
Salt- and pH-Dependent Thermal Stability of Photocomplexes from Extremophilic Bacteriochlorophyll b-Containing Halorhodospira Species

 and
and 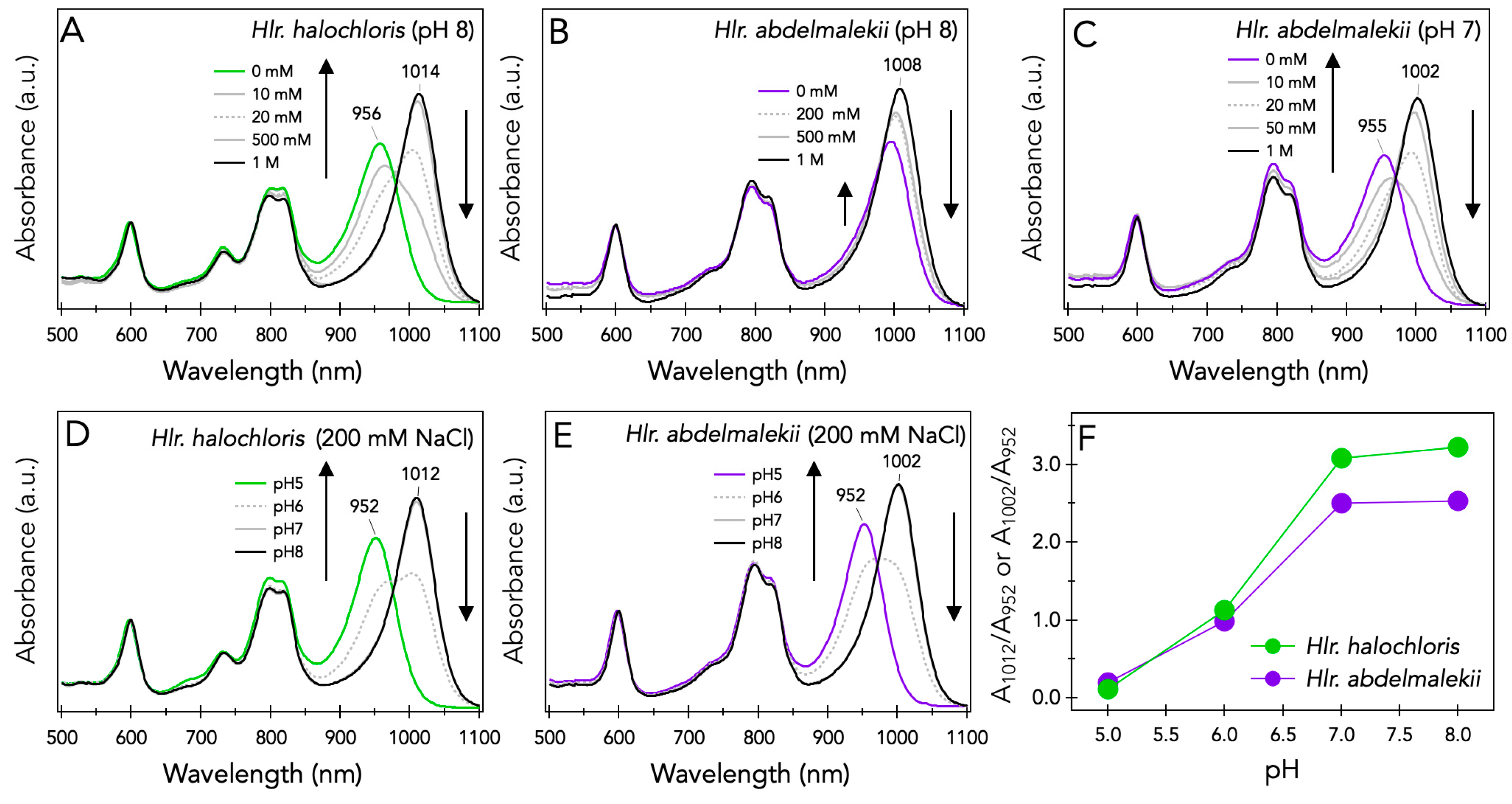{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Preparation of LH1-RC Complexes
2.2. Evaluation of Thermal Stability
3. Results
3.1. NaCl and pH Effects on Spectroscopic Properties of LH1-RC Complexes from Halorhodospira Species
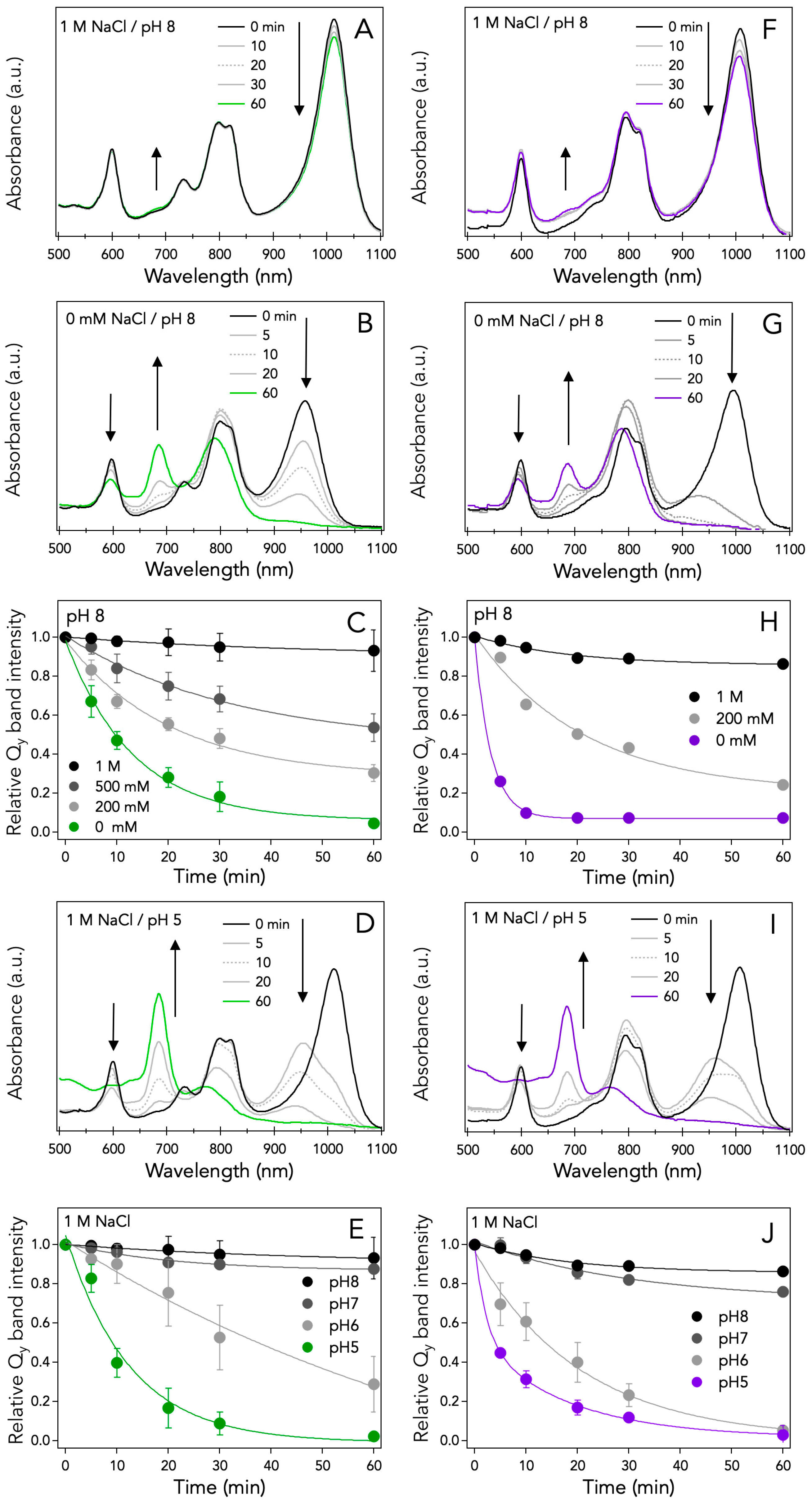
3.2. NaCl and pH Effects on the Thermal Stability of LH1-RC Complexes from Halorhodospira Species
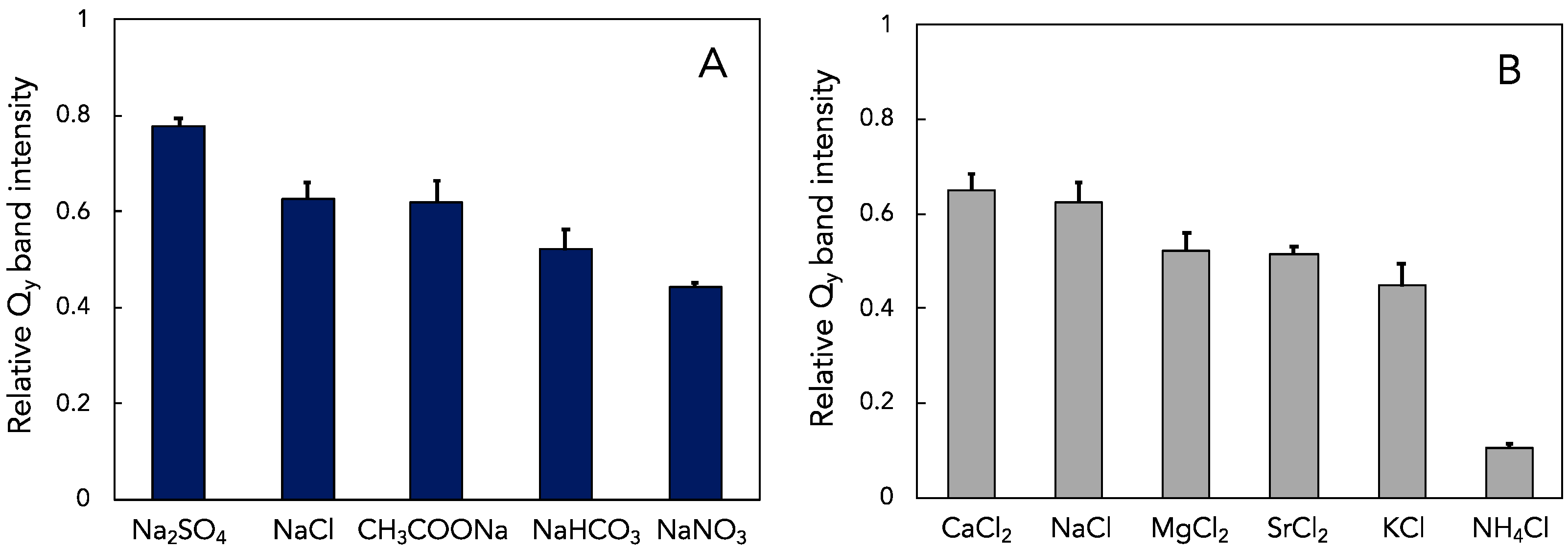
3.3. Effect of Other Salts on Thermal Stability of LH1-RCs from Hlr. halochloris
4. Discussion
4.1. Effects of Salt Concentration and pH on the Spectral Properties of the LH1-RC Complexes from Halorhodospira Species
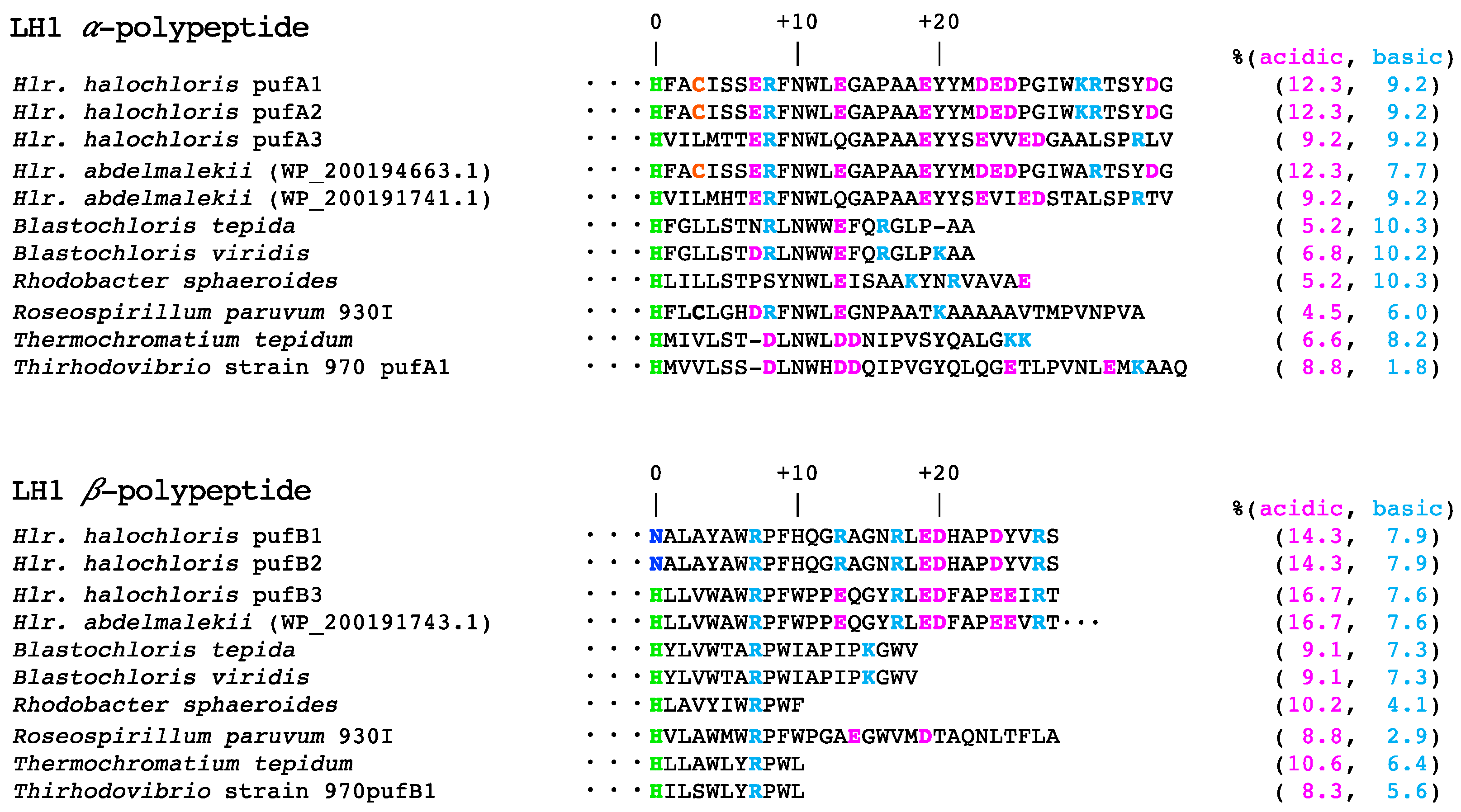
4.2. Effects of Amino Acid Compositions on the Thermal Stability of the LH1-RC Complexes from Halorhodospira Species
4.3. Effects of Anions and Cations on the Thermal Stability of the LH1-RC Complexes from Halorhodospira Species
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Madigan, M.T. Anoxygenic phototrophic bacteria from extreme environments. Photosynth. Res. 2003, 76, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F.; Rahn, T.; Kunzel, S.; Neulinger, S.C. Photosynthesis Is widely distributed among proteobacteria as demonstrated by the phylogeny of pufLM reaction center proteins. Front. Microbiol. 2018, 8, 2679. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Trüper, H.G. Ectothiorhodospira halochloris sp. nov., s new extremely halophilic phototropic bacterium containing bacteriochlorophyll b. Arch. Microbiol. 1977, 114, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F.; Suling, J. The phylogenetic relationship among Ectothiorhodospiraceae: A reevaluation of their taxonomy on the basis of 16S rDNA analyses. Arch. Microbiol. 1996, 165, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F.; Trüper, H.G. Ectothiorhodospira abdelmalekii sp. nov. a new halophilic and alkaliphilic phototropic bacterium. Zent. Bakteriol. I Abt. Orig. C Allg. Angew. Okol. Mikrobiol. 1981, 2, 228–234. [Google Scholar] [CrossRef]
- Drews, G.; Giesbrecht, P. Rhodopseudomonas viridis, nov. spec., ein neu isoliertes, obligat phototrophes bakterium. Arch. Mikrobiol. 1966, 53, 255–262. [Google Scholar] [CrossRef]
- Eimhjellen, K.E. Thiocapsa pfennigii sp. nov. a new species of the phototrophic sulfur bacteria. Arch. Mikrobiol. 1970, 73, 193–194. [Google Scholar] [CrossRef]
- Keppen, O.I.; Gorlenko, V.M. New species of purple budding bacteria containing bacteriochlorophyll-b. Microbiol. 1975, 44, 224–229. [Google Scholar]
- Resnick, S.M.; Madigan, M.T. Isolation and characterization of a mildly thermophilic nonsulfur purple bacterium containing bacteriochlorophyll-b. FEMS Microbiol. Lett. 1989, 65, 165–170. [Google Scholar] [CrossRef]
- Hiraishi, A. Transfer of the bacteriochlorophyll b-containing phototrophic bacteria Rhodopseudomonas viridis and Rhodopseudomonas sulfoviridis to the genus Blastochloris gen. nov. Int. J. Syst. Bacteriol. 1997, 47, 217–219. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.T.; Resnick, S.M.; Kempher, M.L.; Dohnalkova, A.C.; Takaichi, S.; Wang-Otomo, Z.Y.; Toyoda, A.; Kurokawa, K.; Mori, H.; Tsukatani, Y. Blastochloris tepida, sp. nov., a thermophilic species of the bacteriochlorophyll b-containing genus Blastochloris. Arch. Microbiol. 2019, 201, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Bryantseva, I.A.; Gorlenko, V.M.; Kompantseva, E.I.; Imhoff, J.F. Thioalkalicoccus limnaeus gen. nov., sp nov., a new alkaliphilic purple sulfur bacterium with bacteriochlorophyll b. Int. J. Syst. Evol. Micr. 2000, 50, 2157–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogdell, R.J.; Gall, A.; Köhler, J. The architecture and function of the light-harvesting apparatus of purple bacteria: From single molecules to in vivo membranes. Q. Rev. Biophys. 2006, 39, 227–324. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.T. A novel photosynthetic purple bacterium isolated from a Yellowstone hot spring. Science 1984, 225, 313–315. [Google Scholar] [CrossRef]
- Madigan, M.T.; Absher, J.N.; Mayers, J.E.; Asao, M.; Jung, D.O.; Bender, K.S.; Kempher, M.L.; Hayward, M.K.; Sanguedolce, S.A.; Brown, A.C.; et al. Allochromatium tepidum, sp. nov., a hot spring species of purple sulfur bacteria. Arch. Microbiol. 2022, 204, 115. [Google Scholar] [CrossRef] [PubMed]
- Castenholz, R.W.; Bauld, J.; Jorgenson, B.B. Anoxygenic microbial mats of hot springs—Thermophilic Chlorobium sp. Fems. Microbiol. Ecol. 1990, 74, 325–336. [Google Scholar] [CrossRef]
- Saini, M.K.; ChihChe, W.; Soulier, N.; Sebastian, A.; Albert, I.; Thiel, V.; Bryant, D.A.; Hanada, S.; Tank, M. Caldichromatium japonicum gen. nov., sp. nov., a novel thermophilic phototrophic purple sulphur bacterium of the Chromatiaceae isolated from Nakabusa hot springs, Japan. Int. J. Syst. Evol. Micr. 2020, 70, 5701–5710. [Google Scholar] [CrossRef]
- Raymond, J.C.; Sistrom, W.R. Ectothiorhodospira Halophila—A New Species of Genus Ectothiorhodospira. Arch. Mikrobiol. 1969, 69, 121–126. [Google Scholar] [CrossRef]
- Kimura, Y.; Yu, L.J.; Hirano, Y.; Suzuki, H.; Wang, Z.Y. Calcium ions are required for the enhanced thermal stability of the light-harvesting-reaction center core complex from thermophilic purple sulfur bacterium Thermochromatium tepidum. J. Biol. Chem. 2009, 284, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Yura, Y.; Hayashi, Y.; Li, Y.; Onoda, M.; Yu, L.-J.; Wang-Otomo, Z.-Y.; Ohno, T. Spectroscopic and thermodynamic characterization of the metal-binding sites in the LH1–RC complex from thermophilic photosynthetic bacterium Thermochromatium tepidum. J. Phys. Chem. B 2016, 120, 12466–12473. [Google Scholar] [CrossRef]
- Yu, L.J.; Suga, M.; Wang-Otomo, Z.Y.; Shen, J.R. Structure of photosynthetic LH1-RC supercomplex at 1.9 Å resolution. Nature 2018, 556, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Lyu, S.; Okoshi, A.; Okazaki, K.; Nakamura, N.; Ohashi, A.; Ohno, T.; Kobayashi, M.; Imanishi, M.; Takaichi, S.; et al. Effects of calcium ions on the thermostability and spectroscopic properties of the LH1-RC complex from a new thermophilic purple bacterium Allochromatium tepidum. J. Phys. Chem. B 2017, 121, 5025–5032. [Google Scholar] [CrossRef] [PubMed]
- Tani, K.; Kobayashi, K.; Hosogi, N.; Ji, X.-C.; Nagashima, S.; Nagashima, K.V.P.; Tsukatani, Y.; Kanno, R.; Hall, M.; Yu, L.-J.; et al. A Ca2+-binding motif underlies the unusual properties of certain photosynthetic bacterial core light-harvesting complexes. J. Biol. Chem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Seto, R.; Takaichi, S.; Kurihara, T.; Kishi, R.; Honda, M.; Takenaka, S.; Tsukatani, Y.; Madigan, M.T.; Wang-Otomo, Z.Y.; Kimura, Y. Lycopene-family carotenoids confer thermostability on photocomplexes from a new thermophilic purple bacterium. Biochemistry 2020, 59, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Nojima, S.; Nakata, K.; Yamashita, T.; Wang, X.P.; Takenaka, S.; Akimoto, S.; Kobayashi, M.; Madigan, M.T.; Wang-Otomo, Z.Y.; et al. Electrostatic charge controls the lowest LH1 Qy transition energy in the triply extremophilic purple phototrophic bacterium, Halorhodospira halochloris. Biochim. Biophys. Acta 2021, 1862, 148473. [Google Scholar] [CrossRef]
- Steiner, R.; Scheer, H. Characterization of a B800-1020 antenna from the photosynthetic bacteria Ectothiorhodospira halochloris and Ectothiorhodospira abdelmalekii. Biochim. Biophys. Acta 1985, 807, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Hirano, Y.; Yu, L.J.; Suzuki, H.; Kobayashi, M.; Wang, Z.Y. Calcium ions are involved in the unusual red shift of the light-harvesting 1 Qy transition of the core complex in thermophilic purple sulfur bacterium Thermochromatium tepidum. J. Biol. Chem. 2008, 283, 13867–13873. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, M.; Takenouchi, M.; Takaichi, S.; Nakagawa, S.; Saga, Y.; Takenaka, S.; Madigan, M.T.; Overmann, J.; Wang-Otomo, Z.Y.; Kimura, Y. A dual role for Ca2+ in expanding the spectral diversity and stability of light-harvesting 1 reaction center photocomplexes of purple phototrophic bacteria. Biochemistry 2019, 58, 2844–2852. [Google Scholar] [CrossRef]
- Hofmeister, F. Zur Lehre von der Wirkung der Salze. Arch. Exp. Pathol. Pharmacol. 1888, 24, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Hofmeister effects: An explanation for the impact of ionic liquids on biocatalysis. J. Biotechnol. 2009, 144, 12–22. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Cremer, P.S. Interactions between macromolecules and ions: The Hofmeister series. Curr. Opin. Chem. Biol. 2006, 10, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Wagnerhuber, R.; Brunisholz, R.A.; Bissig, I.; Frank, G.; Zuber, H. A new possible binding-site for bacteriochlorophyll-b in a light-harvesting polypeptide of the bacterium Ectothiorhodospira halochloris. FEBS Lett. 1988, 233, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Tuschak, C.; Beatty, J.T.; Overmann, J. Photosynthesis genes and LH1 proteins of Roseospirillum parvum 930I, a purple non-sulfur bacterium with unusual spectral properties. Photosynth. Res. 2004, 81, 181–199. [Google Scholar] [CrossRef]
- Tsukatani, Y.; Hirose, Y.; Harada, J.; Yonekawa, C.; Tamiaki, H. Unusual features in the photosynthetic machinery of Halorhodospira halochloris DSM 1059 revealed by complete genome sequencing. Photosynth. Res. 2019, 140, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Awoonor-Williams, E.; Rowley, C.N. Evaluation of methods for the calculation of the pka of cysteine residues in proteins. J. Chem. Theory. Comput. 2016, 12, 4662–4673. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.; Siebert, C.A.; Wang, P.Y.; Canniffe, D.P.; Hunter, C.N. Cryo-EM structure of the Blastochloris viridis LH1-RC complex at 2.9 Å. Nature 2018, 556, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Wagnerhuber, R.; Brunisholz, R.A.; Bissig, I.; Frank, G.; Suter, F.; Zuber, H. The primary structure of the antenna polypeptides of Ectothiorhodospira halochloris and Ectothiorhodospira halophila—Four core-type antenna polypeptides in E. halochloris and E. halophila. Eur. J. Biochem. 1992, 205, 917–925. [Google Scholar] [CrossRef]
- Mevarech, M.; Frolow, F.; Gloss, L.M. Halophilic enzymes: Proteins with a grain of salt. Biophys. Chem. 2000, 86, 155–164. [Google Scholar] [CrossRef]
- Roberts, M.F. Organic compatible solutes of halotolerant and halophilic microorganisms. Saline Syst. 2005, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Galinski, E.A.; Trüper, H.G. Betaine, a Compatible Solute in the Extremely Halophilic Phototropic Bacterium Ectothiorhodospira halochloris. FEMS Microbiol. Lett. 1982, 13, 357–360. [Google Scholar] [CrossRef]
- Galinski, E.A.; Pfeiffer, H.P.; Trüper, H.G. 1,4,5,6-Tetrahydro-2-Methyl-4-Pyrimidinecarboxylic Acid—A Novel Cyclic Amino-Acid from Halophilic Phototrophic Bacteria of the Genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Life at high salt concentrations, intracellular KCl concentrations, and acidic proteomes. Front. Microbiol. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deole, R.; Challacombe, J.; Raiford, D.W.; Hoff, W.D. An Extremely Halophilic Proteobacterium Combines a Highly Acidic Proteome with a Low Cytoplasmic Potassium Content. J. Biol. Chem. 2013, 288, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanyi, J.K. Salt-Dependent Properties of Proteins from Extremely Halophilic Bacteria. Bacteriol. Rev. 1974, 38, 272–290. [Google Scholar] [CrossRef] [PubMed]
- Elcock, A.H.; McCammon, J.A. Electrostatic contributions to the stability of halophilic proteins. J. Mol. Biol. 1998, 280, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Fukuchi, S.; Yoshimune, K.; Wakayama, M.; Moriguchi, M.; Nishikawa, K. Unique amino acid composition of proteins in halophilic bacteria. J. Mol. Biol. 2003, 327, 347–357. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P. Halophiles and their enzymes: Negativity put to good use. Curr. Opin. Microbiol. 2015, 25, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Fukuchi, S.; Nishikawa, K. Protein surface amino acid compositions distinctively differ between thermophilic and mesophilic bacteria. J. Mol. Biol. 2001, 309, 835–843. [Google Scholar] [CrossRef]
- Richard, S.B.; Madern, D.; Garcin, E.; Zaccai, G. Halophilic adaptation: Novel solvent protein interactions observed in the 2.9 and 2.6 a resolution structures of the wild type and a mutant of malate dehydrogenase from Haloarcula marismortui. Biochemistry 2000, 39, 992–1000. [Google Scholar] [CrossRef]
- Britton, K.L.; Baker, P.J.; Fisher, M.; Ruzheinikov, S.; Gilmour, D.J.; Bonete, M.J.; Ferrer, J.; Pire, C.; Esclapez, J.; Rice, D.W. Analysis of protein solvent interactions in glucose dehydrogenase from the extreme halophile Haloferax mediterranei. Proc. Natl. Acad. Sci. USA 2006, 103, 4846–4851. [Google Scholar] [CrossRef] [Green Version]
- Okur, H.I.; Hladilkova, J.; Rembert, K.B.; Cho, Y.; Heyda, J.; Dzubiella, J.; Cremer, P.S.; Jungwirth, P. Beyond the Hofmeister Series: Ion-Specific Effects on Proteins and Their Biological Functions. J. Phys. Chem. B 2017, 121, 1997–2014. [Google Scholar] [CrossRef] [PubMed]
- Tani, K.; Kanno, R.; Makino, Y.; Hall, M.; Takenouchi, M.; Imanishi, M.; Yu, L.J.; Overmann, J.; Madigan, M.T.; Kimura, Y.; et al. Cryo-EM structure of a Ca2+-bound photosynthetic LH1-RC complex containing multiple αβ-polypeptides. Nat. Commun. 2020, 11, 4955. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimura, Y.; Nakata, K.; Nojima, S.; Takenaka, S.; Madigan, M.T.; Wang-Otomo, Z.-Y. Salt- and pH-Dependent Thermal Stability of Photocomplexes from Extremophilic Bacteriochlorophyll b-Containing Halorhodospira Species. Microorganisms 2022, 10, 959. https://doi.org/10.3390/microorganisms10050959
Kimura Y, Nakata K, Nojima S, Takenaka S, Madigan MT, Wang-Otomo Z-Y. Salt- and pH-Dependent Thermal Stability of Photocomplexes from Extremophilic Bacteriochlorophyll b-Containing Halorhodospira Species. Microorganisms. 2022; 10(5):959. https://doi.org/10.3390/microorganisms10050959
Chicago/Turabian StyleKimura, Yukihiro, Kazuna Nakata, Shingo Nojima, Shinji Takenaka, Michael T. Madigan, and Zheng-Yu Wang-Otomo. 2022. "Salt- and pH-Dependent Thermal Stability of Photocomplexes from Extremophilic Bacteriochlorophyll b-Containing Halorhodospira Species" Microorganisms 10, no. 5: 959. https://doi.org/10.3390/microorganisms10050959