
Responses of Soil Microbial Communities and Networks to Precipitation Change in a Typical Steppe Ecosystem of the Loess Plateau


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
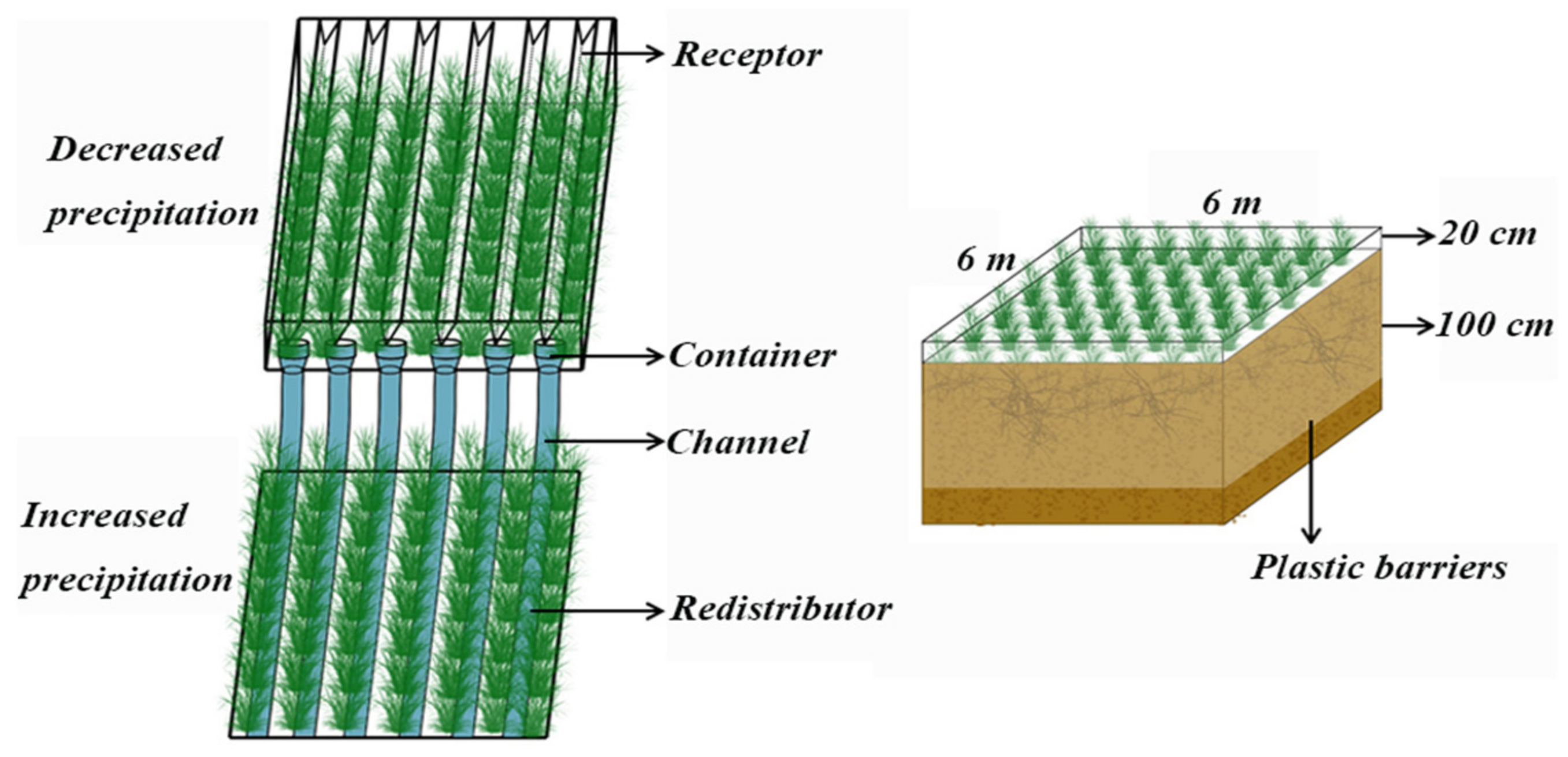
2.2. Experimental Design and Sample Collection and Processing
2.3. Plant and Soil Physicochemical Characteristic Analyses
2.4. Soil DNA Extraction and Gene Sequencing
2.5. Statistical Analyses
3. Results
3.1. Soil and Vegetation Characteristic under Precipitation Treatments
3.2. Diversity of Soil Microbial Communities under Precipitation Treatments
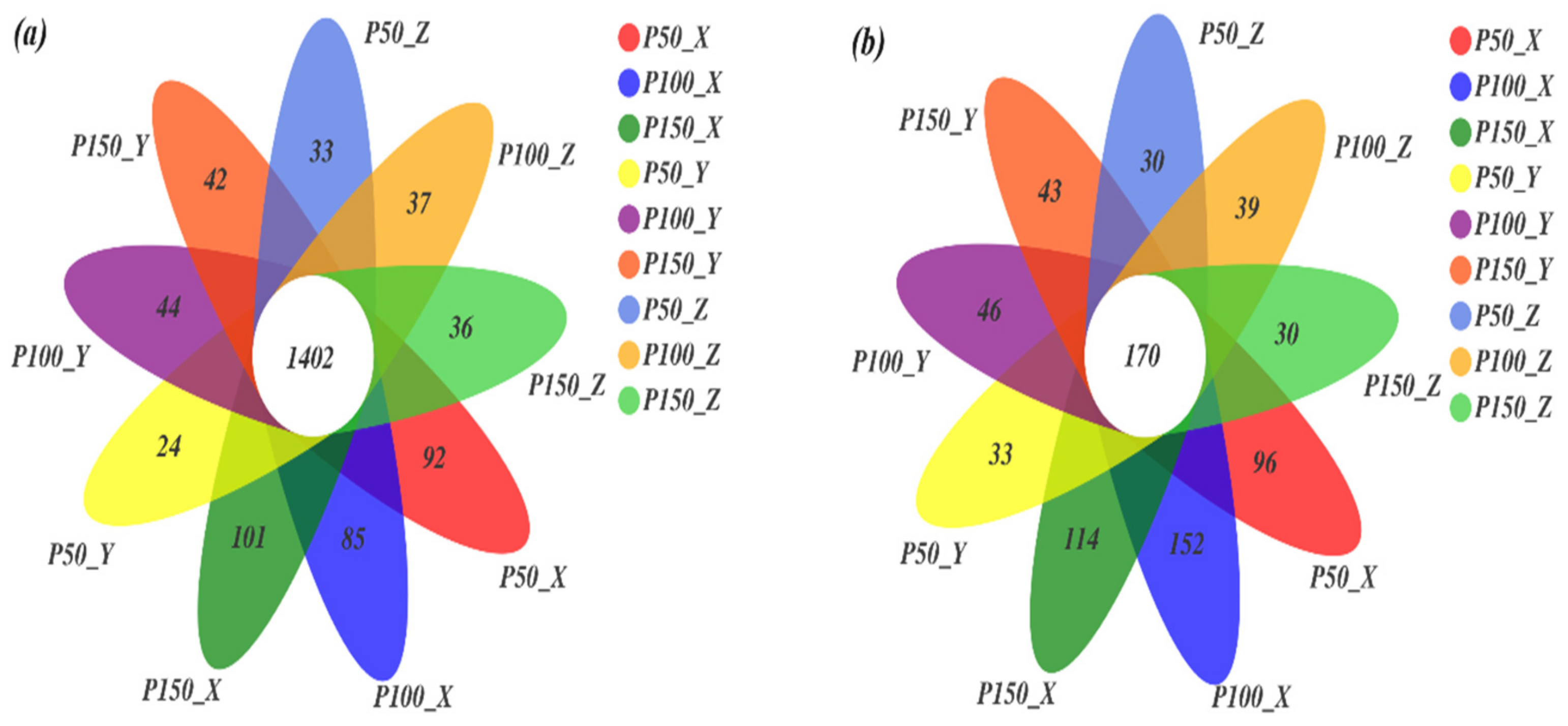
3.2.1. Venn Diagram of Microorganisms under Precipitation Treatments
3.2.2. Diversity of Microbes under Different Precipitation Treatments
3.3. Composition of Soil Microbial Communities under Precipitation Treatments
3.4. Microbial β-Diversity Analysis under Treatments
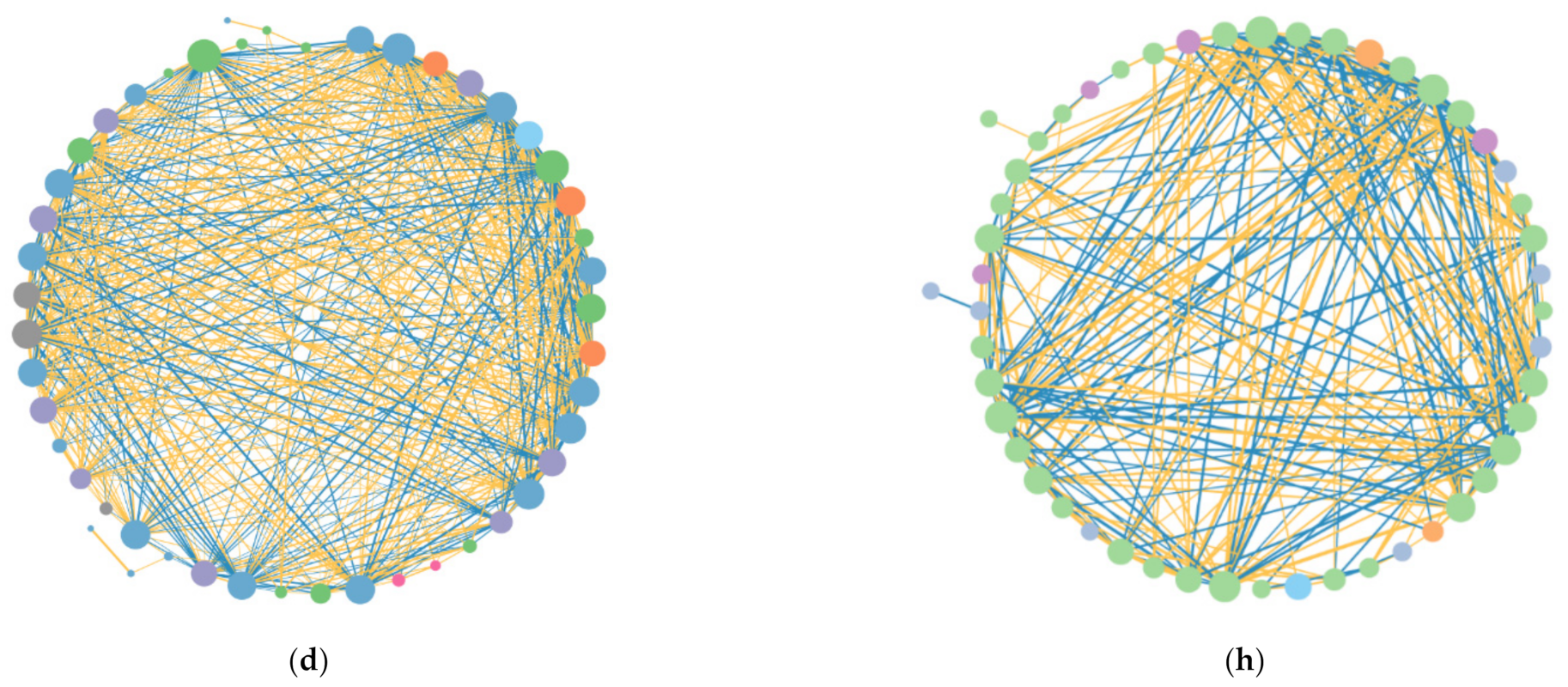
3.5. Interaction of Dominant Groups of Microorganisms under Treatments
3.6. Relationship between Environmental Factors and Community Composition under Different Treatments
4. Discussion
4.1. Response of Microbial Communities to Changes in Precipitation
4.2. Network Relationships Revealed the Bacterial-Fungal Interactions in Different Precipitation Treatments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Q.; Jiao, F.; Huang, Y.; Li, N.; Wang, B.; Gao, H.; An, S. Response of soil fungal community composition and functions on the alteration of precipitation in the grassland of Loess Plateau. Sci. Total Environ. 2021, 751, 142273. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Chen, L.; Ma, Z. Simulation of historical and projected climate change in arid and semiarid areas by CMIP5 models. Chin. Sci. Bull. 2014, 59, 412–429. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, P. Rainfall characteristics determine respiration rate of biological soil crusts in drylands. Ecol. Indic. 2021, 124, 107452. [Google Scholar] [CrossRef]
- Su, B.; Huang, J.; Fischer, T.; Wang, Y.; Kundzewicz, Z.W.; Zhai, J.; Sun, H.; Wang, A.; Zeng, X.; Wang, G.; et al. Drought losses in China might double between the 1.5 °C and 2.0 °C warming. Proc. Natl. Acad. Sci. USA 2018, 115, 10600–10605. [Google Scholar] [CrossRef] [Green Version]
- Sponseller, R.A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Glob. Chang. Biol. 2007, 13, 426–436. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–52. [Google Scholar] [CrossRef]
- Kang, H.; Yu, W.; Dutta, S.; Gao, H. Soil microbial community composition and function are closely associated with soil organic matter chemistry along a latitudinal gradient. Geoderma 2021, 383, 114744. [Google Scholar] [CrossRef]
- Ren, C.; Chen, J.; Lu, X.; Doughty, R.; Zhao, F.; Zhong, Z.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Responses of soil total microbial biomass and community compositions to rainfall reductions. Soil Biol. Biochem. 2018, 116, 4–10. [Google Scholar] [CrossRef]
- Li, J.; Shangguan, Z.; Deng, L. Dynamics of soil microbial metabolic activity during grassland succession after farmland abandonment. Geoderma 2020, 363, 114167. [Google Scholar] [CrossRef]
- Crowther, T.W.; van den Hoogen, J.; Wan, J.; Mayes, M.A.; Keiser, A.D.; Mo, L.; Averill, C.; Maynard, D.S. The global soil community and its influence on biogeochemistry. Science 2019, 365, eaav0550. [Google Scholar] [CrossRef]
- Beier, C.; Beierkuhnlein, C.; Wohlgemuth, T.; Penuelas, J.; Emmett, B.; Körner, C.; de Boeck, H.; Christensen, J.H.; Leuzinger, S.; Janssens, I.A.; et al. Precipitation manipulation experiments—challenges and recommendations for the future. Ecology Lett. 2012, 15, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Fry, E.L.; Manning, P.; Allen, D.G.; Hurst, A.; Everwand, G.; Rimmler, M.; Power, S.A. Plant functional group composition modifies the effects of precipitation change on grassland ecosystem function. PLoS ONE 2013, 8, e57027. [Google Scholar] [CrossRef] [Green Version]
- Na, X.; Yu, H.; Wang, P.; Zhu, W.; Niu, Y.; Huang, J. Vegetation biomass and soil moisture coregulate bacterial community succession under altered precipitation regimes in a desert steppe in northwestern China. Soil Biol. Biochem. 2019, 136, 107520. [Google Scholar] [CrossRef]
- Abbasi, A.O.; Salazar, A.; Oh, Y.; Reinsch, S.; del Rosario Uribe, M.; Li, J.; Rashid, I.; Dukes, J.S. Reviews and syntheses: Soil responses to manipulated precipitation changes—an assessment of meta-analyses. Biogeosciences 2020, 17, 3859–3873. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, K.; Li, Q.; Liu, X.; He, J.S.; Chu, H. Interannual climate variability and altered precipitation influence the soil microbial community structure in a Tibetan Plateau grassland. Sci. Total Environ. 2020, 714, 136794. [Google Scholar] [CrossRef] [PubMed]
- Kaisermann, A.; Maron, P.A.; Beaumelle, L.; Lata, J.C. Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Appl. Soil Ecol. 2015, 86, 158–164. [Google Scholar] [CrossRef]
- McHugh, T.A.; Schwartz, E. A watering manipulation in a semiarid grassland induced changes in fungal but not bacterial community composition. Pedobiologia 2016, 59, 121–127. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Physiological and community responses of established grassland bacterial populations to water stress. Appl. Environ. Microbiol. 2003, 69, 6961–6968. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, W.; Kang, X.; Cui, X.; Wang, Y.; Zhao, H.; Qian, X.; Hao, Y. Changes in soil microbial community response to precipitation events in a semi-arid steppe of the Xilin River Basin, China. J. Arid. Land 2018, 11, 97–110. [Google Scholar] [CrossRef] [Green Version]
- McHugh, T.A.; Schwartz, E. Changes in plant community composition and reduced precipitation have limited effects on the structure of soil bacterial and fungal communities present in a semiarid grassland. Plant Soil 2014, 388, 175–186. [Google Scholar] [CrossRef]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil microbial community responses to multiple experimental climate change drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Wang, B.; An, S.; Jiao, F.; Huang, Q. Response of Soil Bacterial Community Structure to Precipitation Change in Grassland of Loess Plateau. Environ. Sci. 2020, 41, 4284–4293. [Google Scholar] [CrossRef]
- Xi, N.; Bloor, J.M.G. Interactive effects of precipitation and nitrogen spatial pattern on carbon use and functional diversity in soil microbial communities. Appl. Soil Ecol. 2016, 100, 207–210. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Han, X.; Deng, Y. Higher precipitation strengthens the microbial interactions in semi-arid grassland soils. Glob. Ecol. Biogeogr. 2018, 27, 570–580. [Google Scholar] [CrossRef]
- Treves, D.S.; Xia, B.; Zhou, J.; Tiedje, J.M. A two-species test of the hypothesis that spatial isolation influences microbial diversity in soil. Microb. Ecol. 2003, 45, 20–28. [Google Scholar] [CrossRef]
- Jiao, P.; Li, Z.; Yang, L.; He, J.; Chang, X.; Xiao, H.; Nie, X.; Tong, D. Bacteria are more sensitive than fungi to moisture in eroded soil by natural grass vegetation restoration on the Loess Plateau. Sci. Total Environ. 2021, 756, 143899. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Chen, D.; Mi, J.; Chu, P.; Cheng, J.; Zhang, L.; Pan, Q.; Xie, Y.; Bai, Y. Patterns and drivers of soil microbial communities along a precipitation gradient on the Mongolian Plateau. Landsc. Ecol. 2014, 30, 1669–1682. [Google Scholar] [CrossRef]
- Cregger, M.A.; Schadt, C.W.; McDowell, N.G.; Pockman, W.T.; Classen, A.T. Response of the soil microbial community to changes in precipitation in a semiarid ecosystem. Appl. Environ. Microbiol. 2012, 78, 8587–8594. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Niu, D.; Gherardi, L.A.; Liu, Y.; Wang, Y.; Elser, J.J.; Fu, H. Linkages of stoichiometric imbalances to soil microbial respiration with increasing nitrogen addition: Evidence from a long-term grassland experiment. Soil Biol. Biochem. 2019, 138, 107580. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Rapson, G.; Ma, H.; Jing, L.; Zhang, Y.; Zhang, J.; Li, J. Increased precipitation enhances soil respiration in a semi-arid grassland on the Loess Plateau, China. PeerJ 2021, 9, e10729. [Google Scholar] [CrossRef] [PubMed]
- Kalembasa, S.J.; Jenkinson, D.S. A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Anal. Bioanal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.; Adams, A.F. Studies on soil organic matter: I. Influence of phosphorus content of parent materials on accumulations of carbon, nitrogen, sulfur, and organic phosphorus in grassland soils. Soil Sci. 1958, 85, 307–318. [Google Scholar] [CrossRef]
- Le Bissonnais, Y. Aggregate stability and assessment of soil crustability and erodibility: I. Theory and methodology. Eur. J. Soil Sci. 2016, 47, 425–437. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glockner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C.; Flyvbjerg, H. Octave Plots for Visualizing Diversity of Microbial OTUs. Available online: https://www.biorxiv.org/content/10.1101/389833v1 (accessed on 11 August 2018).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0. Available online: http://library.wur.nl/WebQuery/wurpubs/431861 (accessed on 21 February 2022).
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, U.N.; Ball, B.A. Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi-arid ecosystems. Glob. Chang. Biol. 2015, 21, 1407–1421. [Google Scholar] [CrossRef]
- Pan, Y.; Kang, P.; Hu, J.; Song, N. Bacterial community demonstrates stronger network connectivity than fungal community in desert-grassland salt marsh. Sci. Total Environ. 2021, 798, 149118. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, H.; Wang, C.; Xu, J.R. Comparative analysis of fungal genomes reveals different plant cell wall degrading capacity in fungi. BMC Genom. 2013, 14, 274. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xu, T.-L.; Veresoglou, S.D.; Hu, H.-W.; Hao, Z.-P.; Hu, Y.-J.; Liu, L.; Deng, Y.; Rillig, M.C.; Chen, B.-D. Plant diversity represents the prevalent determinant of soil fungal community structure across temperate grasslands in northern China. Soil Biol. Biochem. 2017, 110, 12–21. [Google Scholar] [CrossRef]
- Che, R.; Wang, S.; Wang, Y.; Xu, Z.; Wang, W.; Rui, Y.; Wang, F.; Hu, J.; Tao, J.; Cui, X. Total and active soil fungal community profiles were significantly altered by six years of warming but not by grazing. Soil Biol. Biochem. 2019, 139, 107611. [Google Scholar] [CrossRef]
- Guan, Y.; Jiang, N.; Wu, Y.; Yang, Z.; Bello, A.; Yang, W. Disentangling the role of salinity-sodicity in shaping soil microbiome along a natural saline-sodic gradient. Sci. Total Environ. 2021, 765, 142738. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Shannon-Wiener | Pielou | Patrick | Coverage | Above-Ground Biomass (g m−2) | Litter Biomass (g m−2) | Below-Ground Biomass (g m−2) |
|---|---|---|---|---|---|---|---|
| P50 | 1.46 ± 0.11b | 0.88 ± 0.03a | 5.33 ± 0.58b | 0.65 ± 0.15b | 152.74 ± 68.65a | 96.41 ± 37.43a | 40.54 ± 16.84b |
| P100 | 2.35 ± 0.08a | 0.84 ± 0.01ab | 16.33 ± 1.15a | 0.89 ± 0.02a | 245.59 ± 49.07a | 87.19 ± 32.61a | 117.28 ± 25.89a |
| P150 | 2.29 ± 0.13a | 0.81 ± 0.02b | 17 ± 2.65a | 0.98 ± 0.01a | 276.69 ± 55.95a | 88.30 ± 32.51a | 56.43 ± 19.95b |
| Treatment | 0–10 cm | 10–20 cm | 20–30 cm | p Value from Two-Way ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P50 | P100 | P150 | P50 | P100 | P150 | P50 | P100 | P150 | L | P | L × P | |
| C (g kg−1) | 16.72 ± 1.34 | 18.22 ± 1.57 | 18.76 ± 3.05 | 13.89 ± 3.81 | 15.29 ± 1.35 | 18.07 ± 2.92 | 14.05 ± 1.70 | 16.31 ± 2.28 | 13.80 ± 1.73 | 0.03 | 0.18 | 0.38 |
| N (g kg−1) | 2.21 ± 0.08 | 2.12 ± 0.16 | 2.19 ± 0.05 | 2.29 ± 0.12 | 2.24 ± 0.09 | 2.15 ± 0.09 | 2.28 ± 0.29 | 2.10 ± 0.04 | 1.89 ± 0.25 | 0.19 | 0.07 | 0.35 |
| P (g kg−1) | 0.74 ± 0.15 | 0.82 ± 0.00 | 0.79 ± 0.05 | 0.85 ± 0.02 | 0.85 ± 0.03 | 0.78 ± 0.02 | 0.83 ± 0.01 | 0.87 ± 0.03 | 0.76 ± 0.01 | 0.28 | 0.05 | 0.28 |
| AP (mg kg−1) | 4.80 ± 1.38 | 3.68 ± 0.89 | 4.45 ± 0.32 | 3.86 ± 1.61 | 2.85 ± 0.32 | 3.49 ± 0.79 | 2.45 ± 0.44 | 2.49 ± 0.27 | 2.99 ± 0.95 | 0.00 | 0.22 | 0.79 |
| pH | 7.78 ± 0.05 | 7.84 ± 0.04 | 7.72 ± 0.08 | 7.83 ± 0.08 | 8.02 ± 0.15 | 7.91 ± 0.08 | 7.87 ± 0.08 | 8.06 ± 0.04 | 7.98 ± 0.22 | 0.00 | 0.02 | 0.63 |
| SM (%) | 5.20 ± 0.17 | 7.29 ± 1.22 | 8.88 ± 0.10 | 6.94 ± 1.41 | 8.71 ± 0.40 | 8.17 ± 1.49 | 4.76 ± 0.43 | 8.64 ± 1.16 | 9.05 ± 0.38 | 0.20 | 0.00 | 0.046 |
| SFD | 2.79 ± 0.03 | 2.77 ± 0.05 | 2.78 ± 0.06 | 2.87 ± 0.05 | 2.80 ± 0.08 | 2.75 ± 0.03 | 2.81 ± 0.05 | 2.78 ± 0.11 | 2.82 ± 0.08 | 0.67 | 0.37 | 0.44 |
| MWD | 1.73 ± 0.19 | 1.74 ± 0.37 | 1.78 ± 0.34 | 1.24 ± 0.37 | 1.58 ± 0.65 | 1.89 ± 0.07 | 1.71 ± 0.25 | 1.67 ± 0.52 | 1.59 ± 0.45 | 0.63 | 0.58 | 0.55 |
| GMD | 1.22 ± 0.04 | 1.22 ± 0.09 | 1.32 ± 0.08 | 1.11 ± 0.08 | 1.19 ± 0.15 | 1.26 ± 0.02 | 1.21 ± 0.06 | 1.21 ± 0.13 | 1.19 ± 0.11 | 0.66 | 0.56 | 0.56 |
| Compactness (Pa) | 1397.30 ± 336.67 | 705.67 ± 86.16 | 943.08 ± 331.02 | 1296.58 ± 610.82 | 580.92 ± 75.13 | 914.33 ± 361.56 | 1466.17 ± 592.27 | 808.00 ± 189.21 | 1259.25 ± 384.18 | 0.39 | 0.00 | 0.98 |
| Network Topological Features | Bacterial | Fungal | ||||
|---|---|---|---|---|---|---|
| P50 | P100 | P150 | P50 | P100 | P150 | |
| Nodes | 49 | 49 | 48 | 47 | 47 | 50 |
| Total Link | 506 | 535 | 498 | 165 | 233 | 285 |
| Positive link | 332 | 294 | 297 | 100 | 119 | 154 |
| Negative link | 174 | 241 | 201 | 65 | 114 | 131 |
| Average degree | 20.653 | 21.837 | 20.75 | 7.021 | 10.311 | 11.4 |
| Transitivity | 0.865 | 0.784 | 0.769 | 0.497 | 0.524 | 0.603 |
| Clustering coefficient | 0.813 | 0.765 | 0.688 | 0.461 | 0.527 | 0.522 |
| Network Density | 0.43 | 0.455 | 0.441 | 0.153 | 0.234 | 0.233 |
| Network heterogeneity | 0.517 | 0.482 | 0.496 | 0.523 | 0.511 | 0.608 |
| Network centralization | 0.225 | 0.264 | 0.294 | 0.181 | 0.254 | 0.289 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Xie, Y.; Ma, H.; Zhang, Y.; Zhang, J.; Zhang, H.; Luo, X.; Li, J. Responses of Soil Microbial Communities and Networks to Precipitation Change in a Typical Steppe Ecosystem of the Loess Plateau. Microorganisms 2022, 10, 817. https://doi.org/10.3390/microorganisms10040817
Wang Y, Xie Y, Ma H, Zhang Y, Zhang J, Zhang H, Luo X, Li J. Responses of Soil Microbial Communities and Networks to Precipitation Change in a Typical Steppe Ecosystem of the Loess Plateau. Microorganisms. 2022; 10(4):817. https://doi.org/10.3390/microorganisms10040817
Chicago/Turabian StyleWang, Yutao, Yingzhong Xie, Hongbin Ma, Yi Zhang, Juan Zhang, Hao Zhang, Xu Luo, and Jianping Li. 2022. "Responses of Soil Microbial Communities and Networks to Precipitation Change in a Typical Steppe Ecosystem of the Loess Plateau" Microorganisms 10, no. 4: 817. https://doi.org/10.3390/microorganisms10040817