
Interaction of Microbiota between Fish and the Environment of an In-Pond Raceway System in a Lake

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Water Environment Factors
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Data Analysis
3. Results
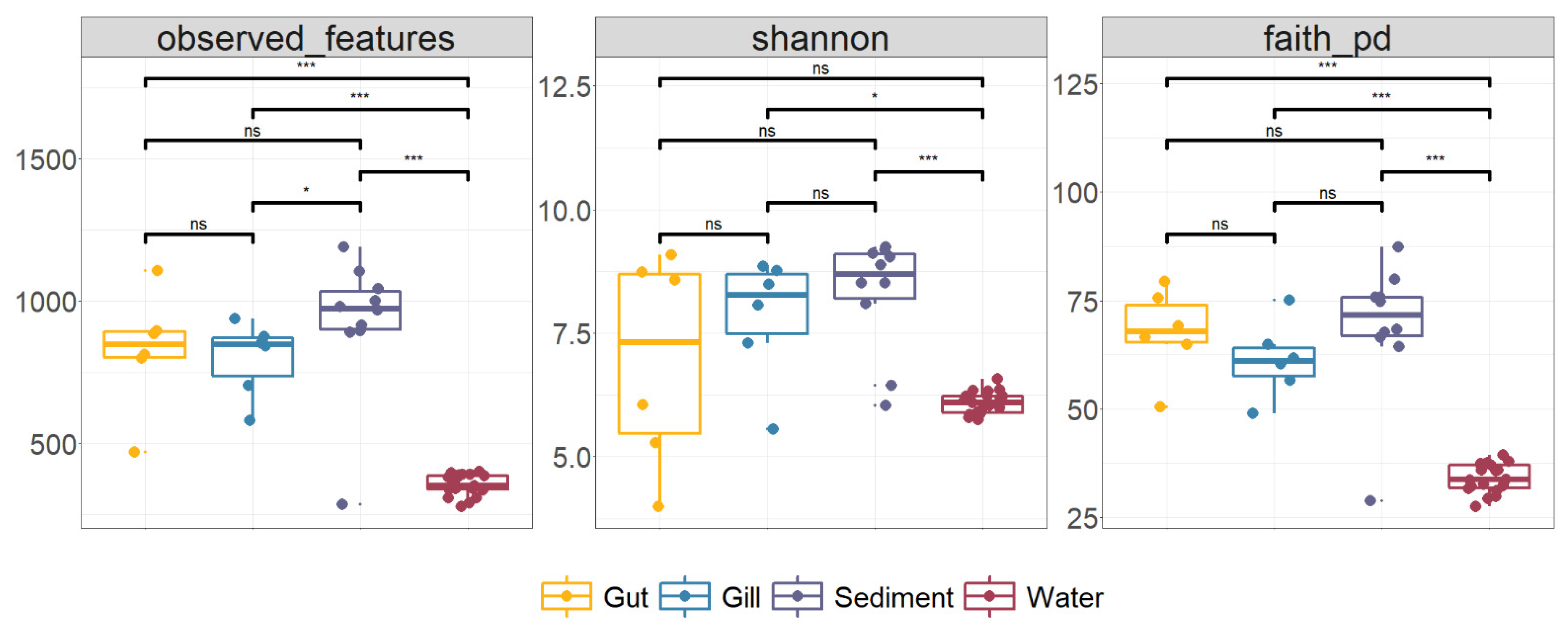
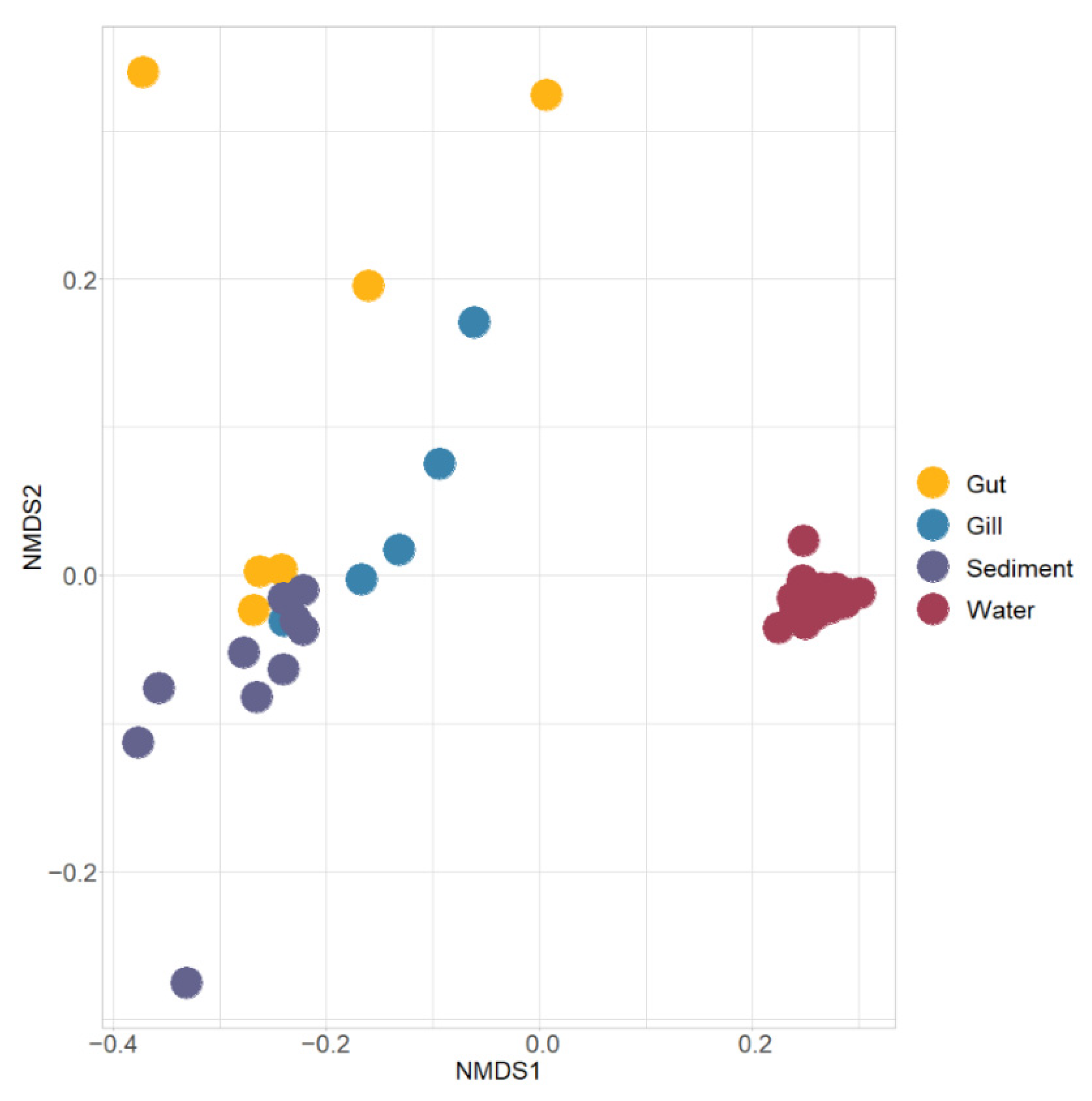
3.1. Diversity of Microbiota
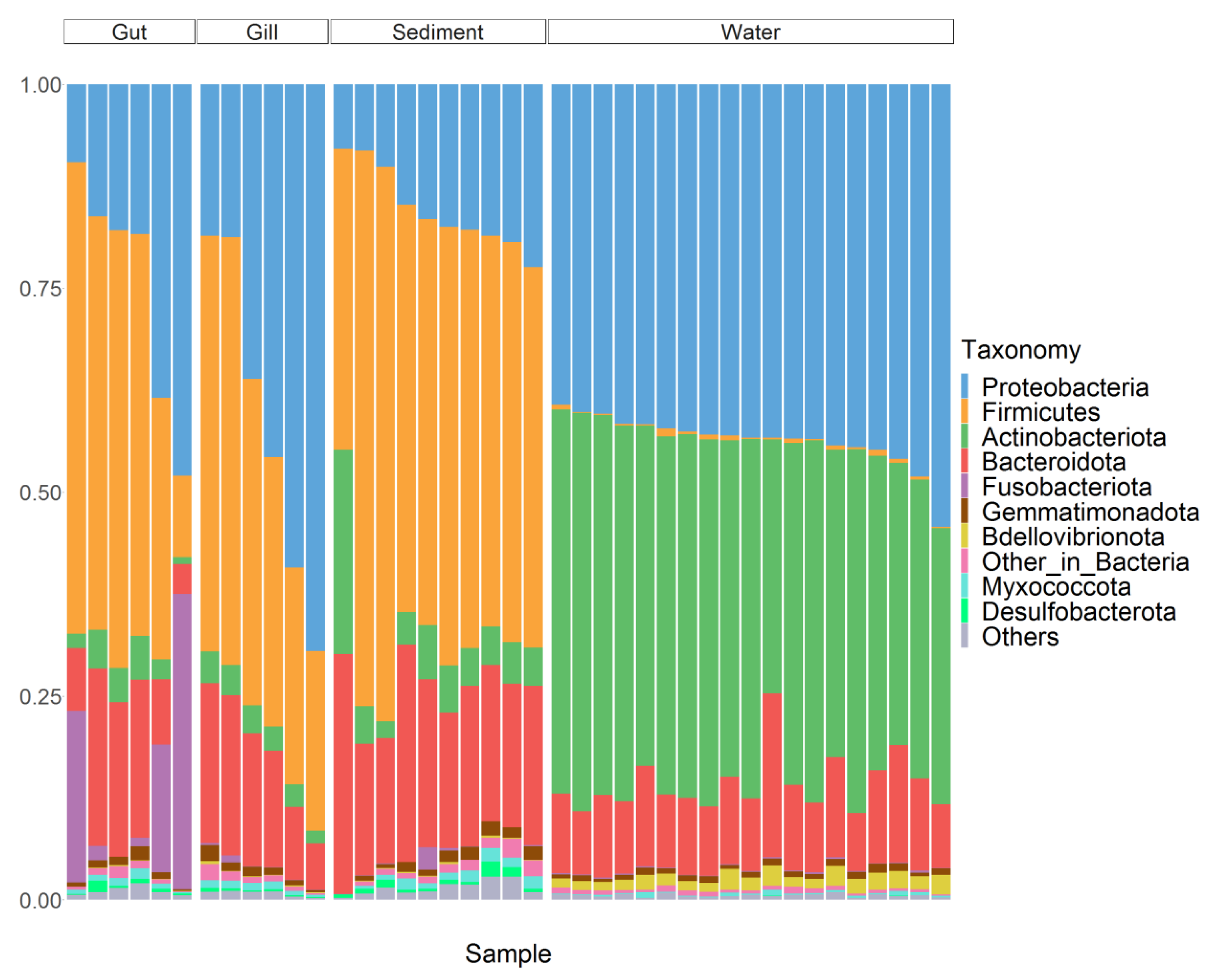
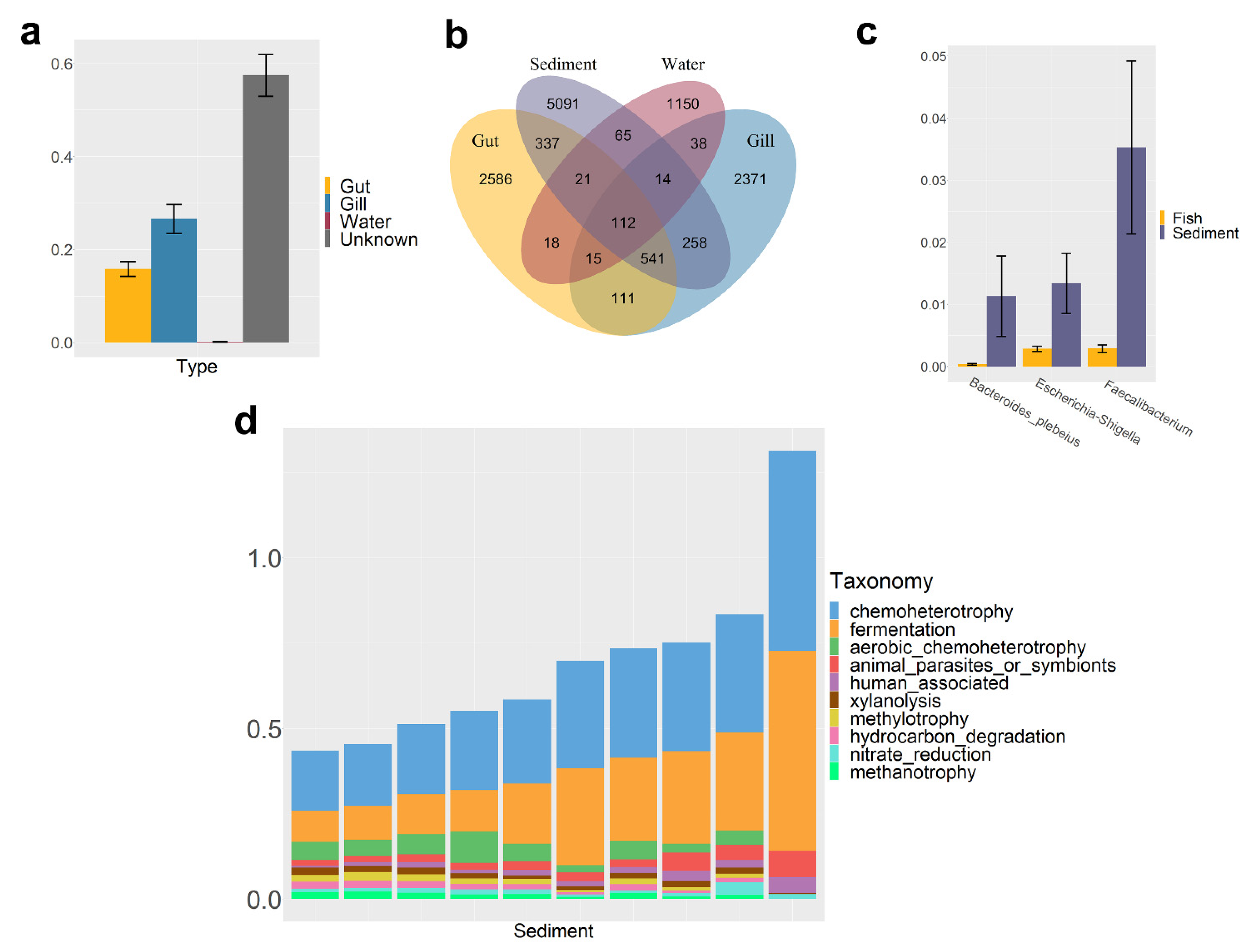
3.2. The Composition of Fish Microbiota
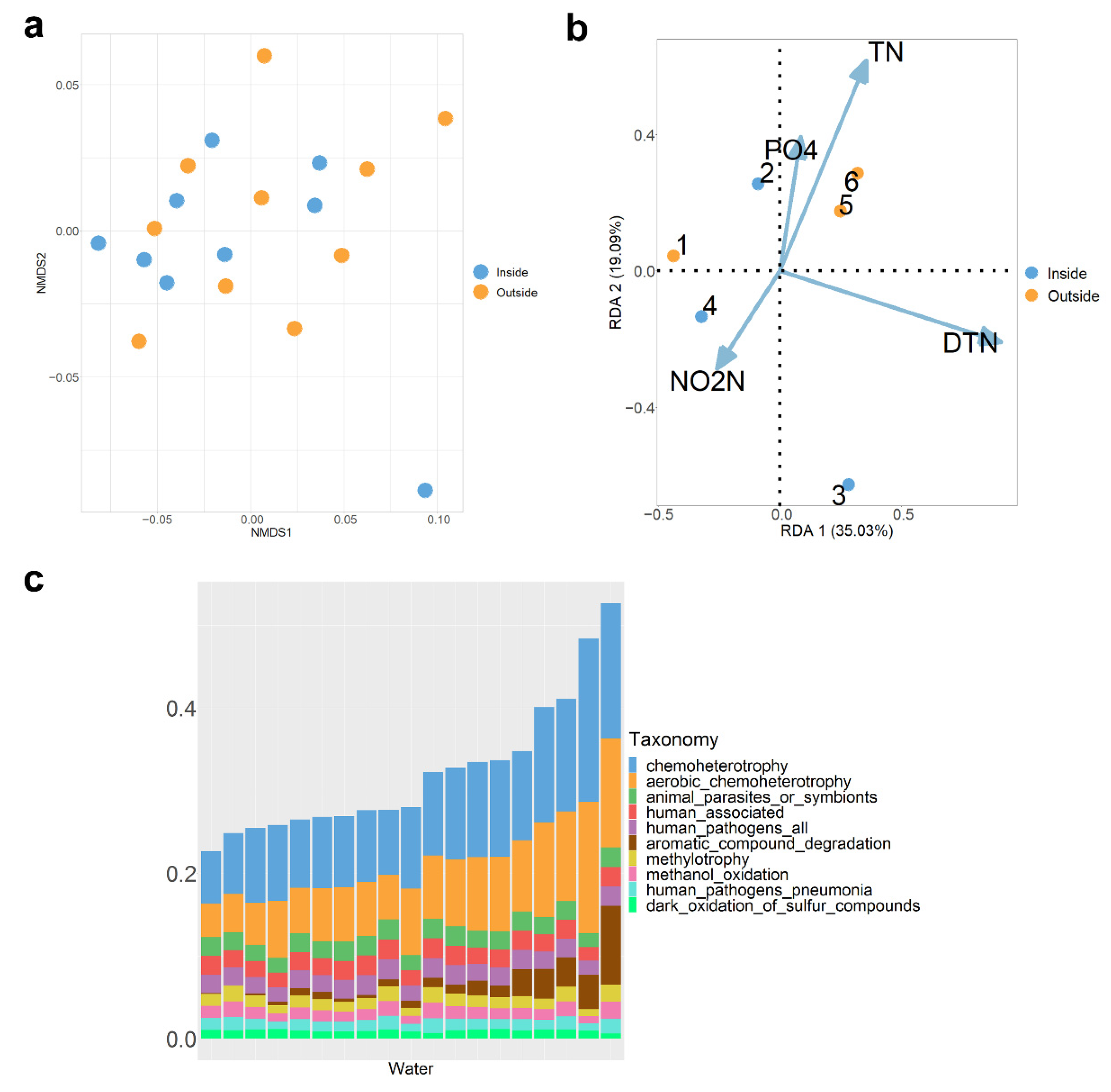
3.3. Characteristics of Water Microbiota
3.4. Characteristics of Sediment Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, F.; Zhong, H.; Wu, C.; Wang, S.; Guo, Z.; Tao, M.; Zhang, C.; Gong, D.; Gao, X.; Tang, C.; et al. Development of fisheries in China. Reprod. Breed. 2021, 1, 64–79. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Q.; Zhang, T.; Ye, S.; Li, W.; Yuan, J.; Li, Z. Development of Lake and Reservoir Aquaculture Related Practices in China. In Aquaculture in China: Success Stories and Modern Trends; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 599–610. [Google Scholar] [CrossRef]
- Bureau of Fisheries. China Fishery Statistical Yearbook; China Statistics Press: Beijing, China, 2019. [Google Scholar]
- Guo, L.; Li, Z.; Xie, P.; Ni, L. Assessment effects of cage culture on nitrogen and phosphorus dynamics in relation to fallowing in a shallow lake in China. Aquac. Int. 2008, 17, 229–241. [Google Scholar] [CrossRef]
- Guo, L.; Li, Z. Effects of nitrogen and phosphorus from fish cage-culture on the communities of a shallow lake in middle Yangtze River basin of China. Aquaculture 2003, 226, 201–212. [Google Scholar] [CrossRef]
- Jiang, Z.J.; Fang, J.G.; Mao, Y.Z.; Wang, W. Eutrophication assessment and bioremediation strategy in a marine fish cage culture area in Nansha Bay, China. J. Appl. Phycol. 2009, 22, 421–426. [Google Scholar] [CrossRef]
- Bojarski, B.; Jakubiak, M.; Szczerbik, P.; Bień, M.; Klaczak, A.; Stański, T.; Witeska, M. The Influence of Fish Ponds on Fish Assemblages of Adjacent Watercourses. Pol. J. Environ. Stud. 2022, 31, 609–617. [Google Scholar] [CrossRef]
- Jakubiak, M.; Bojarski, B.; Bień, M.; Stonawski, B.; Oglęcki, P. Influence of Fish Ponds on the Benthic Invertebrate Composition in Hydrological Networks of Selected Fish Farms in Southern Poland. Folia Biol. 2022, 70, 11–18. [Google Scholar] [CrossRef]
- Li, W.; Cheng, X.; Xie, J.; Wang, Z.; Yu, D. Hydrodynamics of an in-pond raceway system with an aeration plug-flow device for application in aquaculture: An experimental study. R. Soc. Open Sci. 2019, 6, 182061. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.; Cheng, X.; Xie, J.; Zhu, D. Assessment of aeration plug-flow devices used with recirculating aquaculture systems on the growth of tilapia Oreochromis niloticus. Aquac. Eng. 2020, 91, 102116. [Google Scholar] [CrossRef]
- Yu, W.; Wang, J. Application and Analysis of Cluster-Type Internal Recirculation Pond Flowing Water Aquaculture Facilities. Fish. Mod. 2016, 43, 1–5. (In Chinese) [Google Scholar]
- Wang, Y.; Ni, J.; Nie, Z.; Gao, J.; Sun, Y.; Shao, N.; Li, Q.; Hu, J.; Xu, P.; Xu, G. Effects of Stocking Density on Growth, Serum Parameters, Antioxidant Status, Liver and Intestine Histology and Gene Expression of Largemouth Bass (Micropterus salmoides) Farmed in the in-Pond Raceway System. Aquac. Res. 2020, 51, 5228–5240. [Google Scholar] [CrossRef]
- Fan, Z.; Lifa, D.; Jian, P.; Liupu, C.; Xingwei, X.; Yijiang, B.E.I.; Wenjun, M.A.; Xueyan, D. Comparative Analysis of Muscle Nutrient Composition Bewteen In-Pond Raceway and Usual-Pond System Cultured Liza Haematocheila. Nfsckx 2021, 17, 70–76. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, P.; Nie, Z.; Li, Q.; Shao, N.; Xu, G. Growth, digestive enzymes activities, serum biochemical parameters and antioxidant status of juvenile genetically improved farmed tilapia (Oreochromis niloticus) reared at different stocking densities in in-pond raceway recirculating culture system. Aquac. Res. 2019, 50, 1338–1347. [Google Scholar] [CrossRef]
- FAO. FishStatJ, a Tool for Fishery Statistics Analysis; Release: 4.00. 10. Universal Software for Fishery Statistical Time Series; Fisheries and Aquaculture Division (NFI): Rome, Italy, 2020. [Google Scholar]
- Yuan, J.; Ni, M.; Liu, M.; Wang, H.; Zhang, C.; Mi, G.; Gu, Z. Analysis of the growth performances, muscle quality, blood biochemistry and antioxidant status of Micropterus salmoides farmed in in-pond raceway systems versus usual-pond systems. Aquaculture 2019, 511, 734241. [Google Scholar] [CrossRef]
- Ni, M.; Liu, M.; Lou, J.; Mi, G.; Yuan, J.; Gu, Z. Stocking density alters growth performance, serum biochemistry, digestive enzymes, immune response, and muscle quality of largemouth bass (Micropterus salmoides) in in-pond raceway system. Fish Physiol. Biochem. 2021, 47, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, S.; Nie, Z.; Li, Q.; Sun, Y.; Shao, N.; Gao, J.; Hu, J.; Xu, P.; Xu, G. Optimum feeding frequency of juvenile largemouth bass (Micropterus salmoides) reared in in-pond raceway recirculating culture system. Fish Physiol. Biochem. 2020, 46, 2197–2212. [Google Scholar] [CrossRef]
- Lastauskienė, E.; Valskys, V.; Stankevičiūtė, J.; Kalcienė, V.; Gėgžna, V.; Kavoliūnas, J.; Ružauskas, M.; Armalytė, J. The Impact of Intensive Fish Farming on Pond Sediment Microbiome and Antibiotic Resistance Gene Composition. Front. Vet. Sci. 2021, 8, 673756. [Google Scholar] [CrossRef]
- Li, J.; Fang, L.; Liang, X.-F.; Guo, W.; Lv, L.; Li, L. Influence of environmental factors and bacterial community diversity in pond water on health of Chinese perch through Gut Microbiota change. Aquac. Rep. 2021, 20, 100629. [Google Scholar] [CrossRef]
- Li, Q.; Guo, J.-L.; Wang, Y.-C.; Jiang, J.-H.; Sun, L.-H.; Mi, G.-Q.; Chen, J.-M.; Gu, Z.-M. On Changes of Intestinal Microbiota of a New Hybrid Strain of (Female Culter Alburnus) × (Male Megalobrama Terminalis) Reared in in-Pond Raceway Aquaculture System. Oceanol. Limnol. Sin. 2020, 51, 364–370. (In Chinese) [Google Scholar] [CrossRef]
- Liu, M.; Lian, Q.; Ni, M.; Guo, A.; Yuan, J. Effects of Inner-Pond Raceway Aquaculture on the Growth Performance, Antioxidant Enzymes, Digestive Enzymes, Digestive Tract Structure, and Bacterial Flora of Largemouth Bass (Micropterus salmoides). J. Fish. China 2021, 45, 2011–2028. (In Chinese) [Google Scholar]
- Wang, P.; Xu, G.; Xu, P. Temporal and Spatial Variation of Dissolved Oxygen and Reaction of Bacterial Community in In-Pond Raceway System (Iprs) of Largemouth Bass (Micropterus salmoides). ACTA Hydrobiol. Sin. 2019, 43, 1290–1299. (In Chinese) [Google Scholar]
- Hou, Y.; Li, B.; Xu, G.; Li, D.; Zhang, C.; Jia, R.; Li, Q.; Zhu, J. Dynamic and Assembly of Benthic Bacterial Community in an Industrial-Scale In-Pond Raceway Recirculating Culture System. Front. Microbiol. 2021, 12, 797817. [Google Scholar] [CrossRef] [PubMed]
- HJ 495-2009; Water Quality Technical Regulation on the Design of Sampling Programmes; China National Environment Monitoring Centre: Beijing, China; Liaoning Province Environmental Monitoring Center Station: Shenyang, China, 2009.
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, 9490. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors That Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; The Comprehensive R Archive Network: Vienna, Austria, 2020. [Google Scholar]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.T.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lai, Z.; Gao, Y.; Wang, C.; Zeng, Y.; Liu, E.; Mai, Y.; Yang, W.; Li, H. Connection between the Gut Microbiota of Largemouth Bass (Micropterus salmoides) and Microbiota of the Pond Culture Environment. Microorganisms 2021, 9, 1770. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.-M.; Hou, S.-W.; Wang, G.-X.; Gong, J.-Y.; Zhou, L.; Huang, Y.-H.; Huang, X.-D.; Liu, L. Effects of zymolytic black soldier fly (Hermetia illucens) pulp as dietary supplementation in largemouth bass (Micropterus salmoides). Aquac. Rep. 2021, 21, 100823. [Google Scholar] [CrossRef]
- Attramadal, K.J.; Øien, J.V.; Kristensen, E.; Evjemo, J.O.; Kjørsvik, E.; Vadstein, O.; Bakke, I. UV treatment in RAS influences the rearing water microbiota and reduces the survival of European lobster larvae (Homarus gammarus). Aquac. Eng. 2021, 94, 102176. [Google Scholar] [CrossRef]
- He, M.; Li, X.; Poolsawat, L.; Guo, Z.; Yao, W.; Zhang, C.; Leng, X. Effects of fish meal replaced by fermented soybean meal on growth performance, intestinal histology and microbiota of largemouth bass (Micropterus salmoides). Aquac. Nutr. 2020, 26, 1058–1071. [Google Scholar] [CrossRef]
- Yang, H.; Bian, Y.; Huang, L.; Lan, Q.; Ma, L.; Li, X.; Leng, X. Effects of replacing fish meal with fermented soybean meal on the growth performance, intestinal microbiota, morphology and disease resistance of largemouth bass (Micropterus salmoides). Aquac. Rep. 2021, 22, 100954. [Google Scholar] [CrossRef]
- Huizinga, H.W.; Esch, G.; Hazen, T.C. Histopathology of red-sore disease (Aeromonas hydrophila) in naturally and experimentally infected largemouth bass Micropterus salmoides (Lacepede). J. Fish Dis. 1979, 2, 263–277. [Google Scholar] [CrossRef]
- Pei, C.; Song, H.; Zhu, L.; Qiao, D.; Yan, Y.; Li, L.; Zhao, X.; Zhang, J.; Jiang, X.; Kong, X. Identification of Aeromonas veronii isolated from largemouth bass Micropterus salmoides and histopathological analysis. Aquaculture 2021, 540, 736707. [Google Scholar] [CrossRef]
- Yuan, X.Y.; Zhang, X.T.; Xia, Y.T.; Zhang, Y.Q.; Wang, B.; Ye, W.W.; Ye, Z.F.; Qian, S.C.; Huang, M.M.; Yang, S.; et al. Transcriptome and 16S rRNA analyses revealed differences in the responses of largemouth bass (Micropterus salmoides) to early Aeromonas hydrophila infection and immunization. Aquaculture 2021, 541, 736759. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, H.; Zhang, L.; Li, Q.; Huang, Z.; Zhao, Z.; Ke, H.; Xiao, Y.; Su, X.; Liu, Q.; et al. MiRNA-seq analysis of spleen and head kidney tissue from aquacultured largemouth bass (Micropterus salmoides) in response to Aeromonas hydrophila infection. Funct. Integr. Genom. 2021, 21, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Liu, G.; Yao, T.; Lu, J. Monitoring of antimicrobial resistance genes in the spotted sea bass (Lateolabrax maculatus): Association with the microbiome and its environment in aquaculture ponds. Environ. Pollut. 2021, 276, 116714. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.; Tan, K.; Gong, P.; Lei, P.; Guo, Z.; Wang, S.; Gao, S.; Zhou, Y.; Shu, Y.; Zhou, X.; et al. Correlation of microbiota in the gut of fish species and water. 3 Biotech 2020, 10, 472. [Google Scholar] [CrossRef] [PubMed]
- Bartelme, R.P.; Smith, M.C.; Sepulveda-Villet, O.J.; Newton, R.J. Component Microenvironments and System Biogeography Structure Microorganism Distributions in Recirculating Aquaculture and Aquaponic Systems. mSphere 2019, 4, e00143-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Schryver, P.; Vadstein, O. Ecological theory as a foundation to control pathogenic invasion in aquaculture. ISME J. 2014, 8, 2360–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attramadal, K.J.; Truong, T.M.H.; Bakke, I.; Skjermo, J.; Olsen, Y.; Vadstein, O. RAS and microbial maturation as tools for K-selection of microbial communities improve survival in cod larvae. Aquaculture 2014, 432, 483–490. [Google Scholar] [CrossRef]
- Bakke, I.; Åm, A.L.; Kolarevic, J.; Ytrestøyl, T.; Vadstein, O.; Attramadal, K.J.K.; Terjesen, B.F. Microbial community dynamics in semi-commercial RAS for production of Atlantic salmon post-smolts at different salinities. Aquac. Eng. 2017, 78, 42–49. [Google Scholar] [CrossRef]
- Liu, H.; Shiver, A.L.; Price, M.N.; Carlson, H.K.; Trotter, V.V.; Chen, Y.; Escalante, V.; Ray, J.; Hern, K.E.; Petzold, C.J.; et al. Functional genetics of human gut commensal Bacteroides thetaiotaomicron reveals metabolic requirements for growth across environments. Cell Rep. 2021, 34, 108789. [Google Scholar] [CrossRef]
- Meehan, C.; Beiko, R.G. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Miquel, S.; Martin, R.; Rossi, O.; Bermudez-Humaran, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, J.; Zhang, H.; Lee, Y.-K.; Zhai, Q.; Chen, W. Roles of intestinal bacteroides in human health and diseases. Crit. Rev. Food Sci. Nutr. 2020, 61, 3518–3536. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martín, R.; Bridonneau, C.; Robert, V.; Sokol, H.; Bermudez-Humaran, L.G.; Thomas, M.; Langella, P. Ecology and metabolism of the beneficial intestinal commensal bacteriumFaecalibacterium prausnitzii. Gut Microbes 2014, 5, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Bai, X.; Li, Y.; Jing, L.; Chen, R.; Teng, Y. Characterization and source-tracking of antibiotic resistomes in the sediments of a peri-urban river. Sci. Total Environ. 2019, 679, 88–96. [Google Scholar] [CrossRef]
- Duan, Y.; Xiong, D.; Li, Y.; Ding, X.; Dong, H.; Wang, W.; Zhang, J. Changes in the microbial communities of the rearing water, sediment and gastrointestinal tract of Lateolabrax maculatus at two growth stages. Aquac. Rep. 2021, 20, 100742. [Google Scholar] [CrossRef]
- Jiang, S.C.; Han, M.; Chandrasekaran, S.; Fang, Y.; Kellogg, C.A. Assessing the water quality impacts of two Category-5 hurricanes on St. Thomas, Virgin Islands. Water Res. 2019, 171, 115440. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; You, Y.; Peng, F.; Tang, X.; Zhou, Y.; Liu, J.; Lin, D.; Zhou, Y. Interaction of Microbiota between Fish and the Environment of an In-Pond Raceway System in a Lake. Microorganisms 2022, 10, 1143. https://doi.org/10.3390/microorganisms10061143
Zhang X, You Y, Peng F, Tang X, Zhou Y, Liu J, Lin D, Zhou Y. Interaction of Microbiota between Fish and the Environment of an In-Pond Raceway System in a Lake. Microorganisms. 2022; 10(6):1143. https://doi.org/10.3390/microorganisms10061143
Chicago/Turabian StyleZhang, Xizhao, Yang You, Fei Peng, Xuemei Tang, Yifan Zhou, Jianyu Liu, Danqing Lin, and Yanfeng Zhou. 2022. "Interaction of Microbiota between Fish and the Environment of an In-Pond Raceway System in a Lake" Microorganisms 10, no. 6: 1143. https://doi.org/10.3390/microorganisms10061143