
Cyclic Electron Flow-Coupled Proton Pumping in Synechocystis sp. PCC6803 Is Dependent upon NADPH Oxidation by the Soluble Isoform of Ferredoxin:NADP-Oxidoreductase


Abstract
:1. Introduction
2. Materials and Methods
3. Results
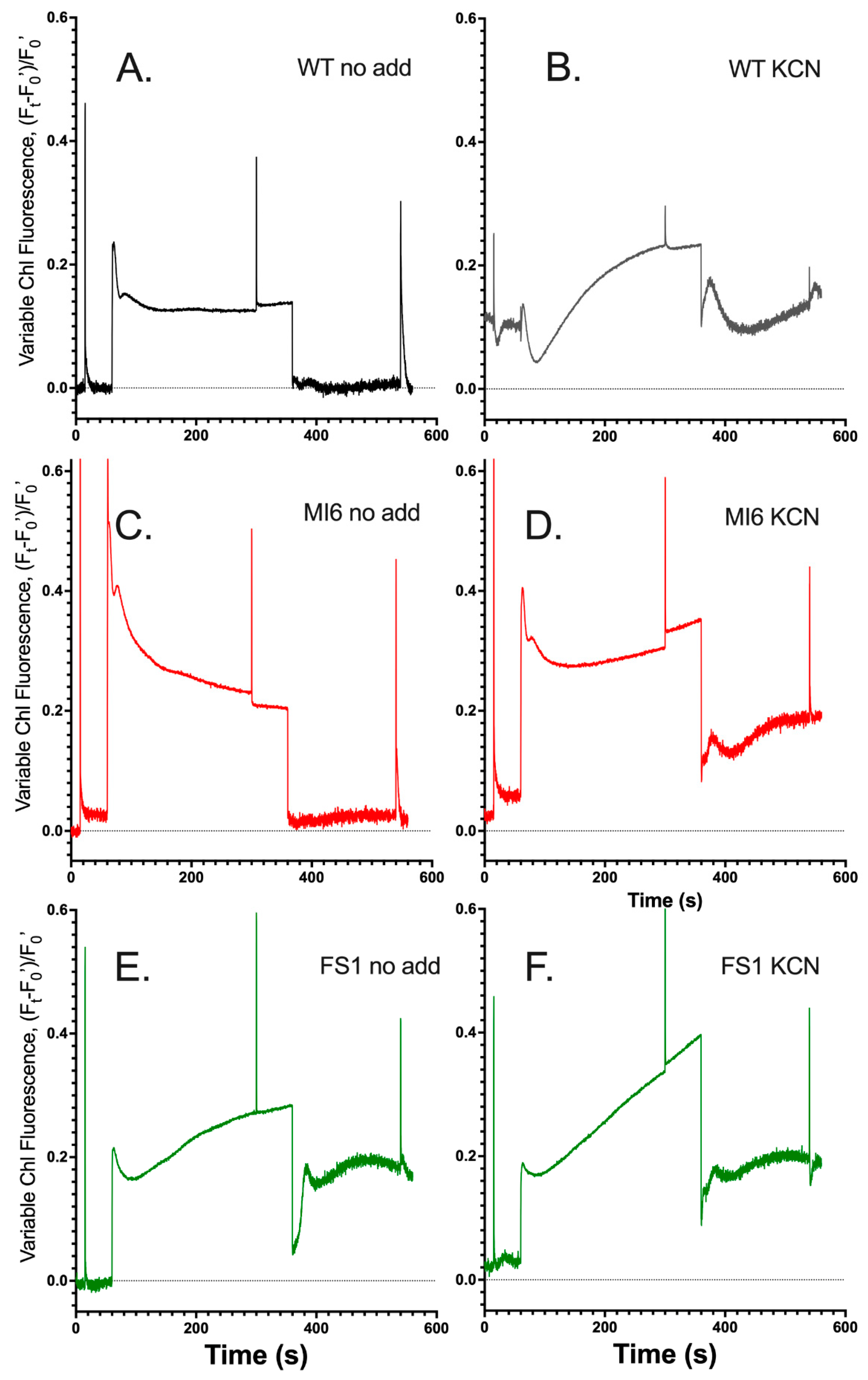
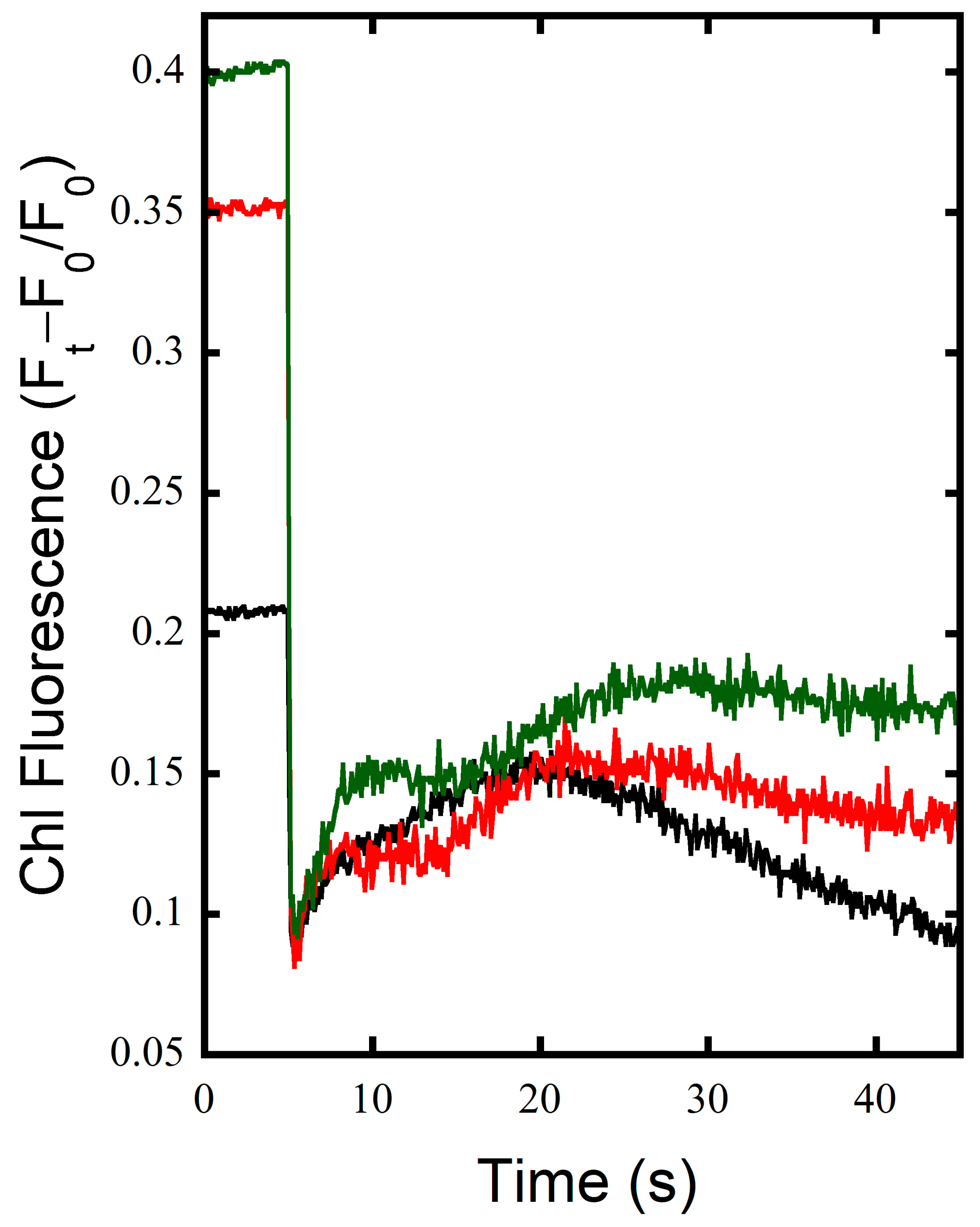
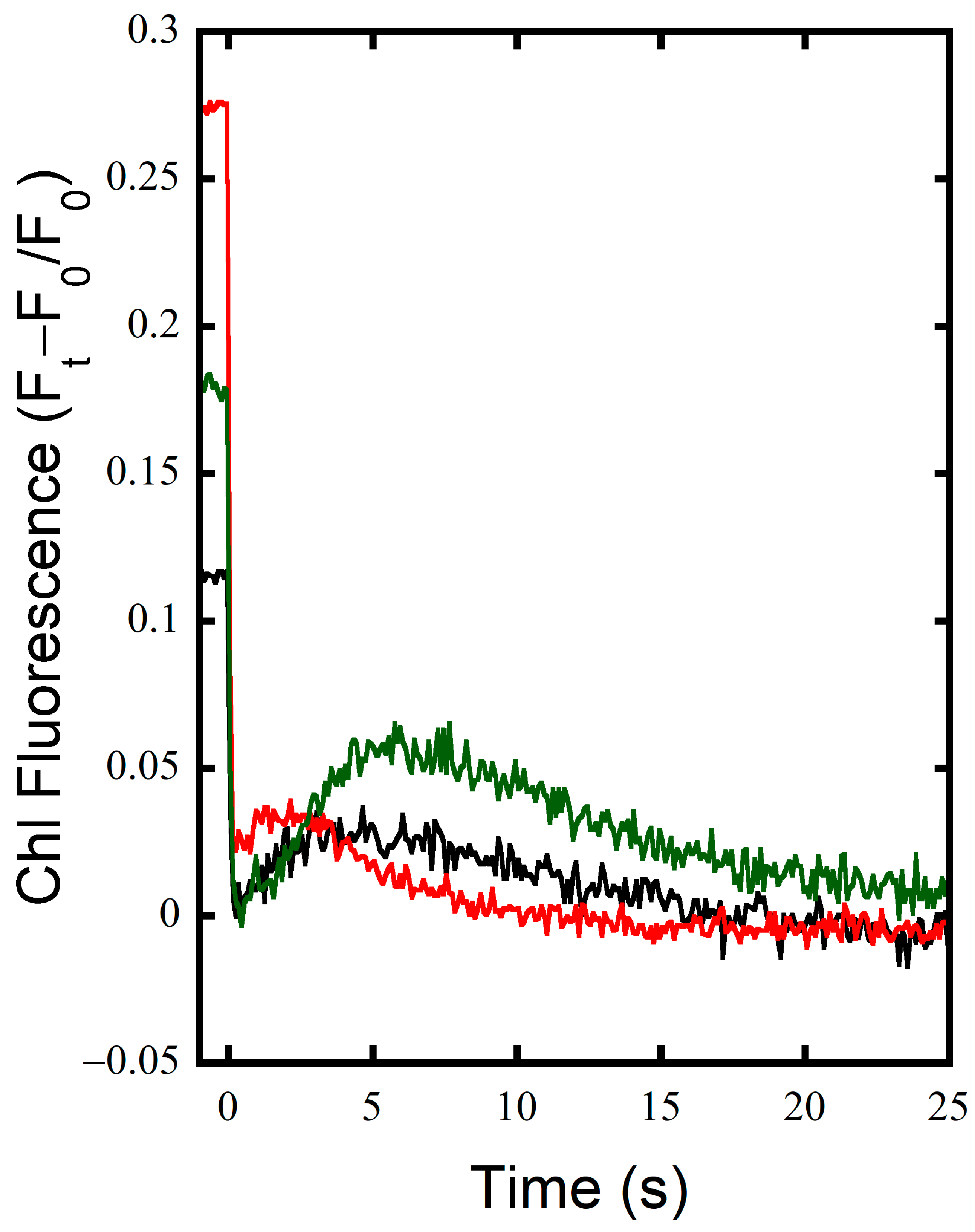
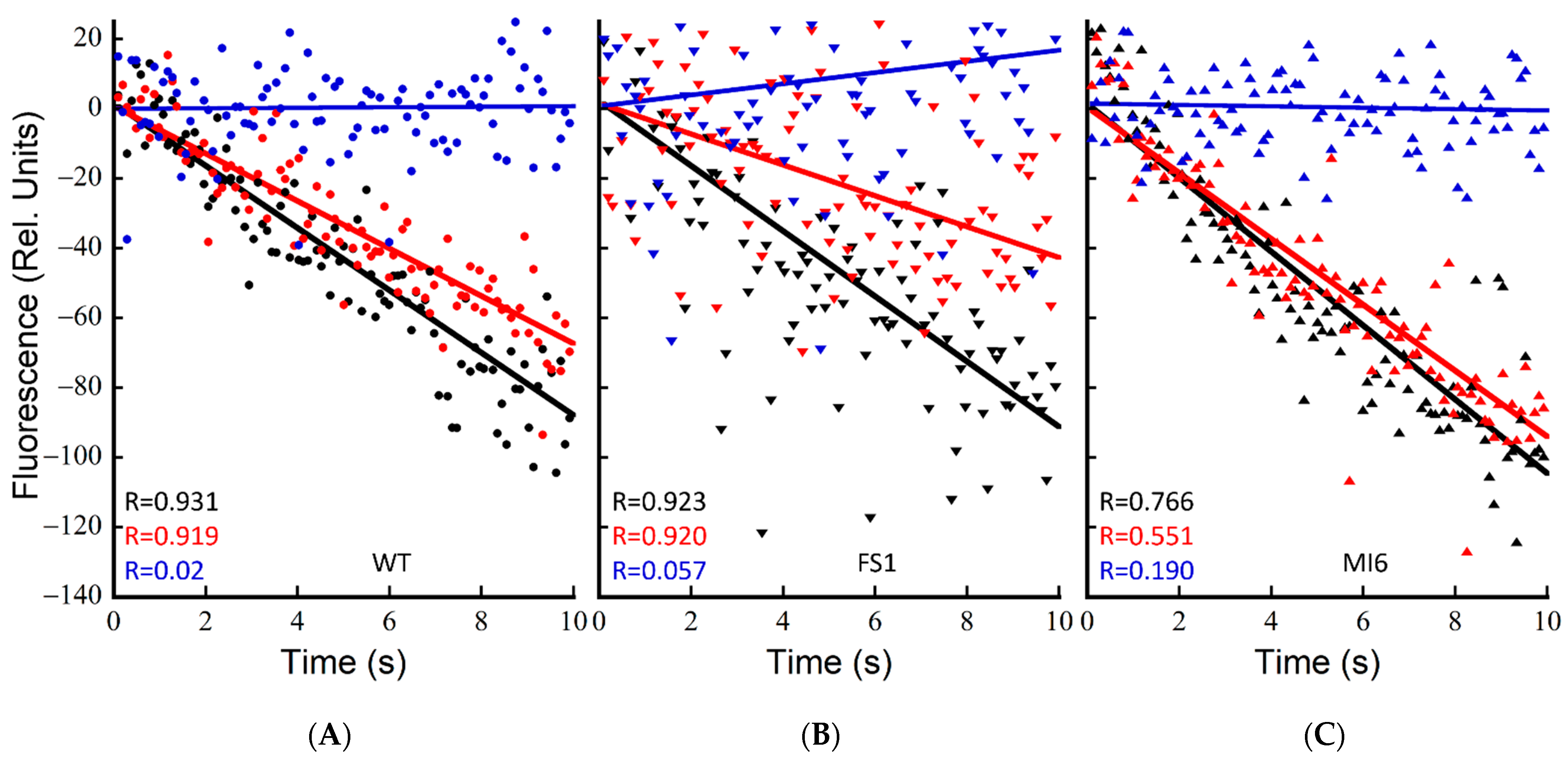
3.1. FNRs Has a Large Contribution to NDH-1 Cyclic Electron Flow
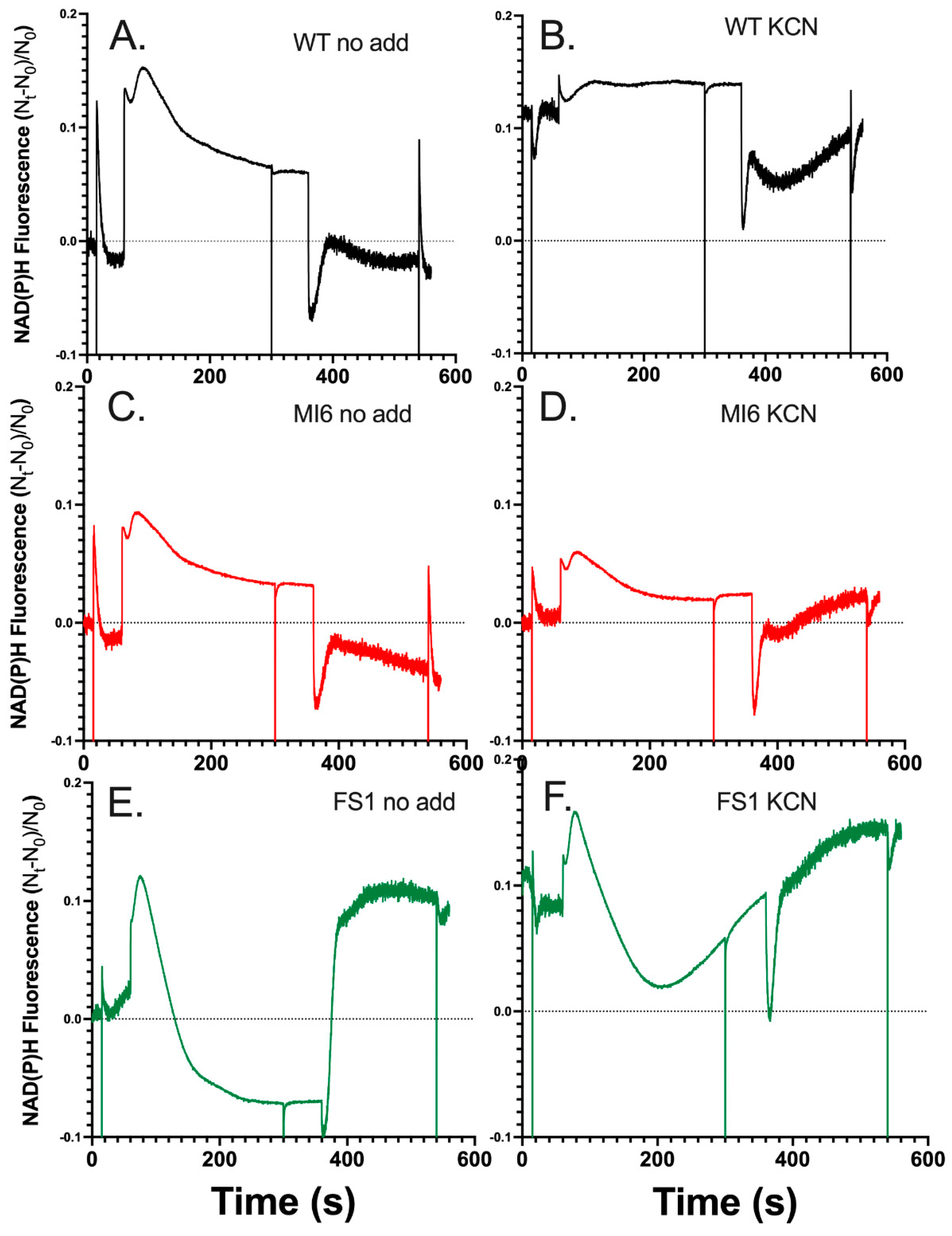
3.2. FNRS Enhances NADPH Oxidation during Illumination
3.3. The Presence of FNRs Enhances NDH-1 Powered Proton Pumping
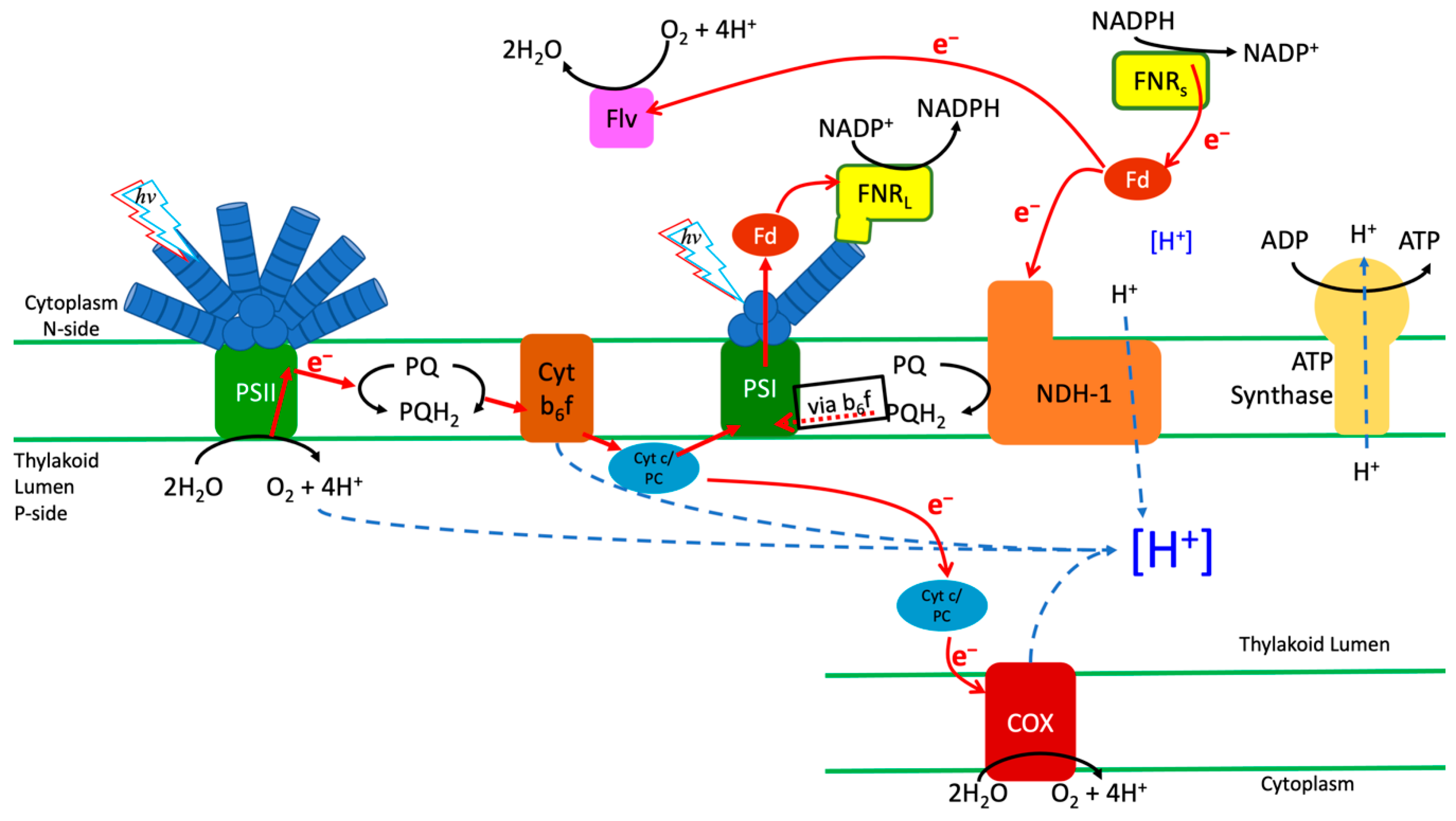
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Battchikova, N.; Wei, L.; Du, L.; Bersanini, L.; Aro, E.M.; Ma, W. Identification of novel Ssl0352 protein (NdhS), essential for efficient operation of cyclic electron transport around photosystem I, in NADPH:plastoquinone oxidoreductase (NDH-1) complexes of Synechocystis sp. PCC 6803. J. Biol. Chem. 2011, 286, 36992–37001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernat, G.; Appel, J.; Ogawa, T.; Rogner, M. Distinct roles of multiple NDH-1 complexes in the cyanobacterial electron transport network as revealed by kinetic analysis of P700+ reduction in various Ndh-deficient mutants of Synechocystis sp. strain PCC6803. J. Bacteriol. 2011, 193, 292–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikkanen, L.; Solymosi, D.; Jokel, M.; Allahverdiyeva, Y. Regulatory electron transport pathways of photosynthesis in cyanobacteria and microalgae: Recent advances and biotechnological prospects. Physiol. Plant 2021, 173, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Fisher, N.; Bricker, T.M.; Kramer, D.M. Regulation of photosynthetic cyclic electron flow pathways by adenylate status in higher plant chloroplasts. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 148081. [Google Scholar] [CrossRef] [PubMed]
- Dann, M.; Leister, D. Evidence that cyanobacterial Sll1217 functions analogously to PGRL1 in enhancing PGR5-dependent cyclic electron flow. Nat. Commun. 2019, 10, 5299. [Google Scholar] [CrossRef] [Green Version]
- Yeremenko, N.; Jeanjean, R.; Prommeenate, P.; Krasikov, V.; Nixon, P.J.; Vermaas, W.F.; Havaux, M.; Matthijs, H.C. Open reading frame ssr2016 is required for antimycin A-sensitive photosystem I-driven cyclic electron flow in the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2005, 46, 1433–1436. [Google Scholar] [CrossRef]
- Wang, C.; Yamamoto, H.; Shikanai, T. Role of cyclic electron transport around photosystem I in regulating proton motive force. Biochim. Biophys. Acta 2015, 1847, 931–938. [Google Scholar] [CrossRef] [Green Version]
- Miller, N.T.; Vaughn, M.D.; Burnap, R.L. Electron flow through NDH-1 complexes is the major driver of cyclic electron flow-dependent proton pumping in cyanobacteria. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148354. [Google Scholar] [CrossRef]
- Allahverdiyeva, Y.; Suorsa, M.; Tikkanen, M.; Aro, E.M. Photoprotection of photosystems in fluctuating light intensities. J. Exp. Bot. 2015, 66, 2427–2436. [Google Scholar] [CrossRef]
- Nikkanen, L.; Santana Sanchez, A.; Ermakova, M.; Rogner, M.; Cournac, L.; Allahverdiyeva, Y. Functional redundancy between flavodiiron proteins and NDH-1 in Synechocystis sp. PCC 6803. Plant J. 2020, 103, 1460–1476. [Google Scholar] [CrossRef]
- Mi, H.; Endo, T.; Schreiber, U.; Ogawa, T.; Asada, K. Electron donation from cyclic and respiratory flows to the photosynthetic intersystem chain is mediated by pyridine nucleotide dehydrogenase in the cyanobacterium Synechocystis PCC 6803. Plant Cell Physiol. 1992, 33, 1233–1237. [Google Scholar]
- Holland, S.C.; Kappell, A.D.; Burnap, R.L. Redox changes accompanying inorganic carbon limitation in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 2015, 1847, 355–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, H.; Endo, T.; Ogawa, T.; Asada, K. Thylakoid Membrane-Bound, NADPH-Specific Pyridine Nucleotide Dehydrogenase Complex Mediates Cyclic Electron Transport in the Cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 1995, 36, 661–668. [Google Scholar] [CrossRef]
- Zhang, C.; Shuai, J.; Ran, Z.; Zhao, J.; Wu, Z.; Liao, R.; Wu, J.; Ma, W.; Lei, M. Structural insights into NDH-1 mediated cyclic electron transfer. Nat. Commun. 2020, 11, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schluchter, W.M.; Bryant, D.A. Molecular characterization of ferredoxin NADP+ oxidoreductase in cyanobacteria: Cloning and sequence of the petH gene of Synechococcus sp. PCC 7002 and studies on the gene product. Biochemistry 1992, 31, 3092–3102, Erratum in Biochemistry 1992, 31, 5952. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.C.; Ughy, B.; Lagoutte, B.; Ajlani, G. A second isoform of the ferredoxin:NADP oxidoreductase generated by an in-frame initiation of translation. Proc. Natl. Acad. Sci. USA 2006, 103, 18368–18373. [Google Scholar] [CrossRef] [Green Version]
- Korn, A.; Ajlani, G.; Lagoutte, B.; Gall, A.; Setif, P. Ferredoxin:NADP+ oxidoreductase association with phycocyanin modulates its properties. J. Biol. Chem. 2009, 284, 31789–31797. [Google Scholar] [CrossRef] [Green Version]
- Korn, A. Respective Roles of the Ferredoxin: NADP-Oxidoreductase Isoforms in the Cyanobacterium Synechocystis sp. PCC 6803. Université Paris Sud-Paris XI. 2010. Available online: https://tel.archives-ouvertes.fr/tel-00460234 (accessed on 19 April 2022).
- Liu, H.; Weisz, D.A.; Zhang, M.M.; Cheng, M.; Zhang, B.; Zhang, H.; Gerstenecker, G.S.; Pakrasi, H.B.; Gross, M.L.; Blankenship, R.E. Phycobilisomes harbor FNR(L) in cyanobacteria. MBio 2019, 10, e00669-19. [Google Scholar] [CrossRef] [Green Version]
- Moal, G.; Lagoutte, B. Photo-induced electron transfer from photosystem I to NADP(+): Characterization and tentative simulation of the in vivo environment. Biochim. Biophys. Acta 2012, 1817, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Omairi-Nasser, A.; de Gracia, A.G.; Ajlani, G. A larger transcript is required for the synthesis of the smaller isoform of ferredoxin:NADP oxidoreductase. Mol. Microbiol. 2011, 81, 1178–1189. [Google Scholar] [CrossRef]
- Schuller, J.M.; Birrell, J.A.; Tanaka, H.; Konuma, T.; Wulfhorst, H.; Cox, N.; Schuller, S.K.; Thiemann, J.; Lubitz, W.; Setif, P.; et al. Structural adaptations of photosynthetic compolex I enable ferredoxin-dependent electron transfer. Science 2019, 363, 257–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, M.M. Simple conditions for growth of unicellular blue-green algae on plates. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.; Hurry, V.; Clarke, A.K.; Gustafsson, P.; Oquist, G. Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol. Mol. Biol. R 1998, 62, 667–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, U.; Klughammer, C. New NADPH/9-AA Module for the DUAL-PAM-100: Description, Operation and Examples of Application; Heinz Walz GmbH: Effeltrich, Germany, 2009; pp. 1–13. [Google Scholar]
- Palmgren, M.G. Acridine orange as a probe for measuring pH gradients across membranes: Mechanism and limitations. Anal. Biochem. 1991, 192, 316–321. [Google Scholar] [CrossRef]
- Nakamaru-Ogiso, E.; Kao, M.C.; Chen, H.; Sinha, S.C.; Yagi, T.; Ohnishi, T. The membrane subunit NuoL(ND5) is involved in the indirect proton pumping mechanism of Escherichia coli complex I. J. Biol. Chem. 2010, 285, 39070–39078. [Google Scholar] [CrossRef] [Green Version]
- Teuber, M.; Roegner, M.; Berry, S. Fluorescent probes for non-invasive bioenergetic studies of whole cyanobacterial cells. BBA 2001, 1506, 31–46. [Google Scholar] [CrossRef]
- Checchetto, V.; Segalla, A.; Allorent, G.; La Rocca, N.; Leanza, L.; Giacometti, G.M.; Uozumi, N.; Finazzi, G.; Bergantino, E.; Szabo, I. Thylakoid potassium channel is required for efficient photosynthesis in cyanobacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 11043–11048. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Endo, T.; Mi, H.; Asada, K. Quenching Analysis of Chlorophyll Fluorescence by the Saturation Pulse Method: Particular Aspects Relating to the Study of Eukaryotic Algae and Cyanobacteria. Plant Cell Physiol. 1995, 36, 873–882. [Google Scholar] [CrossRef]
- Renger, G.; Schreiber, U.; Amesz, J.; Fork, D. Practical applications of fluorometric methods to algae and higher plant research. In Light Emission by Plants and Bacteria; Govindjee, Amesz, J., Eds.; Academic Press: Orlando, FL, USA, 1986; pp. 589–616. [Google Scholar]
- Artier, J.; Holland, S.C.; Miller, N.; Zhang, M.; Burnap, R.L. Synthetic DNA system for structure function studies of the high affinity CO2 uptake (CupA) protein complex in cyanobacteria. BBA-Bioenerg. 2018, 1859, 1108–1118. [Google Scholar] [CrossRef]
- Shikanai, T.; Endo, T.; Hashimoto, T.; Yamada, Y.; Asada, K.; Yokota, A. Directed disruption of the tobacco ndhB gene impairs cyclic electron flow around photosystem I. Proc. Natl. Acad. Sci. USA 1998, 95, 9705–9709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Harada, T.; Ozaki, H.; Sonoike, K. Disruption of the ndhF1 gene affects Chl fluorescence through state transition in the cyanobacterium Synechocystis sp. PCC 6803, resulting in apparent high efficiency of photosynthesis. Plant Cell Physiol. 2013, 54, 1164–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernát, G.; Steinbach, G.; Kaňa, R.; Govindjee; Misra, A.N.; Prašil, O. On the origin of the slow M-T chlorophyll a fluorescence decline in cyanobacteria: Interplay of short-term light-responses. Photosynth. Res. 2018, 136, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Lazár, D.; Papageorgiou, G.C.; Govindjee. Chapter 5-Chlorophyll a Fluorescence in Cyanobacteria: Relation to Photosynthesis☆. In Cyanobacteria; Mishra, A.K., Tiwari, D.N., Rai, A.N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 79–130. [Google Scholar]
- Mullineaux, C.W.; Allen, J.F. State 1-State 2 transitions in the cyanobacterium Synechococcus 6301 are controlled by the redox state of electron carriers between photosystems I and II. Photosynth. Res. 1990, 23, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, A.F.; Choubeh, R.R.; Kirilovsky, D.; Wientjes, E.; van Amerongen, H. State transitions in cyanobacteria studied with picosecond fluorescence at room temperature. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148255. [Google Scholar] [CrossRef]
- Ranjbar Choubeh, R.; Wientjes, E.; Struik, P.C.; Kirilovsky, D.; van Amerongen, H. State transitions in the cyanobacterium Synechococcus elongatus 7942 involve reversible quenching of the photosystem II core. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 1059–1066. [Google Scholar] [CrossRef]
- Kana, R.; Kotabova, E.; Komarek, O.; Sediva, B.; Papageorgiou, G.C.; Govindjee; Prasil, O. The slow S to M fluorescence rise in cyanobacteria is due to a state 2 to state 1 transition. Biochim. Biophys. Acta 2012, 1817, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Ye, J.; Mi, H. Effects of low CO2 on NAD(P)H dehydrogenase, a mediator of cyclic electron transport around Photosystem I in the cyanobacterium Synechocystis PCC6803. Plant Cell Physiol. 2003, 44, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, T.G.; Savage, D.F.; Davies, K.M. Recent advances on the structure and function of NDH-1: The complex I of oxygenic photosynthesis. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2020, 11, 148254. [Google Scholar] [CrossRef]
- Tamoi, M.; Miyazaki, T.; Fukamizo, T.; Shigeoka, S. The Calvin cycle in cyanobacteria is regulated by CP12 via the NAD(H)/NADP(H) ratio under light/dark conditions. Plant J. 2005, 42, 504–513. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitors Added | Wild-Type | FS1 | MI6 | |
|---|---|---|---|---|
| Dark-adapted | Val + DCCD + KCN | 6.67 ± 0.65 s−1 †+ | 9.00 ± 1.65 s−1 ‡+₠ | 6.08 ± 1.27 s−1 ₡•₠ |
| Val + DCCD + KCN + DCMU | 3.44 ± 1.15 s−1 *†§ | 3.52 ± 0.48 s−1 **‡ℓ | 1.63 ± 1.00 s−1 ◊•§ℓ | |
| Val + DCCD + KCN + DCMU + CCCP | −0.26 ± 0.46 s−1 | −0.022 ± 0.28 s−1 | −0.88 ± 0.55 s−1 | |
| Light-adapted | Val + DCCD + KCN | 8.95 ± 2.57 s−1 | 10.60 ± 2.04 s−1 | 9.36 ± 2.59 s−1 ₡¤ |
| Val + DCCD + KCN + DCMU | 6.81 ± 0.50 s−1 *¢ø | 9.45 ± 0.72 s−1 **¢ж | 4.43 ± 2.22 s−1 ◊¤øж | |
| Val + DCCD + KCN + DCMU + CCCP | −0.08 ± 0.97 s−1 | 0.21 ± 0.27 s−1 | −1.61 ± 1.67 s−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, N.T.; Ajlani, G.; Burnap, R.L. Cyclic Electron Flow-Coupled Proton Pumping in Synechocystis sp. PCC6803 Is Dependent upon NADPH Oxidation by the Soluble Isoform of Ferredoxin:NADP-Oxidoreductase. Microorganisms 2022, 10, 855. https://doi.org/10.3390/microorganisms10050855
Miller NT, Ajlani G, Burnap RL. Cyclic Electron Flow-Coupled Proton Pumping in Synechocystis sp. PCC6803 Is Dependent upon NADPH Oxidation by the Soluble Isoform of Ferredoxin:NADP-Oxidoreductase. Microorganisms. 2022; 10(5):855. https://doi.org/10.3390/microorganisms10050855
Chicago/Turabian StyleMiller, Neil T., Ghada Ajlani, and Robert L. Burnap. 2022. "Cyclic Electron Flow-Coupled Proton Pumping in Synechocystis sp. PCC6803 Is Dependent upon NADPH Oxidation by the Soluble Isoform of Ferredoxin:NADP-Oxidoreductase" Microorganisms 10, no. 5: 855. https://doi.org/10.3390/microorganisms10050855