossRef]
- Hill, P.W.; Farrar, J.F.; Jones, D.L. Decoupling of microbial glucose uptake and mineralization in soil. Soil Biol. Biochem. 2008, 40, 616–624. [Google Scholar] [CrossRef]
- Hopkins, D.W.; Chudek, J.A.; Webster, E.A.; Barraclough, D. Following the decomposition of ryegrass labelled with C-13 and N-15 in soil by solid-state nuclear magnetic resonance spectroscopy. Eur. J. Soil Sci. 1997, 48, 623–631. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, W.; Xiao, J.; Zhang, Z.; Qiao, M.; Liu, Q.; Yin, H. Exudate components exert different influences on microbially mediated C losses in simulated rhizosphere soils of a spruce plantation. Plant Soil 2017, 419, 127–140. [Google Scholar] [CrossRef]
- Kirkby, C.A.; Richardson, A.E.; Wade, L.J.; Passioura, J.B.; Batten, G.D.; Blanchard, C.; Kirkegaard, J.A. Nutrient availability limits carbon sequestration in arable soils. Soil Biol. Biochem. 2014, 68, 402–409. [Google Scholar] [CrossRef]
- Qiu, Q.Y.; Yao, K.L.; Liu, Z.Q.; Xu, W.B.; Liu, H.X.; Hu, Y.L. Effects of labile organic carbon input on the priming effect along an ecological restoration gradient. Acta Ecol. Sin. 2019, 39, 4855–4864. [Google Scholar]
- Xiao, C.W.; Guenet, B.; Zhou, Y.; Su, J.Q.; Janssens, I.A. Priming of soil organic matter decomposition scales linearly with microbial biomass response to litter input in steppe vegetation. Oikos 2015, 124, 649–657. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (cm) | 0–10 | 10–20 |
|---|---|---|
| Water content (%) | 83.50 ± 2.08 | 82.66 ± 2.16 |
| TC (mg g−1) | 366.33 ± 8.20 | 384.45 ± 14.57 |
| TN (mg g−1) | 15.65 ± 0.52 | 18.68 ± 1.06 |
| MBC (mg kg−1) | 1421.60 ± 482.45 | 1478.70 ± 85.86 |
| DOC (mg kg−1) | 620.25 ± 81.92 | 338.42 ± 44.03 |
| NH4+-N (mg kg−1) | 66.60 ± 17.05 | 47.91 ± 6.54 |
| NO3−-N (mg kg−1) | 0.55 ± 0.07 | 0.58 ± 0.09 |
| Invertase (mg glucose g−1 soil 24 h−1) | 324.84 ± 23.30 | 213.98 ± 50.37 |
| β-Glucosidase (mg pNP kg−1 h−1) | 2215.30 ± 41.75 | 2732.10 ± 47.49 |
| Urease (mg N-NH4+ g−1 soil 24 h−1) | 4.88 ± 0.57 | 3.21 ± 0.78 |
| MBC | DOC | NH4+-N | NO3-N | Invertase | β-Glucosidase | Urease | |
|---|---|---|---|---|---|---|---|
| Cumulative SOC mineralization | −0.25 | −0.02 | 0.46 ** | 0.62 ** | −0.62 ** | 0.25 | −0.62 ** |
| MBC | −0.26 | −0.35 | −0.35 * | 0.23 | 0.17 | 0.07 | |
| DOC | 0.78 ** | 0.66 ** | 0.25 | −0.58 ** | 0.44 * | ||
| NH4+-N | 0.94 ** | −0.04 | −0.46 ** | 0.11 | |||
| NO3-N | −0.24 | −0.28 | −0.12 | ||||
| Invertase | −0.54 ** | 0.84 ** | |||||
| β-Glucosidase | −0.72 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.; Ma, X.; Song, Y.; Gao, S.; Ren, J.; Zhang, H.; Wang, X. Warming-Induced Labile Carbon Change Soil Organic Carbon Mineralization and Microbial Abundance in a Northern Peatland. Microorganisms 2022, 10, 1329. https://doi.org/10.3390/microorganisms10071329
Jiang L, Ma X, Song Y, Gao S, Ren J, Zhang H, Wang X. Warming-Induced Labile Carbon Change Soil Organic Carbon Mineralization and Microbial Abundance in a Northern Peatland. Microorganisms. 2022; 10(7):1329. https://doi.org/10.3390/microorganisms10071329
Chicago/Turabian StyleJiang, Lei, Xiuyan Ma, Yanyu Song, Siqi Gao, Jiusheng Ren, Hao Zhang, and Xianwei Wang. 2022. "Warming-Induced Labile Carbon Change Soil Organic Carbon Mineralization and Microbial Abundance in a Northern Peatland" Microorganisms 10, no. 7: 1329. https://doi.org/10.3390/microorganisms10071329