
Comprehensive ADM1 Extensions to Tackle Some Operational and Metabolic Aspects in Anaerobic Digestion



Abstract
:1. Introduction
2. The Rationale Behind
2.1. Operational Aspects
2.1.1. Feeding Strategy
2.1.2. Working Volume Reduction
2.2. Metabolic Aspects
2.2.1. Extra- Intra- Cellular Transport
2.2.2. Adaptation
3. Methodology
3.1. Model Extensions
3.1.1. Feeding Strategy
3.1.2. Working Volume Reduction
3.1.3. Extra- Intra- Cellular Transport
3.1.4. Adaptation
3.2. Modelling Conditions and Simulation
4. Results
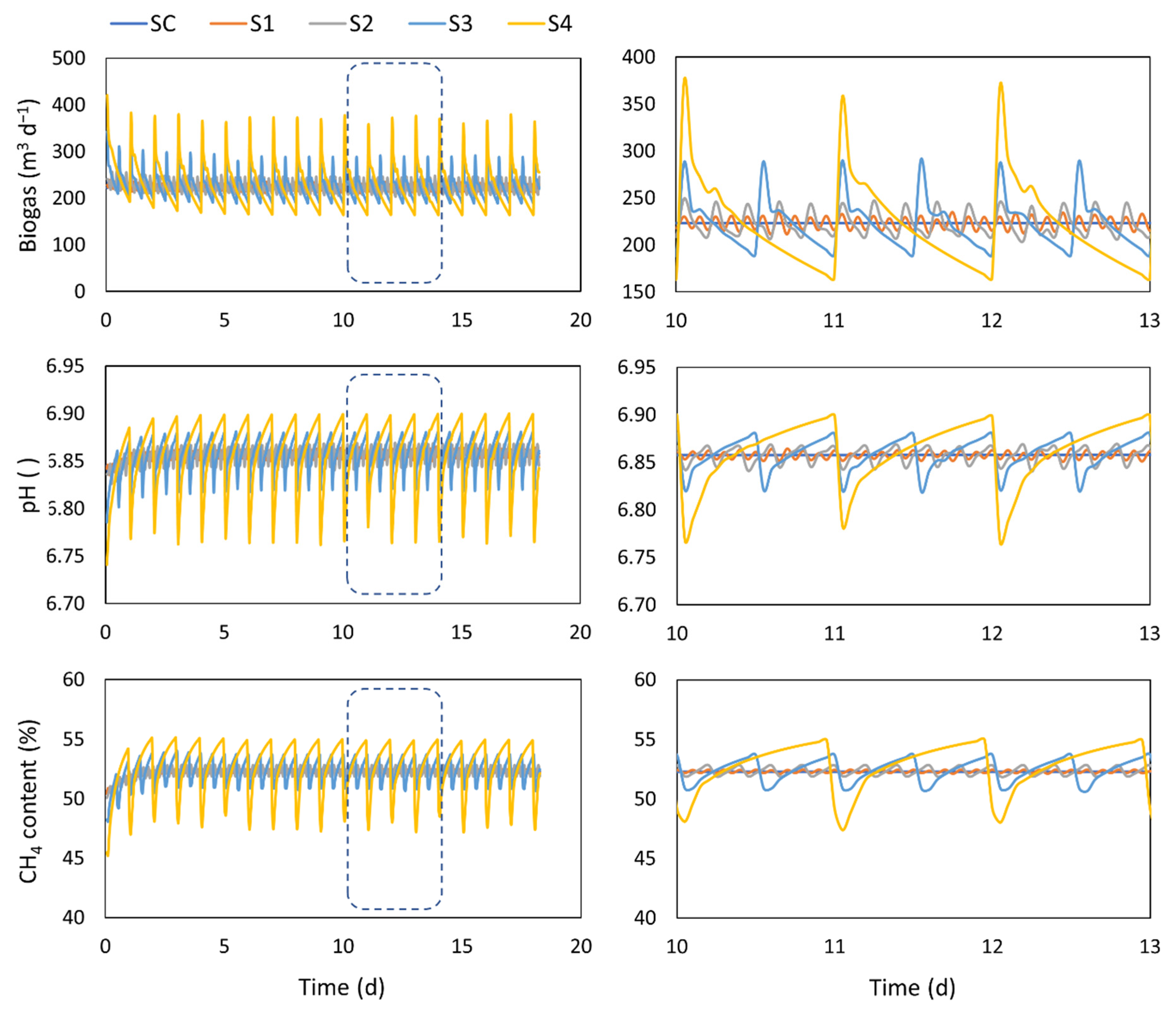
4.1. Feeding Strategy
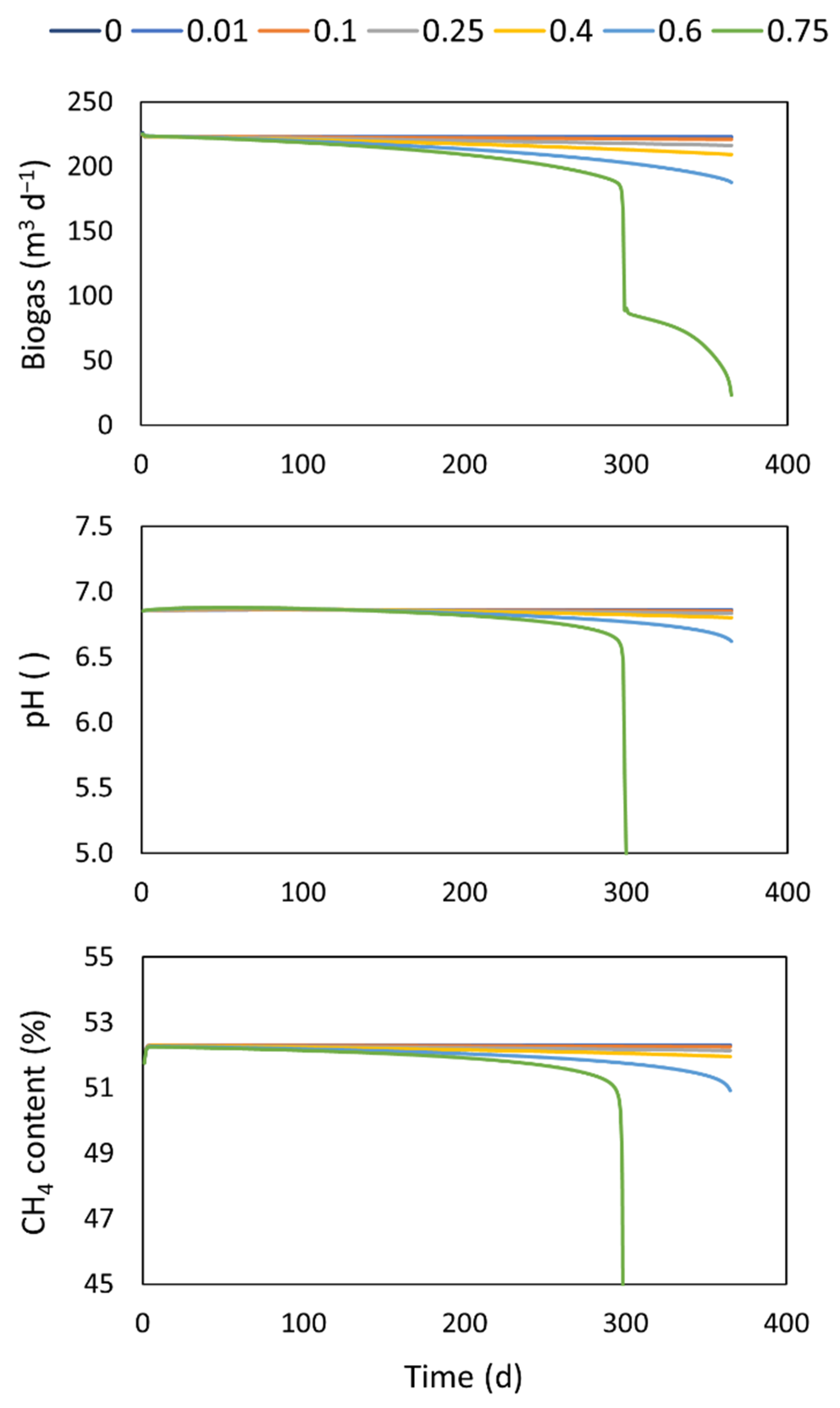
4.2. Working Volume Reduction
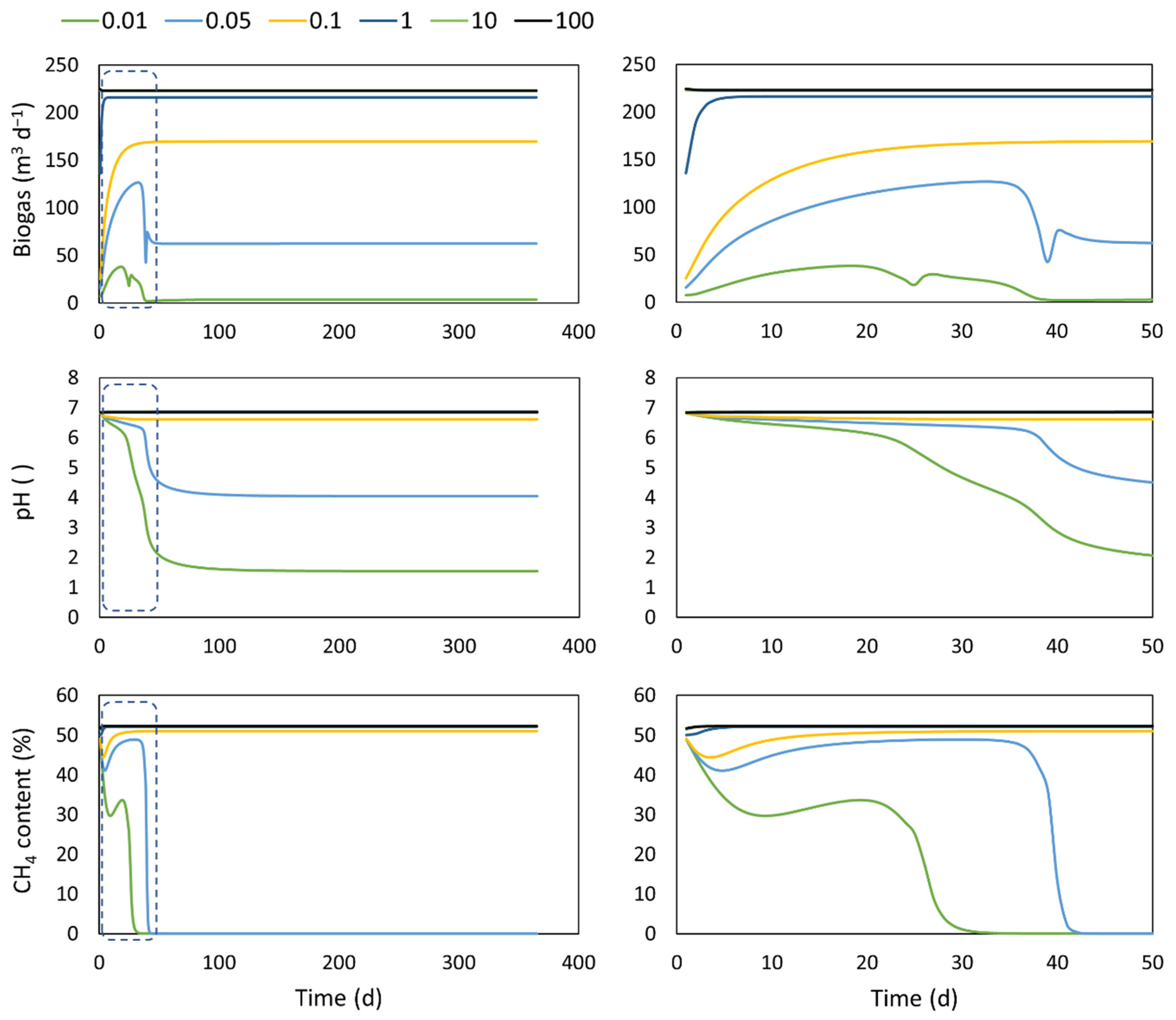
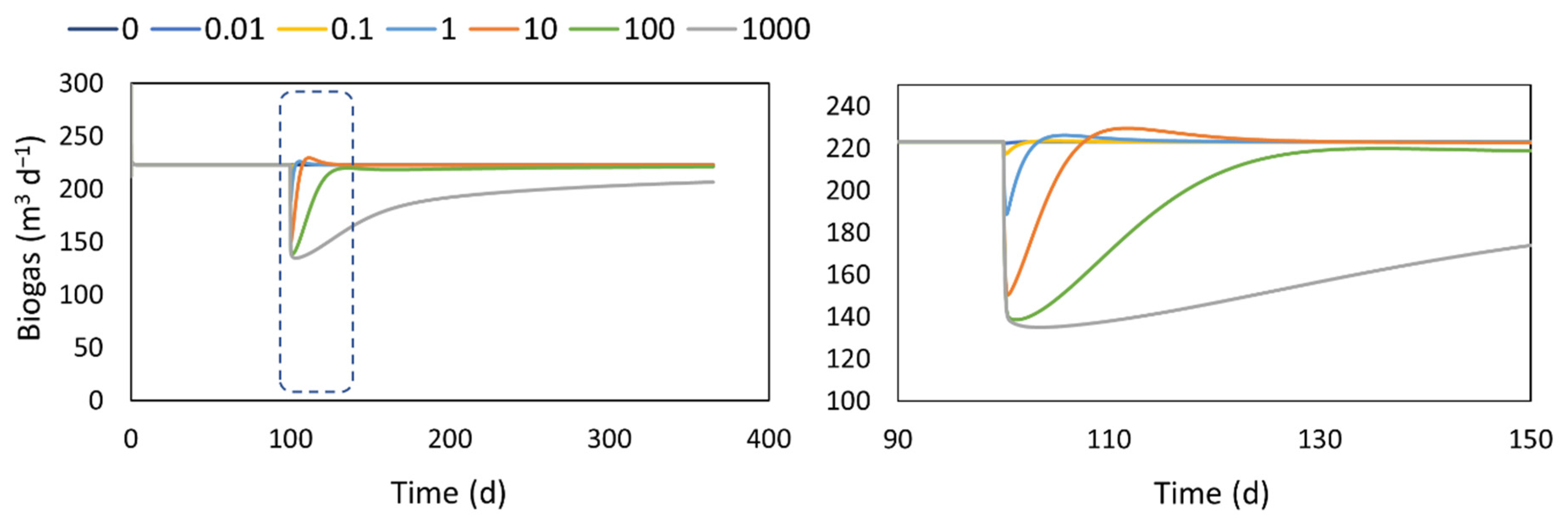
4.3. Extra- Intra- Cellular Transport
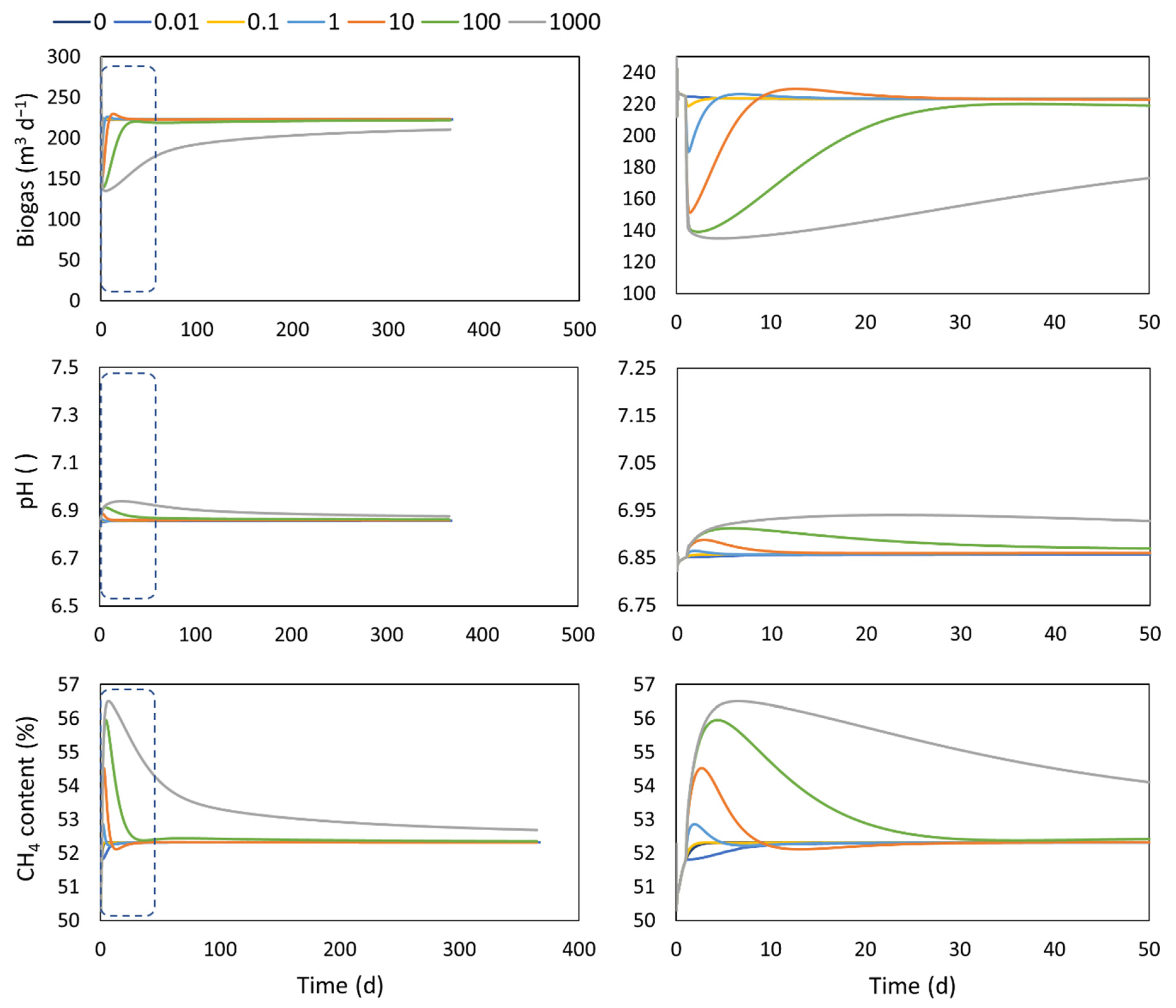
4.4. Adaptation
4.5. On Model Extension Validation
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fagerström, A.; Al Seadi, T.; Rasi, S.; Briseid, T. The Role of Anaerobic Digestion and Biogas in the Circular Economy; IEA Bioenergy: Cork, Ireland, 2018; Volume 8. [Google Scholar]
- Batstone, D.J.; Keller, J.; Angelidaki, I.; Kalyuzhnyi, S.V.; Pavlostathis, S.G.; Rozzi, A.; Sanders, W.T.M.; Siegrist, H.; Vavilin, V.A. The IWA Anaerobic Digestion Model No 1 (ADM1). Water Sci. Technol. 2002, 45, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Hinken, L.; Huber, M.; Weichgrebe, D.; Rosenwinkel, K.H. Modified ADM1 for modelling an UASB reactor laboratory plant treating starch wastewater and synthetic substrate load tests. Water Res. 2014, 64, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Barrera, E.L.; Spanjers, H.; Solon, K.; Amerlinck, Y.; Nopens, I.; Dewulf, J. Modeling the anaerobic digestion of cane-molasses vinasse: Extension of the Anaerobic Digestion Model No. 1 (ADM1) with sulfate reduction for a very high strength and sulfate rich wastewater. Water Res. 2015, 71, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Fedorovich, V.; Lens, P.; Kalyuzhnyi, S. Extension of Anaerobic Digestion Model No. 1 with Process of Sulfate Reduction. Appl. Biochem. Biotechnol. 2003, 109, 33–45. [Google Scholar] [CrossRef]
- Batstone, D.J.; Torrijos, M.; Ruiz, C.; Schmidt, J.E. Use of an anaerobic sequencing batch reactor for parameter estimation in modelling of anaerobic digestion. Water Sci. Technol. 2004, 50, 295–303. [Google Scholar] [CrossRef]
- Zaher, U.; Moussa, M.S.; Widyatmika, I.N.; van Der Steen, P.; Gijzen, H.J. Modelling anaerobic digestion acclimatisation to a biodegradable toxicant: Application to cyanide. Water Sci. Technol. 2006, 54, 129–137. [Google Scholar] [CrossRef]
- Wade, M.J. Not Just Numbers: Mathematical Modelling and Its Contribution to Anaerobic Digestion Processes. Processes 2020, 8, 888. [Google Scholar] [CrossRef]
- Weinrich, S.; Nelles, M. Basics of Anaerobic Digestion-Biochemical Conversion and Process Modelling. DBFZ-Report 2021, 40, 9–76. [Google Scholar]
- Catenacci, A.; Grana, M.; Malpei, F.; Ficara, E. Optimizing ADM1 Calibration and Input Characterization for Effective Co-Digestion Modelling. Water 2021, 13, 3100. [Google Scholar] [CrossRef]
- Weinrich, S.; Koch, S.; Bonk, F.; Popp, D.; Benndorf, D.; Klamt, S.; Centler, F. Augmenting Biogas Process Modeling by Resolving Intracellular Metabolic Activity. Front. Microbiol. 2019, 10, 1095. [Google Scholar] [CrossRef]
- Donoso-Bravo, A.; Sadino-Riquelme, C.; Gómez, D.; Segura, C.; Valdebenito, E.; Hansen, F. Modelling of an anaerobic plug-flow reactor. Process analysis and evaluation approaches with non-ideal mixing considerations. Bioresour. Technol. 2018, 260, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Keshtkar, A.; Meyssami, B.; Abolhamd, G.; Ghaforian, H.; Asadi, M.K. Mathematical modeling of non-ideal mixing continuous flow reactors for anaerobic digestion of cattle manure. Bioresour. Technol. 2003, 87, 113–124. [Google Scholar] [CrossRef]
- Sadino-Riquelme, C.; Hayes, R.E.; Jeison, D.; Donoso-Bravo, A. Computational fluid dynamic (CFD) modelling in anaerobic digestion: General application and recent advances. Crit. Rev. Environ. Sci. Technol. 2018, 48, 39–76. [Google Scholar] [CrossRef]
- Batstone, D.J.; Puyol, D.; Flores-Alsina, X.; Rodríguez, J. Mathematical modelling of anaerobic digestion processes: Applications and future needs. Rev. Environ. Sci. Biotechnol. 2015, 14, 595–613. [Google Scholar] [CrossRef]
- Drosg, B. Process Monitoring in Biogas Plants; IEA Bioenergy: Paris, France, 2013. [Google Scholar]
- Wang, M.; Sun, X.; Li, P.; Yin, L.; Liu, D.; Zhang, Y.; Li, W.; Zheng, G. A novel alternate feeding mode for semi-continuous anaerobic co-digestion of food waste with chicken manure. Bioresour. Technol. 2014, 164, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Mairet, F.; Bernard, O.; Cameron, E.; Ras, M.; Lardon, L.; Steyer, J.P.; Chachuat, B. Three-reaction model for the anaerobic digestion of microalgae. Biotechnol. Bioeng. 2012, 109, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AgSTAR Program. Anaerobic Digester/Biogas System Operator Guidebook; EPA: Washington, DC, USA, 2020. [Google Scholar]
- Arnott, M. The Biogas/Biofertilizer Business Handbook; Peace Corps, Information Collection and Exchange: Washington, DC, USA, 1984. [Google Scholar]
- Flores-Alsina, X.; Solon, K.; Mbamba, C.K.; Tait, S.; Gernaey, K.V.; Jeppsson, U.; Batstone, D.J. Modelling phosphorus (P), sulfur (S) and iron (Fe) interactions for dynamic simulations of anaerobic digestion processes. Water Res. 2016, 95, 370–382. [Google Scholar] [CrossRef] [Green Version]
- Mbamba, C.K.; Flores-Alsina, X.; Batstone, D.J.; Tait, S. Validation of a plant-wide phosphorus modelling approach with minerals precipitation in a full-scale WWTP. Water Res. 2016, 100, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga, H.; Vergara, C.; Donoso-Bravo, A.; Jeison, D. Effect of delays on the response of microalgae when exposed to dynamic environmental conditions. Processes 2020, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.S.; Rosenthal, K.S. Microbial adaptation: Putting the best team on the field. Infect. Dis. Clin. Pract. 2007, 15, 330–334. [Google Scholar] [CrossRef]
- Wintsche, B.; Jehmlich, N.; Popp, D.; Harms, H.; Kleinsteuber, S. Metabolic adaptation of methanogens in anaerobic digesters upon trace element limitation. Front. Microbiol. 2018, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Wojcieszak, M.; Pyzik, A.; Poszytek, K.; Krawczyk, P.S.; Sobczak, A.; Lipinski, L.; Roubinek, O.; Palige, J.; Sklodowska, A.; Drewniak, L. Adaptation of methanogenic inocula to anaerobic digestion of maize silage. Front. Microbiol. 2017, 8, 1881. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.S.; Buezo, K.A.; Saady, N.M.C. Adapting anaerobic consortium to pure and complex lignocellulose substrates at low temperature: Kinetics evaluation. Int. J. Recycl. Org. Waste Agric. 2019, 8, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Kalamaras, S.D.; Vasileiadis, S.; Karas, P.; Angelidaki, I.; Kotsopoulos, T.A. Microbial adaptation to high ammonia concentrations during anaerobic digestion of manure-based feedstock: Biomethanation and 16S rRNA gene sequencing. J. Chem. Technol. Biotechnol. 2020, 95, 1970–1979. [Google Scholar] [CrossRef]
- Agyeman, F.O.; Han, Y.; Tao, W. Elucidating the kinetics of ammonia inhibition to anaerobic digestion through extended batch experiments and stimulation-inhibition modeling. Bioresour. Technol. 2021, 340, 125744. [Google Scholar] [CrossRef]
- Kovalovszki, A.; Treu, L.; Ellegaard, L.; Luo, G.; Angelidaki, I. Modeling temperature response in bioenergy production: Novel solution to a common challenge of anaerobic digestion. Appl. Energy 2020, 263, 114646. [Google Scholar] [CrossRef]
- Rosen, C.; Jeppsson, U. Aspects on ADM1 Implementation within the BSM2 Framework 2 the IWA Benchmark Simulation Models; Department of Industrial Electrical Engineering and Automation, Lund University: Lund, Sweden, 2006; pp. 1–35. [Google Scholar]
- Bonk, F.; Popp, D.; Weinrich, S.; Sträuber, H.; Kleinsteuber, S.; Harms, H.; Centler, F. Intermittent fasting for microbes: How discontinuous feeding increases functional stability in anaerobic digestion. Biotechnol. Biofuels 2018, 11, 274. [Google Scholar] [CrossRef] [Green Version]
- Donoso-Bravo, A.; Olivares, D.; Lesty, Y.; Bossche, H.V. Exploitation of the ADM1 in a XXI century wastewater resource recovery facility (WRRF): The case of codigestion and thermal hydrolysis. Water Res. 2020, 175, 115654. [Google Scholar] [CrossRef]
- Ellermeyer, S.; Hendrix, J.; Ghoochan, N. A theoretical and empirical investigation of delayed growth response in the continuous culture of bacteria. J. Theor. Biol. 2003, 222, 485–494. [Google Scholar] [CrossRef]
- Segura, C.; Vergara, C.; Seeger, M.; Donoso-Bravo, A.; Jeison, D. When constants are no longer constant: The case of inhibition in bioprocesses. Biochem. Eng. J. 2017, 123, 24–28. [Google Scholar] [CrossRef]
- Bertacchi, S.; Ruusunen, M.; Sorsa, A.; Sirviö, A.; Branduardi, P. Mathematical Analysis and Update of ADM1 Model for Biomethane Production by Anaerobic Digestion. Fermentation 2021, 7, 237. [Google Scholar] [CrossRef]
- Rodríguez, J.; Lema, J.M.; van Loosdrecht, M.C.M.; Kleerebezem, R. Variable stoichiometry with thermodynamic control in ADM1. Water Sci. Technol. 2006, 54, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Shi, E.; Li, J.; Zhang, M. Application of IWA Anaerobic Digestion Model No. 1 to simulate butyric acid, propionic acid, mixed acid, and ethanol type fermentative systems using a variable acidogenic stoichiometric approach. Water Res. 2019, 161, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Penumathsa, B.K.V.; Premier, G.C.; Kyazze, G.; Dinsdale, R.; Guwy, A.J.; Esteves, S.; Rodríguez, J. ADM1 can be applied to continuous bio-hydrogen production using a variable stoichiometry approach. Water Res. 2008, 42, 4379–4385. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Bravo, A.; Retamal, C.; Carballa, M.; Ruiz-Filippi, G.; Chamy, R. Influence of temperature on the hydrolysis, acidogenesis and methanogenesis in mesophilic anaerobic digestion: Parameter identification and modeling application. Water Sci. Technol. 2009, 60, 9–17. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inlet Conditions | |||
| VS | 30 kg m−3 | Ammonia nitrogen | 2 kg m−3 |
| COD/VS ratio | 1.5 | Inorganic Carbon | 0.004 kmole m−3 |
| CODs/CODt ratio | 0.1 | VFAs | 0 kg COD m−3 |
| Carbohydrate’s fraction 1 | 0.4 | Soluble gases | 0 kg COD m−3 |
| Protein’s fraction 1 | 0.4 | Anions | 0.04 kmole m−3 |
| Lipid’s fraction 1 | 0.2 | Cations | 0.18 kmole m−3 |
| Inert fraction | 0.2 | ||
| Model Extension: Feeding Strategy | |||
| Strategy | Notation | Momentaneous Flow | |
| Continuous | SC | 10 m3 d−1 | |
| 10 blocks of 1.2 h feeding | S1 | 20 m3 d−1 | |
| 5 blocks of 1.2 h feeding | S2 | 40 m3 d−1 | |
| 2 blocks of 1.2 h feeding | S3 | 100 m3 d−1 | |
| 1 block of 1.2 h feeding | S4 | 200 m3 d−1 | |
| Model Extension: Working Volume Reduction | |||
| Inlet flow | 10 m3 d−1 | Accumulation rate : 0 (base case), 0.01, 0.1, 0.25, 0.4, 0.6, 0.75 m3 d−1 | |
| Model Extension: Extra- Intra- Cellular Transport | |||
| Inlet flow | 10 m3 d−1 | Transport coefficient : 0.01, 0.05, 0.1, 1, 10, 100 d−1 | |
| Model Extension: Adaptation | |||
| Inlet flow | 10 m3 d−1 | Digester start-up = 1 d New (co) substrate addition = 100 d Adaptation constant : 0 (base case), 0.01, 0.1, 1, 10, 100, 1000 d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donoso-Bravo, A.; Sadino-Riquelme, M.C.; Valdebenito-Rolack, E.; Paulet, D.; Gómez, D.; Hansen, F. Comprehensive ADM1 Extensions to Tackle Some Operational and Metabolic Aspects in Anaerobic Digestion. Microorganisms 2022, 10, 948. https://doi.org/10.3390/microorganisms10050948
Donoso-Bravo A, Sadino-Riquelme MC, Valdebenito-Rolack E, Paulet D, Gómez D, Hansen F. Comprehensive ADM1 Extensions to Tackle Some Operational and Metabolic Aspects in Anaerobic Digestion. Microorganisms. 2022; 10(5):948. https://doi.org/10.3390/microorganisms10050948
Chicago/Turabian StyleDonoso-Bravo, Andrés, María Constanza Sadino-Riquelme, Emky Valdebenito-Rolack, David Paulet, Daniel Gómez, and Felipe Hansen. 2022. "Comprehensive ADM1 Extensions to Tackle Some Operational and Metabolic Aspects in Anaerobic Digestion" Microorganisms 10, no. 5: 948. https://doi.org/10.3390/microorganisms10050948