
Attachment of Ferredoxin: NADP+ Oxidoreductase to Phycobilisomes Is Required for Photoheterotrophic Growth of the Cyanobacterium Synechococcus sp. PCC 7002

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions and Mutant Construction
2.2. Protein Extraction and Immunoblotting
2.3. Fluorescence Measurements
2.4. P700 Measurement
3. Results
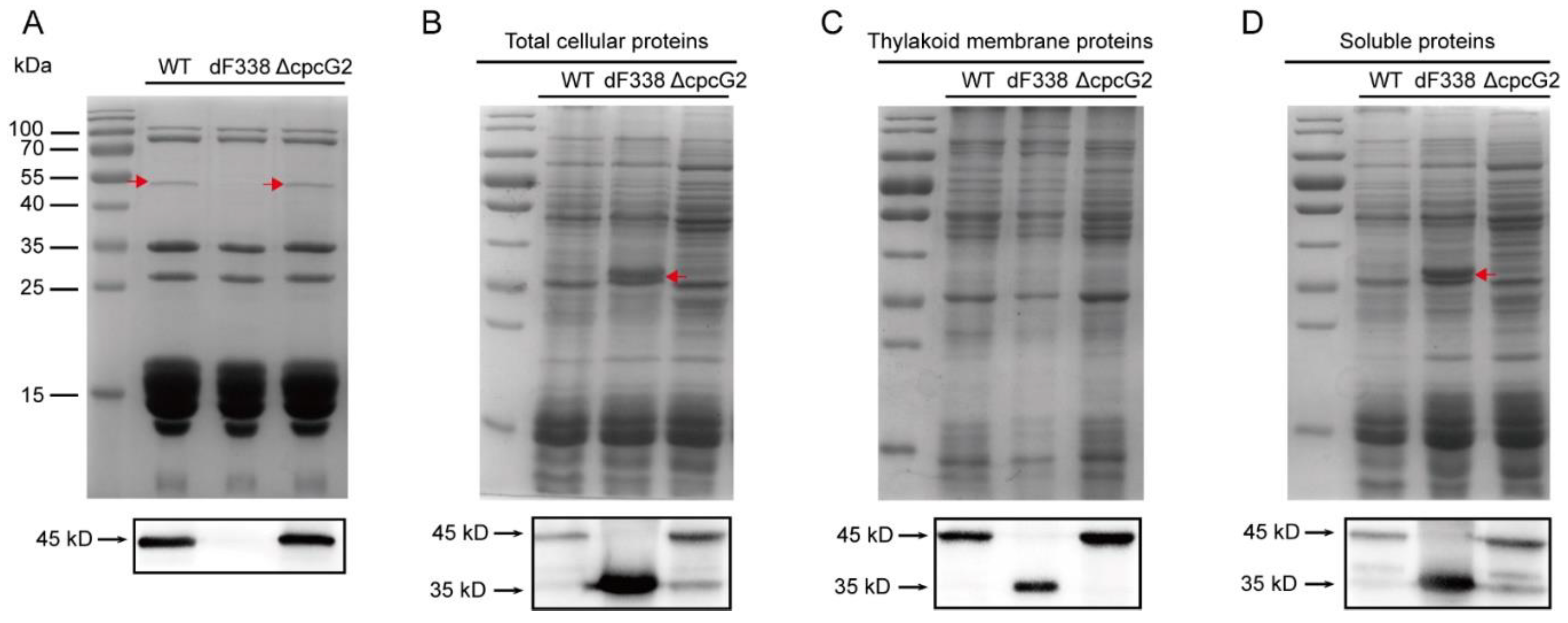
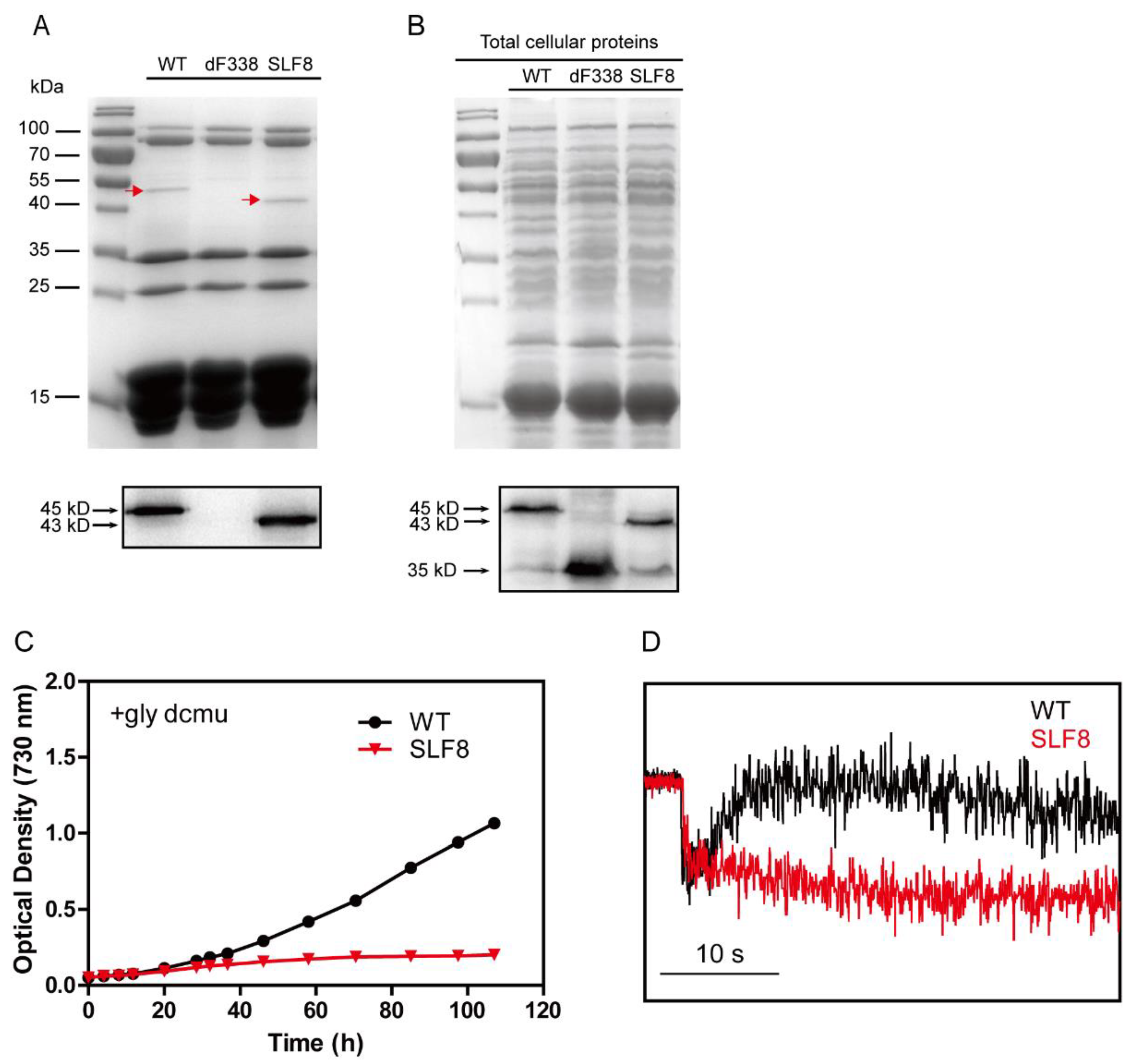
3.1. The CpcD Domain Is Responsible for Association of FNR with PBS
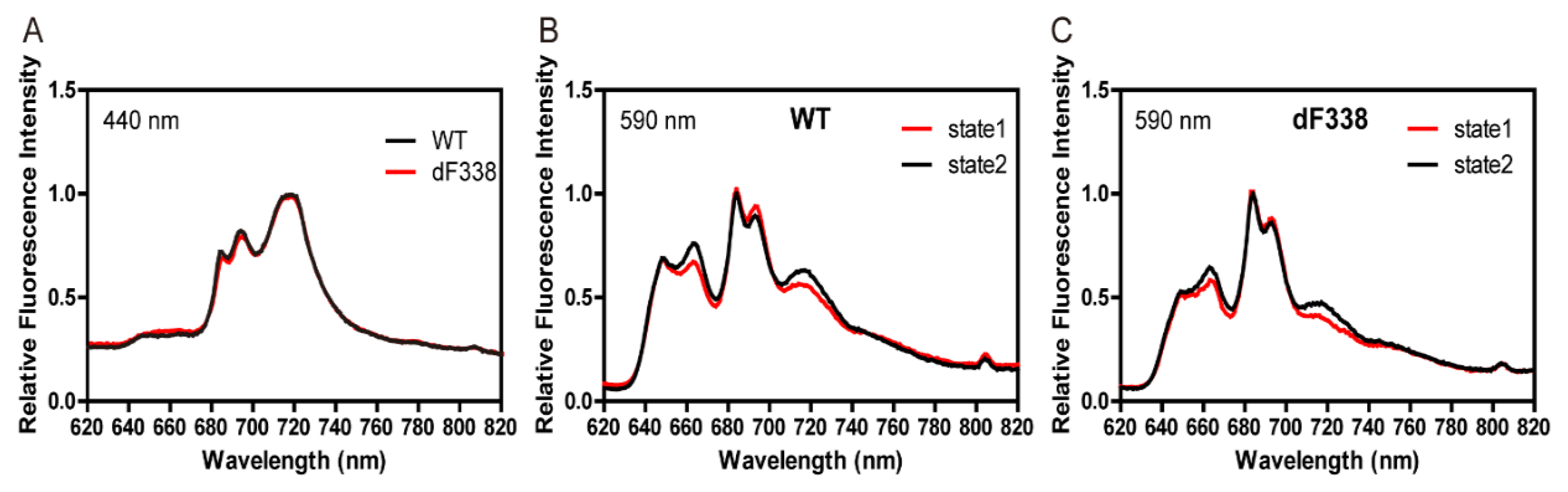
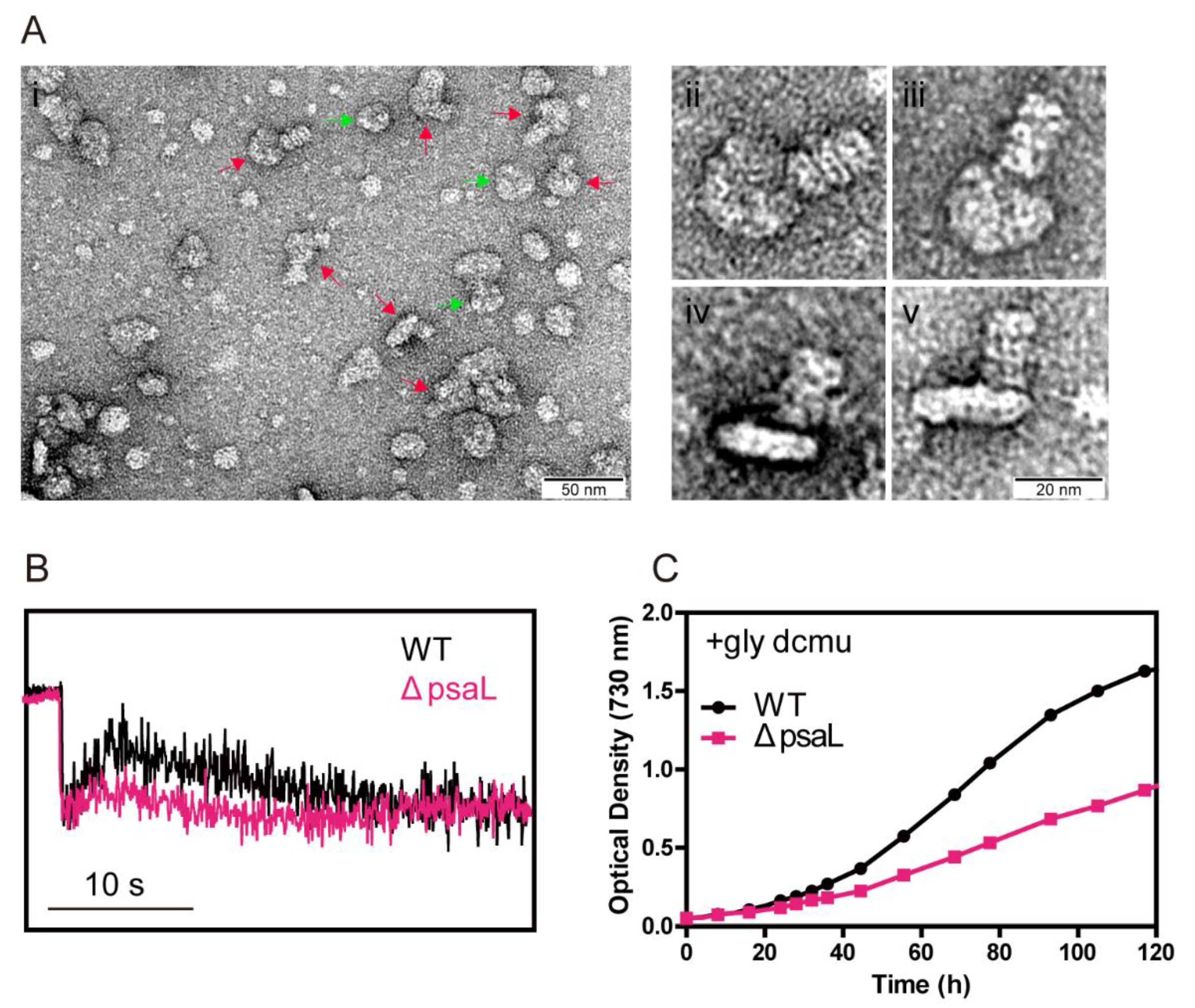
3.2. The Strain dF338 Is Able to Perform State Transitions
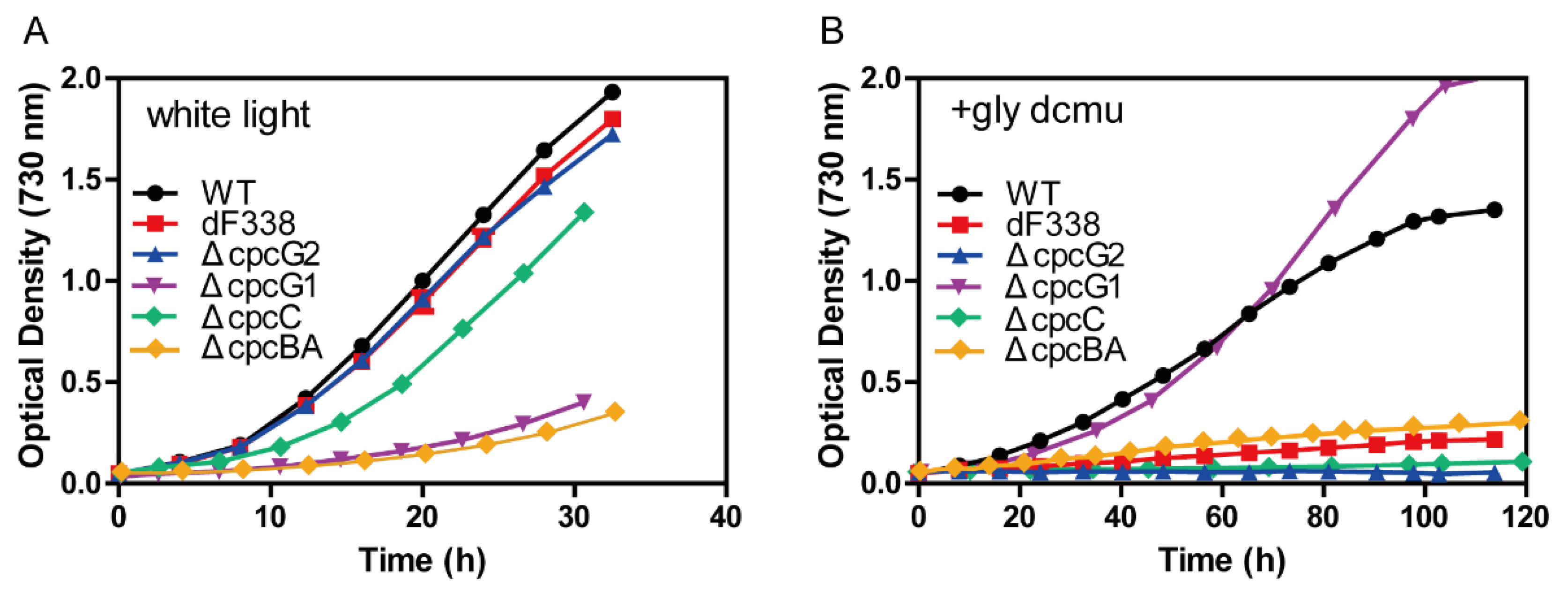
3.3. The Strain dF338 Is Impaired of Photoheterotrophic Growth
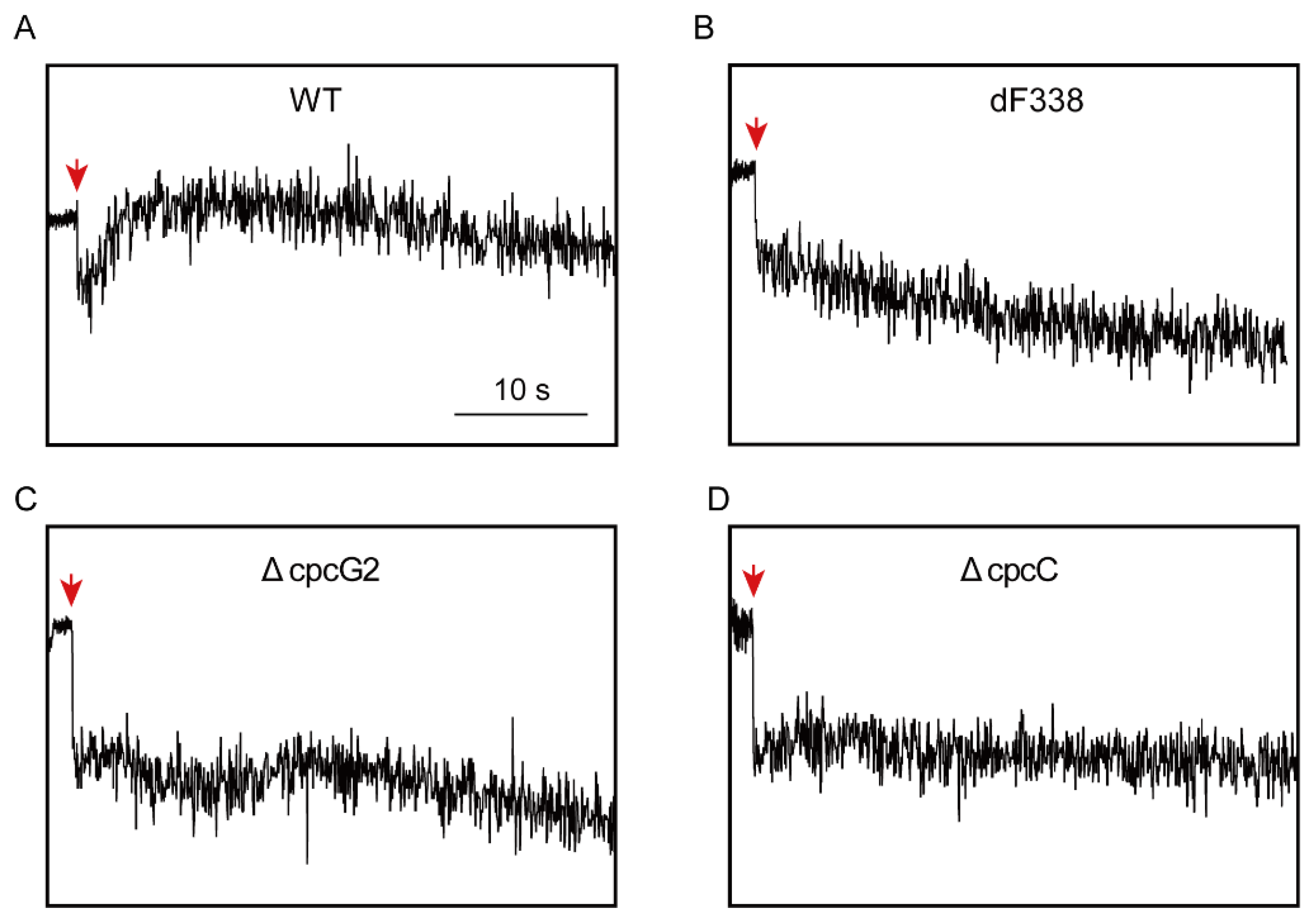
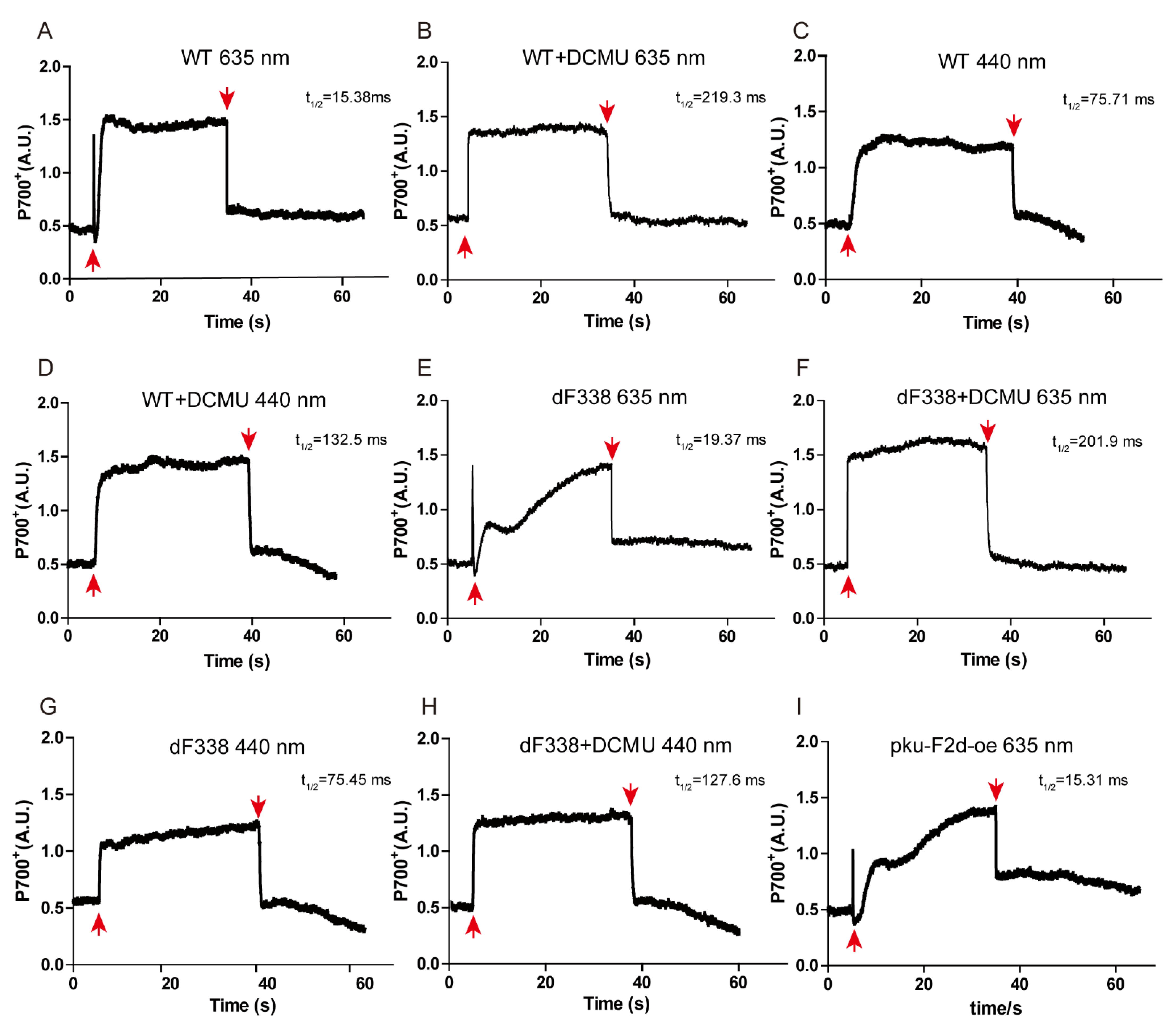
3.4. Photosynthetic Electron Transfers
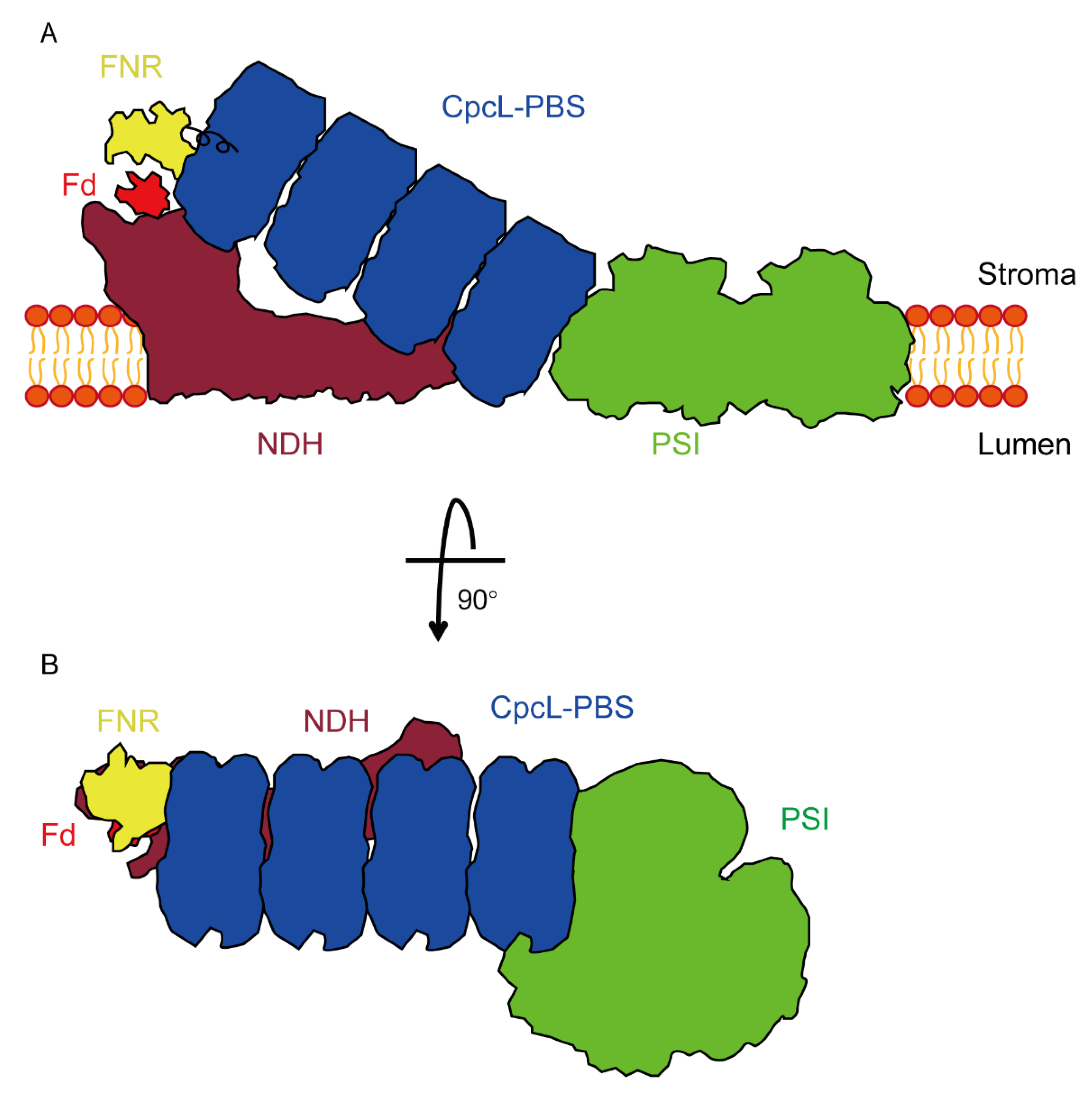
3.5. Association of CpcL-PBS with PSI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glazer, A.N. Light guides. Directional energy transfer in a photosynthetic antenna. J. Biol. Chem. 1989, 264, 1–4. [Google Scholar] [CrossRef]
- MacColl, R. Cyanobacterial phycobilisomes. J. Struct. Biol. 1998, 124, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Adir, N.; Bar-Zvi, S.; Harris, D. The amazing phycobilisome. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148047. [Google Scholar] [CrossRef]
- Sui, S.F. Structure of phycobilisomes. Annu. Rev. Biophys. 2021, 50, 53–72. [Google Scholar] [CrossRef]
- Zheng, L.; Zheng, Z.; Li, X.; Wang, G.; Zhang, K.; Wei, P.; Zhao, J.; Gao, N. Structural insight into the mechanism of energy transfer in cyanobacterial phycobilisomes. Nat. Commun. 2021, 12, 5497. [Google Scholar] [CrossRef] [PubMed]
- De Lorimier, R.; Guglielmi, G.; Bryant, D.A.; Stevens, S.E., Jr. Structure and mutation of a gene encoding a Mr 33.000 phycocyanin-associated linker polypeptide. Arch. Microbiol. 1990, 153, 541–549. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, J.; Liu, D.; Qin, S.; Sun, S.; Zhao, J.; Sui, S.F. Structure of phycobilisome from the red alga Griffithsia pacifica. Nature 2017, 551, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.H.; Glazer, A.N. Cyanobacterial phycobilisomes role of the linker polypeptides in the assembly of phycocyanin. J. Biol. Chem. 1982, 257, 3429–3433. [Google Scholar] [CrossRef]
- Chang, L.; Liu, X.; Li, Y.; Liu, C.C.; Yang, F.; Zhao, J.; Sui, S.F. Structural organization of an intact phycobilisome and its association with photosystem II. Cell Res. 2015, 25, 726–737. [Google Scholar] [CrossRef] [Green Version]
- De Lorimier, R.; Bryant, D.A.; Stevens, S.E., Jr. Genetic analysis of a 9 kDa phycocyanin-associated linker polypeptide. Biochim. Biophys. Acta 1990, 1019, 29–41. [Google Scholar] [CrossRef]
- Kondo, K.; Geng, X.X.; Katayama, M.; Ikeuchi, M. Distinct roles of CpcG1 and CpcG2 in phycobilisome assembly in the cyanobacterium Synechocystis sp. PCC 6803. Photosynth. Res. 2005, 84, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Ochiai, Y.; Katayama, M.; Ikeuchi, M. The membrane-associated CpcG2-phycobilisome in Synechocystis: A new photosystem I antenna. Plant Physiol. 2007, 144, 1200–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, D.A.; Stirewalt, V.L.; Glauser, M.; Frank, G.; Sidler, W.; Zuber, H. A small multigene family encodes the rod-core linker polypeptides of Anabaena sp. PCC 7120 phycobilisomes. Gene 1991, 107, 91–99. [Google Scholar] [CrossRef]
- Deng, G.; Liu, F.; Liu, X.; Zhao, J. Significant energy transfer from CpcG2-phycobilisomes to photosystem I in the cyanobacterium Synechococcus sp. PCC 7002 in the absence of ApcD-dependent state transitions. FEBS Lett. 2012, 586, 2342–2345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Semchonok, D.A.; Webber-Birungi, M.T.; Ehira, S.; Kondo, K.; Narikawa, R.; Ohmori, M.; Boekema, E.J.; Ikeuchi, M. Attachment of phycobilisomes in an antenna-photosystem I supercomplex of cyanobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, 2512–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedzwiedzki, D.M.; Liu, H.; Blankenship, R.E. Excitation energy transfer in intact CpcL-phycobilisomes from Synechocystis sp. PCC 6803. J. Phys. Chem. B 2019, 123, 4695–4704. [Google Scholar] [CrossRef]
- Chereskin, B.M.; Clement-Metral, J.D.; Gantt, E. Characterization of a purified photosystem II-phycobilisome particle preparation from Porphyridium cruentum. Plant Physiol. 1985, 77, 626–629. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.T.; Stevens, C.L.R.; Myers, J. Action spectra for photoreactions I and II of photosynthesis in the blue-green alga Anacystis nidulans. Photochem. Photobiol. 1977, 25, 103–108. [Google Scholar] [CrossRef]
- Bonaventura, C.; Myers, J. Fluorescence and oxygen evolution from Chlorella pyrenoidosa. Biochim. Biophys. Acta 1969, 189, 366–383. [Google Scholar] [CrossRef]
- Mullineaux, C.W. Excitation-energy transfer from phycobilisomes to photosystem-I in a cyanobacterium. Biochim. Biophys. Acta 1992, 1100, 285–292. [Google Scholar] [CrossRef]
- Dong, C.; Tang, A.; Zhao, J.; Mullineaux, C.W.; Shen, G.; Bryant, D.A. ApcD is necessary for efficient energy transfer from phycobilisomes to photosystem I and helps to prevent photoinhibition in the cyanobacterium Synechococcus sp. PCC 7002. Biochim. Biophys. Acta 2009, 1787, 1122–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliverti, A.; Pandini, V.; Pennati, A.; de Rosa, M.; Zanetti, G. Structural and functional diversity of ferredoxin-NADP+ reductases. Arch. Biochem. Biophys. 2008, 474, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Bruns, C.M.; Karplus, P.A. Refined crystal structure of spinach ferredoxin reductase at 1.7 A resolution: Oxidized, reduced and 2’-phospho-5’-AMP bound states. J. Mol. Biol. 1995, 247, 125–145. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, J.A.; Mayoral, T.; Faro, M.; Gómez-Moreno, C.; Sanz-Aparicio, J.; Medina, M. Mechanism of coenzyme recognition and binding revealed by crystal structure analysis of ferredoxin-NADP+ reductase complexed with NADP+. J. Mol. Biol. 2002, 319, 1133–1142. [Google Scholar] [CrossRef]
- Schluchter, W.M.; Bryant, D.A. Molecular characterization of ferredoxin-NADP+ oxidoreductase in cyanobacteria: Cloning and sequence of the petH gene of Synechococcus sp. PCC 7002 and studies on the gene product. Biochemistry 1992, 31, 3092–3102. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, N.; Wei, T.D.; Su, H.N.; Xie, B.B.; Dong, C.C.; Zhang, X.Y.; Chen, X.L.; Zhou, B.C.; Wang, Z.X.; et al. Crystal structure of the N-terminal domain of linker L(R) and the assembly of cyanobacterial phycobilisome rods. J. Mol. Biol. 2011, 82, 698–705. [Google Scholar] [CrossRef]
- van Thor, J.J.; Gruters, O.W.; Matthijs, H.C.; Hellingwerf, K.J. Localization and function of ferredoxin:NADP+ reductase bound to the phycobilisomes of Synechocystis. EMBO J. 1999, 18, 4128–4136. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Lojero, C.; Perez-Gómez, B.; Shen, G.; Schluchter, W.M.; Bryant, D.A. Interaction of ferredoxin:NADP+ oxidoreductase with phycobilisomes and phycobilisome substructures of the cyanobacterium Synechococcus sp. strain PCC 7002. Biochemistry 2003, 42, 13800–13811. [Google Scholar] [CrossRef]
- Liu, H.; Weisz, D.A.; Zhang, M.M.; Cheng, M.; Zhang, B.; Zhang, H.; Gerstenecker, G.S.; Pakrasi, H.B.; Gross, M.L.; Blankenship, R.E. Phycobilisomes Harbor FNRL in Cyanobacteria. MBio 2019, 10, e00669-19. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.C.; Ughy, B.; Lagoutte, B.; Ajlani, G. A second isoform of the ferredoxin: NADP oxidoreductase generated by an in-frame initiation of translation. Proc. Natl. Acad. Sci. USA 2006, 103, 18368–18373. [Google Scholar] [CrossRef] [Green Version]
- Omairi-Nasser, A.; de Gracia, A.G.; Ajlani, G. A larger transcript is required for the synthesis of the smaller isoform of ferredoxin: NADP oxidoreductase. Mol. Microbiol. 2011, 81, 1178–1189. [Google Scholar] [CrossRef]
- Korn, A.; Ajlani, G.; Lagoutte, B.; Gall, A.; Sétif, P. Ferredoxin:NADP+ oxidoreductase association with phycocyanin modulates its properties. J. Biol. Chem. 2009, 284, 31789–31797. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, W.J.; Bailleul, B.; Picot, D.; Cardol, P.; Rappaport, F.; Wollman, F.A.; Joliot, P. The mechanism of cyclic electron flow. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 433–438. [Google Scholar] [CrossRef]
- Ogawa, T.; Mi, H. Cyanobacterial NADPH dehydrogenase complexes. Photosynth. Res. 2007, 93, 69–77. [Google Scholar] [CrossRef]
- Peltier, G.; Aro, E.M.; Shikanai, T. NDH-1 and NDH-2 plastoquinone reductases in oxygenic photosynthesis. Annu. Rev. Plant Biol. 2016, 67, 55–80. [Google Scholar] [CrossRef]
- Schuller, J.M.; Birrell, J.A.; Tanaka, H.; Konuma, T.; Wulfhorst, H.; Cox, N.; Schuller, S.K.; Thiemann, J.; Lubitz, W.; Sétif, P.; et al. Structural adaptations of photosynthetic complex I enable ferredoxin-dependent electron transfer. Science 2019, 363, 257–260. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, T.G.; Bayne, A.N.; Trempe, J.F.; Savage, D.F.; Davies, K.M. Structure of the complex I-like molecule NDH of oxygenic photosynthesis. Nature 2019, 566, 411–414. [Google Scholar] [CrossRef]
- Iwai, M.; Takizawa, K.; Tokutsu, R.; Okamuro, A.; Takahashi, Y.; Minagawa, J. Isolation of the elusive supercomplex that drives cyclic electron flow in photosynthesis. Nature 2010, 464, 1210–1213. [Google Scholar] [CrossRef]
- Shen, L.; Tang, K.; Wang, W.; Wang, C.; Wu, H.; Mao, Z.; An, S.; Chang, S.; Kuang, T.; Shen, J.R.; et al. Architecture of the chloroplast PSI-NDH supercomplex in Hordeum vulgare. Nature 2021, 601, 649–654. [Google Scholar] [CrossRef]
- Gao, F.D.; Zhao, J.H.; Chen, L.P.; Battchikova, N.; Ran, Z.X.; Aro, E.M.; Ogawa, T.; Ma, W.M. The NDH-1L-PSI supercomplex is important for efficient cyclic electron transport in cyanobacteria. Plant Physiol. 2016, 172, 1451–1464. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.M.; Whitelegge, J.P.; Cramer, W.A. Ferredoxin:NADP+ oxidoreductase is a subunit of the chloroplast cytochrome b6f complex. J. Biol. Chem. 2001, 276, 38159–38165. [Google Scholar] [CrossRef]
- Stevens, S.E., Jr.; Patterson, C.O.P.; Myers, J. The production of hydrogen peroxide by blue-green algae: A survey. J. Phycol. 1973, 9, 427–430. [Google Scholar] [CrossRef]
- Alcántara-Sánchez, F.; Leyva-Castillo, L.E.; Chagolla-López, A.; González de la Vara, L.; Gómez-Lojero, C. Distribution of isoforms of ferredoxin-NADP+ reductase (FNR) in cyanobacteria in two growth conditions. Int. J. Biochem. Cell Biol. 2017, 85, 123–134. [Google Scholar] [CrossRef]
- Schluchter, W.M.; Shen, G.; Zhao, J.; Bryant, D.A. Characterization of psaI and psaL mutants of Synechococcus sp. strain PCC 7002: A new model for state transitions in cyanobacteria. Photochem. Photobiol. 1996, 64, 53–66. [Google Scholar] [CrossRef]
- Bryant, D.A. Genetic analysis of phycobilisome biosynthesis, assembly, structure and function in the cyanobacterium Synechococcus sp. PCC 7002. In Light-Energy Transduction in Photosynthesis: Higher Plants and Bacterial Models.; Stevens, S.E., Jr., Bryant, D.A., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 1988; pp. 62–90. [Google Scholar]
- Huang, C.; Yuan, X.; Zhao, J.; Bryant, D.A. Kinetic analyses of state transitions of the cyanobacterium Synechococcus sp. PCC 7002 and its mutant strains impaired in electron transport. Biochim. Biophys. Acta 2003, 1607, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Berla, B.M.; Saha, R.; Immethun, C.M.; Maranas, C.D.; Moon, T.S.; Pakrasi, H.B. Synthetic biology of cyanobacteria: Unique challenges and opportunities. Front. Microbiol. 2013, 4, 246. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Li, Y.; Li, X.; Zhong, Q.; Li, N.; Zhang, K.; Zhang, Y.; Chu, H.; Ma, C.; Li, G.; et al. Structural and functional insights into the tetrameric photosystem I from heterocyst-forming cyanobacteria. Nat. Plants 2019, 5, 1087–1097. [Google Scholar] [CrossRef]
- Noji, T.; Watanabe, M.; Dewa, T.; Itoh, S.; Ikeuchi, M. Direct energy transfer from allophycocyanin-free rod-type CpcL-phycobilisome to photosystem I. J. Phys. Chem. Lett. 2021, 12, 6692–6697. [Google Scholar] [CrossRef]
- Dong, C.; Zhao, J. ApcD is required for state transition but not involved in blue-light induced quenching in the cyanobacterium Anabaena sp. PCC 7120. Sci. Bull. 2008, 53, 3422–3424. [Google Scholar] [CrossRef]
- Holland, S.C.; Kappell, A.D.; Burnap, R.L. Redox changes accompanying inorganic carbon limitation in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 2015, 1847, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Korn, A. Respective Roles of the Ferredoxin: NADP-Oxidoreductase Isoforms in the Cyanobacterium Synechocystis sp. PCC 6803. Ph.D. Thesis, Université Paris Sud, Paris, France, November 2010. [Google Scholar]
- Miller, N.T.; Ajlani, G.; Burnap, R.L. Cyclic electron flow-coupled proton pumping in Synechocystis sp. PCC 6803 is dependent upon NADPH Oxidation by the soluble isoform of Ferredoxin:NADP-Oxidoreductase. Microorganisms 2022, 10, 855. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, Y.; Chen, Z.; Zhang, J.; Lin, Y.; Wang, Y.; Xiong, Q.; Li, T.; Ge, F.; Bryant, D.A.; et al. Proteogenomic analysis and global discovery of posttranslational modifications in prokaryotes. Proc. Natl. Acad. Sci. USA 2014, 111, E5633–E5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, H.; Endo, T.; Ogawa, T.; Asada, K. Thylakoid membrane-bound, NADPH-specific pyridine nucleotide dehydrogenase complex mediates cyclic electron transport in the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 1995, 36, 661–668. [Google Scholar]
- Laughlin, T.G.; Savage, D.F.; Davies, K.M. Recent advances on the structure and function of NDH-1: The complex I of oxygenic photosynthesis. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148254. [Google Scholar] [CrossRef]
- Ogawa, T. A gene homologous to the subunit-2 gene of NADH dehydrogenase is essential to inorganic carbon transport of Synechocystis PCC 6803. Proc. Natl. Acad. Sci. USA 1991, 88, 4275–4279. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Zhao, J.; Muhlenhoff, U.; Bryant, D.A.; Golbeck, J.H. PsaE is required for in vivo cyclic electron flow around photosystem I in the cyanobacterium Synechococcus sp. PCC 7002. Plant Physiol. 1993, 103, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Miller, N.T.; Vaughn, M.D.; Burnap, R.L. Electron flow through NDH-1 complexes is the major driver of cyclic electron flow-dependent proton pumping in cyanobacteria. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148354. [Google Scholar] [CrossRef]
- Zhang, C.; Shuai, J.; Ran, Z.; Zhao, J.; Wu, Z.; Liao, R.; Wu, J.; Ma, W.; Lei, M. Structural insights into NDH-1 mediated cyclic electron transfer. Nat. Commun. 2020, 11, 888. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Cao, D.; Xie, F.; Xu, F.; Su, X.; Mi, H.; Zhang, X.; Li, M. Structural basis for electron transport mechanism of complex I-like photosynthetic NAD(P)H dehydrogenase. Nat. Commun. 2020, 11, 610. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Huang, C.; Wei, P.; Zhang, K.; Dong, C.; Lan, Q.; Zheng, Z.; Zhang, Z.; Zhao, J. Attachment of Ferredoxin: NADP+ Oxidoreductase to Phycobilisomes Is Required for Photoheterotrophic Growth of the Cyanobacterium Synechococcus sp. PCC 7002. Microorganisms 2022, 10, 1313. https://doi.org/10.3390/microorganisms10071313
Li X, Huang C, Wei P, Zhang K, Dong C, Lan Q, Zheng Z, Zhang Z, Zhao J. Attachment of Ferredoxin: NADP+ Oxidoreductase to Phycobilisomes Is Required for Photoheterotrophic Growth of the Cyanobacterium Synechococcus sp. PCC 7002. Microorganisms. 2022; 10(7):1313. https://doi.org/10.3390/microorganisms10071313
Chicago/Turabian StyleLi, Xiying, Chenhui Huang, Peijun Wei, Kun Zhang, Chunxia Dong, Qing Lan, Zhenggao Zheng, Zhengdong Zhang, and Jindong Zhao. 2022. "Attachment of Ferredoxin: NADP+ Oxidoreductase to Phycobilisomes Is Required for Photoheterotrophic Growth of the Cyanobacterium Synechococcus sp. PCC 7002" Microorganisms 10, no. 7: 1313. https://doi.org/10.3390/microorganisms10071313