
Optimization of Bacillus amyloliquefaciens BLB369 Culture Medium by Response Surface Methodology for Low Cost Production of Antifungal Activity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Cultivation
2.2. Determination of the Antifungal Activity
2.3. Identification of Significant Factors Using Plackett–Burman Design
2.4. Optimization by Steepest Ascent Method
2.5. Central Composite Design and Response Surface
CCD Matrix and Antifungal Activity
2.6. Regression Models and Statistical Analysis
3. Results and Discussion
3.1. Screening of the Significant Medium Components Using PBD
3.2. Optimization by Steepest Ascent Method

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Candy Waste | Peptone | NaCl | Antifungal Activity (AU/mL) | |||
|---|---|---|---|---|---|---|---|
0.354 # | X1 (g/L) 1.77 # | 1 # | X2 (g/L) 5 # | 0.289 # | X3 (g/L) 1.58 # | ||
| 1 | 0 | 15.00 | 0 | 5 | 0 | 2.00 | 100 |
| 2 | 0.354 | 16.77 | 1 | 10 | 0.289 | 2.58 | 100 |
| 3 | 0.710 | 18.55 | 2 | 15 | 0.578 | 3.16 | 175 |
| 4 | 1.065 | 20.25 | 3 | 20 | 0.867 | 3.74 | 250 |
| 5 | 1.420 | 22.00 | 4 | 25 | 1.156 | 4.30 | 250 |
| 6 | 1.775 | 23.75 | 5 | 30 | 1.445 | 4.90 | 225 |
3.3. Optimization of the Selected Medium Components Using the CCD
| Run | Candy Waste | Peptone | NaCl | Antifungal Activity | |||
|---|---|---|---|---|---|---|---|
| X1 (g/L) | X2 (g/L) | X3 (g/L) | Y1 (AU/mL) | ||||
| 1 | +1 | 22 | −1 | 15 | +1 | 4.3 | 175 |
| 2 | −1 | 18.4 | +1 | 25 | +1 | 4.3 | 150 |
| 3 | −1 | 18.4 | −1 | 15 | −1 | 3.1 | 175 |
| 4 | 0 | 20.2 | 0 | 20 | 0 | 3.7 | 250 |
| 5 | 0 | 20.2 | 0 | 20 | 0 | 3.7 | 250 |
| 6 | +1 | 22 | +1 | 25 | −1 | 3.1 | 150 |
| 7 | −1 | 18.4 | −1 | 15 | +1 | 4.3 | 175 |
| 8 | +1 | 22 | +1 | 25 | +1 | 4.3 | 150 |
| 9 | −1 | 18.4 | +1 | 25 | −1 | 3.1 | 175 |
| 10 | 0 | 20.2 | 0 | 20 | 0 | 3.7 | 250 |
| 11 | +1 | 22 | −1 | 15 | −1 | 3.1 | 125 |
| 12 | 0 | 20.2 | 0 | 20 | 0 | 3.7 | 250 |
| 13 | 0 | 20.2 | 0 | 20 | −2 | 2.5 | 125 |
| 14 | 0 | 20.2 | 0 | 20 | 0 | 3.7 | 250 |
| 15 | +2 | 23.8 | 0 | 20 | 0 | 3.7 | 250 |
| 16 | 0 | 20.2 | +2 | 30 | 0 | 3.7 | 125 |
| 17 | −2 | 16.6 | 0 | 20 | 0 | 3.7 | 200 |
| 18 | 0 | 20.2 | 0 | 20 | 0 | 3.7 | 250 |
| 19 | 0 | 20.2 | 0 | 20 | +2 | 4.9 | 125 |
| 20 | 0 | 20.2 | −2 | 10 | 0 | 3.7 | 125 |
3.3.1. Regression Models for Antifungal Activity and Their Comparison
Polynomial Regression
Trigonometric Model
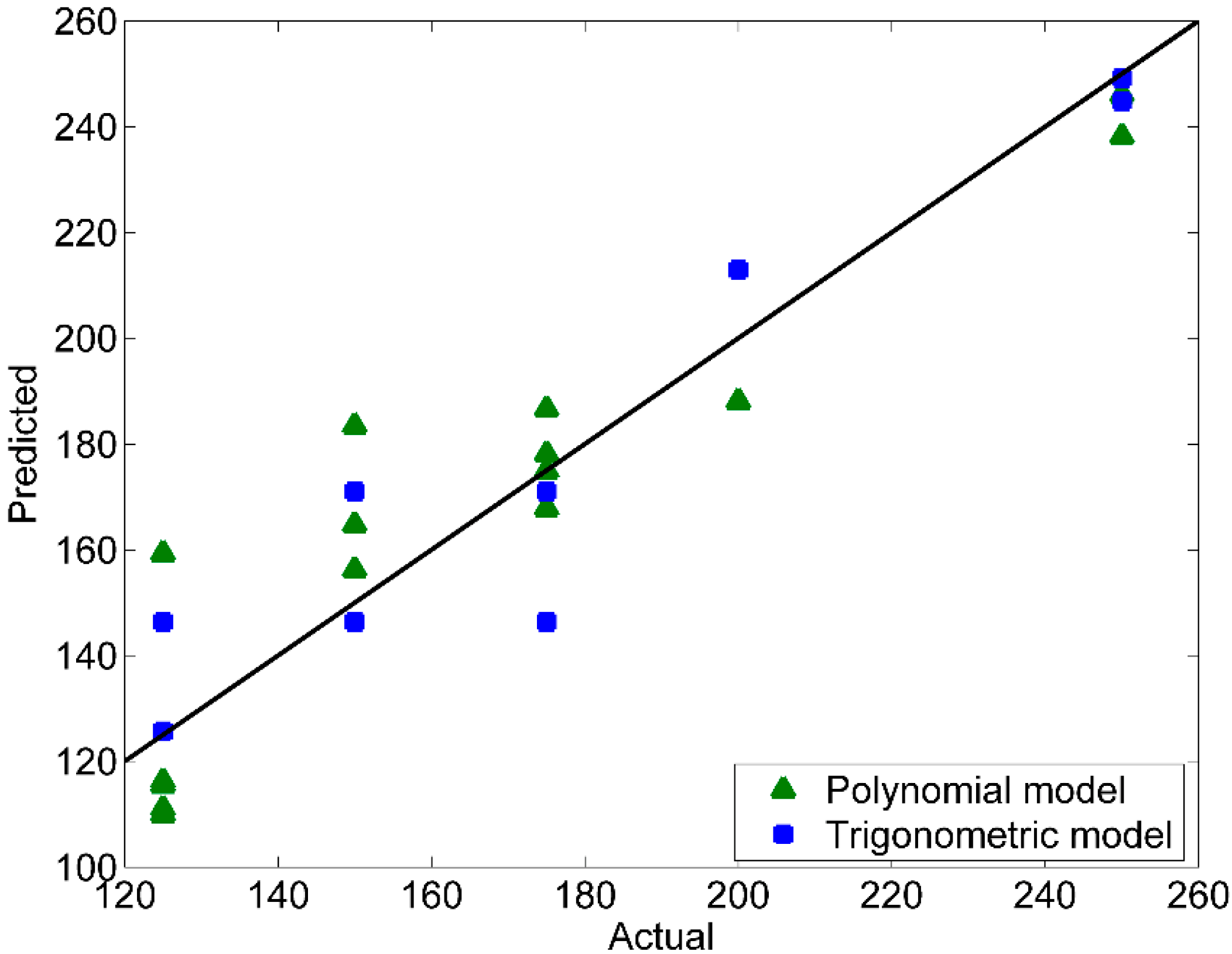
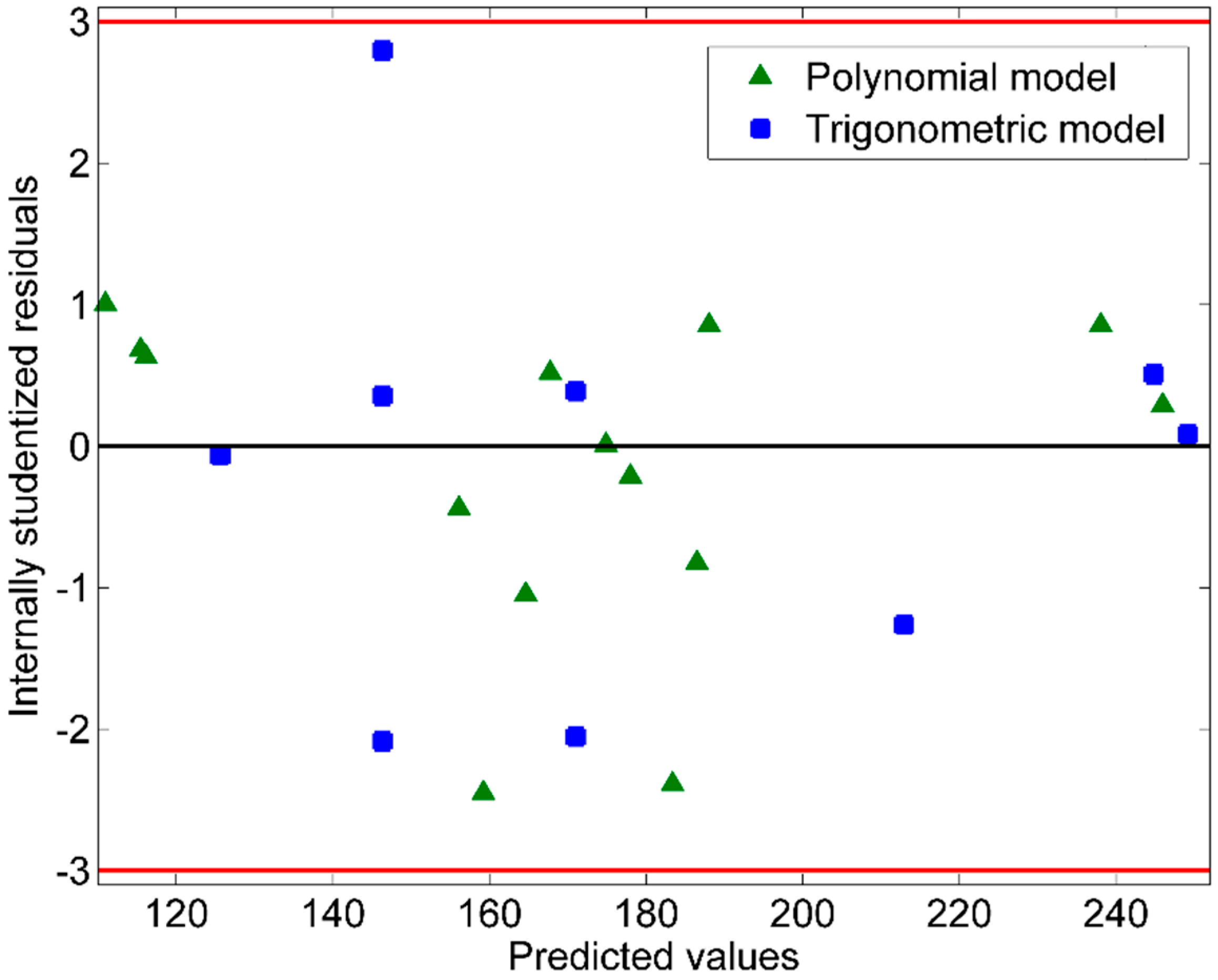
Predicted Versus Actual Plot and Residuals Versus Fits Plot
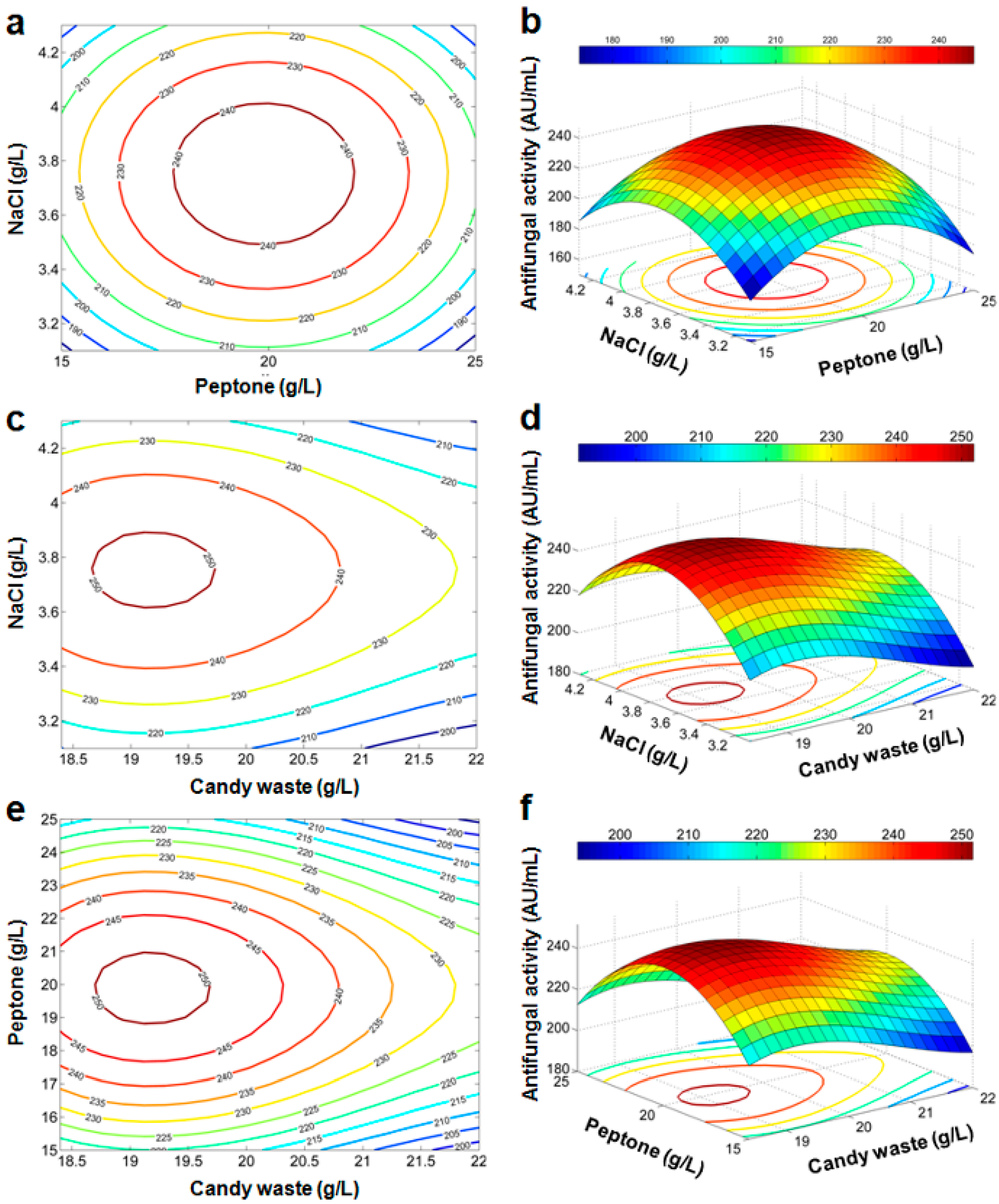
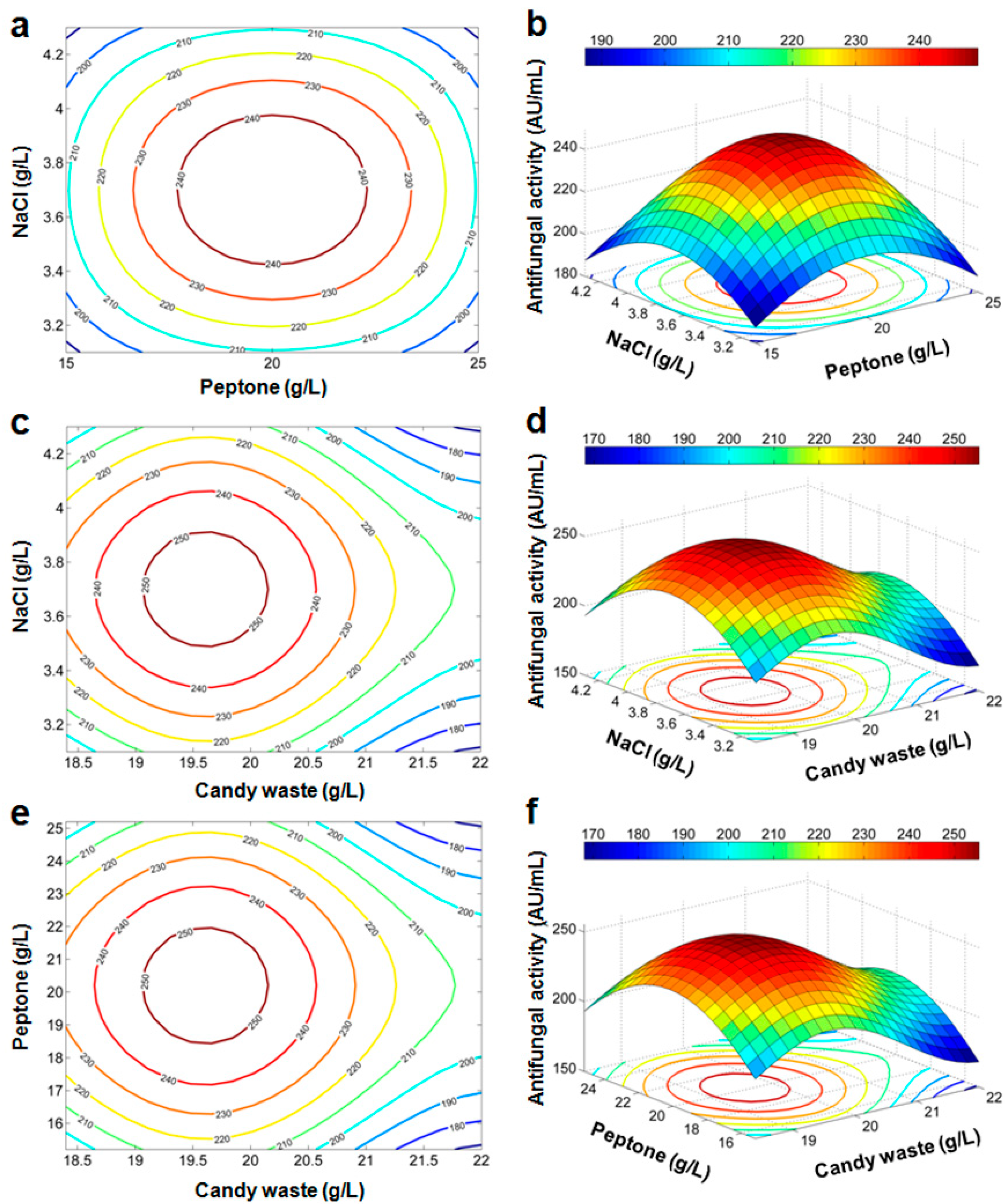
Response Surface and Contour Plots
Optimization of Production Conditions of the Antifungal Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal-Fusarium graminearum interactions: What we have learned so far. Mol. Plant Pathol. 2018, 19, 764–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauwane, M.E.; Ogugua, U.V.; Kalu, C.M.; Ledwaba, L.K.; Woldesemayat, A.A.; Ntushelo, K. Pathogenicity and Virulence Factors of Fusarium graminearum Including Factors Discovered Using Next Generation Sequencing Technologies and Proteomics. Microorganisms 2020, 8, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirgozliev, S.R.; Edwards, S.G.; Hare, M.C.; Jenkinson, P. Strategies for the control of Fusarium head blight in cereals. Eur. J. Plant Pathol. 2003, 109, 731–742. [Google Scholar] [CrossRef]
- Paul, P.A.; Lipps, P.E.; Hershman, D.E.; McMullen, M.P.; Draper, M.A.; Madden, L.V. Efficacy of triazole-based fungicides for fusarium head blight and deoxynivalenol control in wheat: A multivariate meta-analysis. Phytopathology 2008, 98, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.G.; Pirgozliev, S.R.; Hare, M.C.; Jenkinson, P. Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against fusarium head blight of winter wheat. Appl. Environ. Microbiol. 2001, 67, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- Paul, P.A.; McMullen, M.P.; Hershman, D.E.; Madden, L.V. Meta-analysis of the effects of triazole-based fungicides on wheat yield and test weight as influenced by fusarium head blight intensity. Phytopathology 2010, 100, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Machado, F.J.; Santana, F.M.; Lau, D.; Del Ponte, E.M. Quantitative review of the effects of triazole and benzimidazole fungicides on fusarium head blight and wheat yield in Brazil. Plant Dis. 2017, 101, 1633–1641. [Google Scholar] [CrossRef] [Green Version]
- Kotowicz, N.K.; Frąc, M.; Lipiec, J. The importance of Fusarium fungi in wheat cultivation—Pathogenicity and mycotoxins production: A review. J. Anim. Plant Sci. 2014, 21, 3326–3343. [Google Scholar]
- Mateo, E.M.; Valle-Algarra, F.M.; Mateo, R.; Jiménez, M.; Magan, N. Effect of fenpropimorph, prochloraz and tebuconazole on growth and production of T-2 and HT-2 toxins by Fusarium langsethiae in oat-based medium. Int. J. Food Microbiol. 2011, 151, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Kafle., S.; Vaidya, A.; Pradhan., B.; Jørs, E.; Onta, S. Factors Associated with Practice of Chemical Pesticide Use and Acute Poisoning Experienced by Farmers in Chitwan District Nepal. Int. J. Environ. Res. Public Health 2021, 18, 4194. [Google Scholar] [CrossRef]
- Roman, D.L.; Voiculescu, D.I.; Filip, M.; Ostafe, V.; Isvoran., A. Effects of Triazole Fungicides on Soil Microbiota and on the Activities of Enzymes Found in Soil: A Review. Agriculture 2021, 11, 893. [Google Scholar] [CrossRef]
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An Overview of Some Biopesticides and Their Importance in Plant Protection for Commercial Acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- Asari, S.; Ongena, M.; Debois, D.; Pauw, E.; Chen, K.; Bejai, S.; Meijer, J. Insights into the molecular basis of biocontrol of Brassica pathogens by Bacillus amyloliquefaciens UCMB5113 lipopeptides. Ann. Bot. 2017, 20, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Kunova, A.; Bonaldi, M.; Saracchi, M.; Pizzatti, C.; Chen, X.; Cortesi, P. Selection of Streptomyces against soil borne fungal pathogens by a standardized dual culture assay and evaluation of their effects on seed germination and plant growth. BMC Microbiol. 2016, 16, 272. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Yang, Y.; Yuan, Q.; Shi, G.; Wu, L.; Lou, Z.; Huo, R.; Wu, H.; Borriss, R.; Gao, X. Bacillomycin D Produced by Bacillus amyloliquefaciens Is Involved in the Antagonistic Interaction with the Plant-Pathogenic Fungus Fusarium graminearum. Appl. Environ. Microbiol. 2017, 83, e01075-17. [Google Scholar] [CrossRef] [Green Version]
- Penha, R.O.; Vandenberghe, L.P.S.; Faulds, C.; Soccol, V.; Soccol, C. Bacillus lipopeptides as powerful pest control agents for a more sustainable and healthy agriculture: Recent studies and innovations. Planta 2020, 251, 70. [Google Scholar] [CrossRef] [Green Version]
- Mora, I.; Cabrefiga, J.; Montesinos, E. Cyclic lipopeptide biosynthetic genes and products, and inhibitory activity of plant-associated Bacillus against phytopathogenic bacteria. PLoS ONE 2015, 10, e0127738. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, R.A.B.; Stedel, C.; Garagounis, C.; Nefzi, A.; Jabnoun-Khiareddine, H.; Papadopoulou, K.K.; Daami-Remadi, M. Involvement of lipopeptide antibiotics and chitinase genes and induction of host defense in suppression of Fusarium wilt by endophytic Bacillus spp. in tomato. Crop Prot. 2017, 99, 45–58. [Google Scholar] [CrossRef]
- Han, J.H.; Shim, H.; Shin, J.H.; Kim, K.S. Antagonistic activities of Bacillus spp. strains isolated from tidal flat sediment towards anthracnose pathogens Colletotrichum acutatum and C. gloeosporioides in South Korea. Plant Pathol. J. 2015, 31, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Hamid, R.; Khan, M.A.; Ahmad, M.; Ahmad, M.M.; Abdin, M.Z.; Musarrat, J.; Javed, S. Chitinases: An update. J. Pharm. Bioallied Sci. 2013, 5, 21–29. [Google Scholar]
- Zalila-Kolsi, I.; Sallemi, S.; Tounsi, S.; Jamoussi, K. Heterologous expression and periplasmic secretion of an antifungal Bacillus amyloliquefaciens BLB369 endo-β-1,3-1,4-glucanase in Escherichia coli. J. Phytopathol. 2017, 166, 28–33. [Google Scholar] [CrossRef]
- Mulatu, A.; Alemu, T.; Megersa, N.; Vetukuri, R.R. Optimization of Culture Conditions and Production of Bio-Fungicides from Trichoderma Species under Solid-State Fermentation Using Mathematical Modeling. Microorganisms 2021, 9, 1675. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Zhu, P.; Liang, J.; Xu, Z.; Feng, X.; Liu, Y.; Xu, H.; Li, S. Economical production of isomaltulose from agricultural residues in a system with sucrose isomerase displayed on Bacillus subtilis spores. Bioprocess Biosyst. Eng. 2020, 43, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.M.; Bassi, A. Investigation of biomass concentration, lipid production, and cellulose content in Chlorella vulgaris cultures using response surface methodology. Biotechnol. Bioeng. 2013, 110, 2114–2122. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Mehta, P.K.; Bhatia, R.K.; Bhalla, T.C. Optimization of arylacetonitrilase production from Alcaligenes sp. MTCC 10675 and its application in mandelic acid synthesis. Appl. Microbiol. Biotechnol. 2014, 98, 83–94. [Google Scholar] [CrossRef]
- Hsieh, S.C.; Liu, J.M.; Pua, X.H.; Ting, Y.; Hsu, R.J.; Cheng, K.C. Optimization of Lactobacillus acidophilus cultivation using taro waste and evaluation of its biological activity. Appl. Microbiol. Biotechnol. 2016, 100, 2629–2639. [Google Scholar] [CrossRef]
- Samaram, S.; Mirhosseini, H.; Tan, C.P.; Ghazali, H.M.; Bordbar, S.; Serjouie, A. Optimisation of ultrasound-assisted extraction of oil from papaya seed by response surface methodology: Oil recovery, radical scavenging antioxidant activity, and oxidation stability. Food Chem. 2015, 172, 7–17. [Google Scholar] [CrossRef]
- Samavati, V. Central composite rotatable design for investigation of microwave-assisted extraction of okra pod hydrocolloid. Int. J. Biol. Macromol. 2013, 61, 142–149. [Google Scholar] [CrossRef]
- Caldeira, A.T.; Arteiro, J.M.; Roseiro, J.C.; Neves, J.; Vicente, H. An artificial intelligence approach to Bacillus amyloliquefaciens CCMI 1051 cultures: Application to the production of anti-fungal compounds. Bioresour. Technol. 2011, 102, 1496–1502. [Google Scholar] [CrossRef]
- Tang, X.J.; He, G.Q.; Chen, Q.H.; Zhang, X.Y.; Ali, M.A. Medium optimization for the production of thermal stable β-glucanase by Bacillus subtilis ZJF-1A5 using response surface methodology. Bioresour. Technol. 2004, 93, 175–181. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, M.; Li, Y.; Cui, Y.; Zhang, Y.; Wang, Z.; Wang, M.; Huang, Y. Application of response surface methodology to optimize the production of antimicrobial metabolites by Micromonospora Y15. Biotechnol. Biotechnol. Equip. 2017, 31, 1016–1025. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Yu, K.O.; Ramzi, A.B.; Choe, S.H.; Kim, S.W.; Han, S.O. Improvement of surfactin production in Bacillus subtilis using synthetic wastewater by overexpression of specific extracellular signaling peptides, comX and phrC. Biotechnol. Bioeng. 2012, 109, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Hongsheng, W.; Qirong. Use of response surface methodology to evaluate the effect of metal ions (Ca2+, Ni2+, Mn2+, Cu2+) on production of antifungal compounds by Paenibacillus polymyxa. Bioresour. Technol. 2010, 101, 1904–1912. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Han, Y.; Tan, X.Q.; Wang, J.; Zhou, Z.J. Optimization of antifungal lipopeptide production from Bacillus sp. BH072 by response surface methodology. J. Microbiol. 2014, 52, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, Z.; Xiu, Z. High production of optically pure (3R)-acetoin by a newly isolated marine strain of Bacillus subtilis CGMCC 13141. Bioprocess Biosyst. Eng. 2019, 42, 475–483. [Google Scholar] [CrossRef]
- De Oliveira, C.C.; de Souza, A.K.S.; de Castro, R.J.S. Bioconversion of Chicken Feather Meal by Aspergillus niger: Simultaneous Enzymes Production Using a Cost-Effective Feedstock Under Solid State Fermentation. Indian J. Microbiol. 2019, 59, 209–216. [Google Scholar] [CrossRef]
- Sreedharan, S.M.; Singh, S.P.; Singh, R. Flower Shaped Gold Nanoparticles: Biogenic Synthesis Strategies and Characterization. Indian J. Microbiol. 2019, 59, 321–327. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Ben Mahmoud, A.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef]
- Ahimou, F.; Jacques, P.; Daleu, M. Surfactin and iturin A effects on Bacillus subtilis surface hydrophobicity. Enzyme Microb. Technol. 2000, 27, 749–754. [Google Scholar] [CrossRef]
- Plackett, R.L.; Burman, J.P. The design of optimum multifactorial experiments. Biometrika 1946, 33, 305–325. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Hocking, R.R. The analysis and selection of variables in linear regression. Biometrics 1976, 32, 1–49. [Google Scholar] [CrossRef]
- Theil, H. Economic Forecasts and Policy; North-Holland: Amsterdam, The Netherlands, 1961. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Hurvich, C.M.; Tsai, C.L. Model selection for extended quasi-likelihood models in small samples. Biometrics 1995, 5, 1077–1084. [Google Scholar] [CrossRef]




| Run | A: Candy Waste | B: Fish Extract | C: Peptone | D: Yeast Extract | E: NaCl | F: MgSO4 | G: MnSO4 | Antifungal Activity (AU/mL) |
|---|---|---|---|---|---|---|---|---|
| 1 | +1 | −1 | −1 | +1 | −1 | +1 | +1 | 75 |
| 2 | +1 | +1 | −1 | −1 | +1 | −1 | +1 | 75 |
| 3 | +1 | +1 | +1 | −1 | −1 | +1 | −1 | 150 |
| 4 | −1 | +1 | +1 | +1 | −1 | −1 | +1 | 125 |
| 5 | +1 | −1 | +1 | +1 | +1 | −1 | −1 | 175 |
| 6 | −1 | +1 | −1 | +1 | +1 | +1 | −1 | 62.5 |
| 7 | −1 | −1 | +1 | −1 | +1 | +1 | +1 | 150 |
| 8 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 0 |
| Coefficient | Value | p-Value | Significance |
|---|---|---|---|
| α0 | 101.563 | 3.49 × 10−12 | *** |
| α1 | 17.188 | 4.15 × 10−6 | *** |
| α2 | 1.563 | 0.347 | |
| α3 | 48.438 | 1.27 × 10−9 | *** |
| α4 | 7.813 | 1.05 × 10−3 | ** |
| α5 | 14.063 | 1.85 × 10−5 | *** |
| α6 | 7.813 | 1.05 × 10−3 | ** |
| α7 | 4.688 | 0.017 | * |
| Model | Term | Coefficient | p-Value | Significance |
|---|---|---|---|---|
| Polynomial model # | Intercept | −2050.5 | 0.03693 | * |
| X1 | 64.534 | 0.2974 | ||
| X2 | 57.418 | 0.0151 | * | |
| X3 | 573 | 0.0062 | ** | |
| X12 | −2.5428 | 0.0703 | ||
| X22 | −1.3295 | 9.75 × 10−6 | *** | |
| X32 | −92.33 | 9.75 × 10−6 | *** | |
| X1 X2 | 0.3472 | 0.6739 | ||
| X1 X3 | 8.6806 | 0.2227 | ||
| X2 X3 | −3.125 | 0.2227 | ||
| Retained polynomial model after applying the stepwise technique § | Intercept | −12312 | 0.0108 | * |
| X1 | 1623.8 | 0.0211 | * | |
| X2 | 52.849 | 7.74 × 10−7 | *** | |
| X3 | 351.1 | 7.21 × 10−7 | *** | |
| X12 | −78.306 | 0.0245 | * | |
| X22 | −1.329 | 6.38 × 10−7 | *** | |
| X13 | 1.2503 | 0.0285 | * | |
| X33 | −8.3034 | 6.34 × 10−7 | *** | |
| Trigonometric model £ | Intercept | 144.52 | 3.1 × 10−19 | *** |
| 87.202 | 6.6 × 10−13 | *** | ||
| 23.775 | 2.9 × 10−6 | *** |
| Model | Mean of Square | F-Value | p-Value | |
|---|---|---|---|---|
| Retained polynomial model | Total | 2728.6 | ||
| Model | 6876.2 | 22.24 | 5.717 × 10−6 | |
| Residual | 309.17 | |||
| Lack of fit | 530.01 | Inf | 0 | |
| Pure error | 0 | |||
| Trigonometric model | Total | 2728.6 | ||
| Model | 24926 | 212.71 | 9.32 × 10−13 | |
| Residual | 117.18 | |||
| Lack of fit | 175.77 | 2.0624 | 0.1413 | |
| Pure error | 85.227 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalila-Kolsi, I.; Kessentini, S.; Tounsi, S.; Jamoussi, K. Optimization of Bacillus amyloliquefaciens BLB369 Culture Medium by Response Surface Methodology for Low Cost Production of Antifungal Activity. Microorganisms 2022, 10, 830. https://doi.org/10.3390/microorganisms10040830
Zalila-Kolsi I, Kessentini S, Tounsi S, Jamoussi K. Optimization of Bacillus amyloliquefaciens BLB369 Culture Medium by Response Surface Methodology for Low Cost Production of Antifungal Activity. Microorganisms. 2022; 10(4):830. https://doi.org/10.3390/microorganisms10040830
Chicago/Turabian StyleZalila-Kolsi, Imen, Sameh Kessentini, Slim Tounsi, and Kaïs Jamoussi. 2022. "Optimization of Bacillus amyloliquefaciens BLB369 Culture Medium by Response Surface Methodology for Low Cost Production of Antifungal Activity" Microorganisms 10, no. 4: 830. https://doi.org/10.3390/microorganisms10040830