
Colonization of Dogs and Their Owners with Staphylococcus aureus and Staphylococcus pseudintermedius in Households, Veterinary Practices, and Healthcare Facilities

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Populations and Recruitment
2.2. Microbiological Analysis
2.3. Molecular Characterization
2.4. Whole-Genome Sequencing and Phylogenetic Analyses
3. Results
3.1. Cohort I: Humans and Dogs in Household Contacts
3.1.1. S. aureus Nasal Carriage
3.1.2. S. aureus Intrahousehold Transmission
3.1.3. S. pseudintermedius Colonization of Dogs and Humans
3.2. Cohort II: Dogs and Humans after Visiting a Veterinarian Practice
3.3. Cohort III: Dogs Active in AAI and Their Owners
3.4. Results from Spa-Typing and Phenotypic Susceptibility Testing of S. aureus (Cohorts I–III)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sakr, A.; Brégeon, F.; Mège, J.L.; Rolain, J.M.; Blin, O. Staphylococcus aureus nasal colonization: An update on mechanisms, epidemiology, risk factors, and subsequent infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef] [PubMed]
- Mehraj, J.; Witte, W.; Akmatov, M.K.; Layer, F.; Werner, G.; Krause, G. Epidemiology of Staphylococcus aureus nasal carriage patterns in the community. Curr. Top. Microbiol. Immunol. 2016, 398, 55–87. [Google Scholar]
- Cuny, C.; Friedrich, A.; Kozytska, S.; Layer, F.; Nübel, U.; Ohlsen, K.; Strommenger, B.; Walther, B.; Wieler, L.; Witte, W. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) in different animal species. Int. J. Med. Microbiol. 2010, 300, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Witte, W.; Cuny, C.; Klare, I.; Nübel, U.; Strommenger, B.; Werner, G. Emergence and spread of antibiotic-resistant Gram-positive bacterial pathogens. Int. J. Med. Microbiol. 2008, 298, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, J.R.; Chen, L.; Mathema, B.; Kreiswirth, B.N. Global epidemiology of community-associated methicillin resistant Staphylococcus aureus (CA-MRSA). Curr. Opin. Microbiol. 2012, 15, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Wieler, L.H.; Friedrich, A.W.; Kohn, B.; Brunnberg, L.; Lübke-Becker, A. Staphylococcus aureus and MRSA colonization rates among personnel and dogs in a small animal hospital: Association with nosocomial infections. Berl. Munch. Tierarztl. Wochenschr. 2009, 122, 178–185. [Google Scholar]
- Vincze, S.; Stamm, I.; Kopp, P.A.; Hermes, J.; Adlhoch, C.; Semmler, T.; Wieler, L.H.; Lübke-Becker, A.; Walther, B. Alarming proportions of methicillin-resistant Staphylococcus aureus (MRSA) in wound samples from companion animals, Germany 2010–2012. PLoS ONE 2014, 9, e85656. [Google Scholar] [CrossRef] [Green Version]
- Abdelbary, M.M.; Wittenberg, A.; Cuny, C.; Layer, F.; Kurt, K.; Wieler, L.H.; Walther, B.; Skov, R.; Larsen, J.; Hasman, H.; et al. Phylogenetic analysis of Staphylococcus aureus CC398 reveals a sub-lineage epidemiologically associated with infections in horses. PLoS ONE 2014, 9, e88083. [Google Scholar] [CrossRef] [Green Version]
- Cuny, C.; Witte, W. MRSA in equine hospitals and its significance for infections in humans. Vet. Microbiol. 2017, 200, 59–64. [Google Scholar] [CrossRef]
- Fitzgerald, J.R. The Staphylococcus intermedius group of bacterial pathogens: Species re-classification, pathogenesis and the emergence of methicillin resistance. Vet. Dermatol. 2009, 20, 490–495. [Google Scholar] [CrossRef]
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Pires dos Santos, T.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic review on global epidemiology of methicillin-resistant Staphylococcus pseudintermedius: Inference of population structure from multilocus sequence typing data. Front. Microbiol. 2016, 7, 1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talan, D.A.; Goldstein, E.; Staatz, D.; Overturf, G.D. Staphylococcus intermedius: Clinical presentation of a new human dog bite pathogen. Ann. Emerg. Med. 1989, 18, 410–413. [Google Scholar] [CrossRef]
- Börjesson, S.; Gomez-Sanz, E.; Ekstrom, K.; Torres, C.; Gronlund, U. Staphylococcus pseudintermedius can be misdiagnosed as Staphylococcus aureus in humans with dog bite wounds. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 839–844. [Google Scholar] [CrossRef]
- Somayaji, R.; Priyantha, M.A.; Rubin, J.E.; Church, D. Human infections due to Staphylococcus pseudintermedius, anemerging zoonosis of canine origin: Report of 24 cases. Diagn. Microbiol. Infect. Dis. 2016, 85, 471–476. [Google Scholar] [CrossRef]
- Han, J.I.; Yang, C.H.; Park, H.M. Prevalence and risk factors of Staphylococcus spp. carriage among dogs and their owners: A cross-sectional study. Vet. J. 2016, 212, 15–21. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Belas, A.; Marques, C.; Cruz, L.; Gama, L.T.; Pomba, C. Similar articles risk factors for nasal colonization by methicillin-resistant staphylococci in healthy humans in professional daily contact with companion animals in Portugal. Microb. Drug. Resist. 2018, 24, 434–446. [Google Scholar] [CrossRef]
- German federal Ministry of Food and Agriculture. Available online: https://www.tierwohl-staerken.de/heimtiere/heimtiere-in-deutschland/# (accessed on 21 February 2022).
- Guardabassi, L.; Schwarz, S.; Lloyd, D. Pet animals as reservoirs of antimicrobial-resistant bacteria. J. Antimicrob. Chemother. 2004, 54, 321–332. [Google Scholar] [CrossRef]
- Mann, P.H. Bacteriophage typing of staphylococci isolated from animals. Canad. J. Pub. Health. 1960, 51, 153–156. [Google Scholar]
- Kaspar, U.; Von Lützau, A.; Schlattmann, A.; Roesler, U.; Köck, R.; Becker, K. Zoonotic multidrug-resistant microorganisms among small companion animals in Germany. PLoS ONE 2018, 13, e0208364. [Google Scholar] [CrossRef] [Green Version]
- Boost, M.; O`Donoghue, M.; James, A. Investigation of the role of dogs as reservoirs of Staphylococcus aureus and the transmission of strains between pet owners and their dogs. Hong Kong Med. J. 2008, 14, 15–18. [Google Scholar] [PubMed]
- Van Balen, J.C.; Landers, T.; Nutt, E.; Dent, A.; Hoet, A.E. Molecular epidemiological analysis to assess the influence of pet-ownership in the biodiversity of Staphylococcus aureus and MRSA in dog- and non-dog-owning healthy household. Epidemiol. Infect. 2017, 145, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, D.O.; Lautenbach, E.; Zaoutis, T.; Leckerman, K.; Edelstein, P.H.; Rankin, S.C. Potential for pet animals to harbour methicillin-resistant Staphylococcus aureus (MRSA) when residing with human MRSA patients. Zoonoses Public Health 2012, 59, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.B.; Waters, K.C.; Nerby, J.; Olsen, K.E.; Jawahi, S. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from pets living in households with MRSA-infected children. Clin. Infect. Dis. 2012, 54, 49–50. [Google Scholar] [CrossRef] [Green Version]
- Loeffler, A.; McCarthy, A.; Lloyd, D.H.; Musilová, E.; Pfeiffer, D.U.; Lindsay, J.A. Whole-genome com-parison of methicillin-resistant Staphylococcus aureus CC22 SCCmecIV from people and their in-contact pets. Vet. Dermatol. 2013, 24, 538-e128. [Google Scholar] [CrossRef]
- Hanselman, B.A.; Kruth, S.A.; Rousseau, J.; Weese, J.S. Coagulase positive staphylococcal colonization of humans and their household pet. Can. Vet. J. 2009, 50, 954–958. [Google Scholar]
- Faires, M.C.; Tater, K.; Weese, J.S. An investigation of methicillin-resistant Staphylococcus aureus colonization in people and pets in the same household with an infected person or infected pet. J. Am. Vet. Med. Ass. 2009, 235, 540–543. [Google Scholar] [CrossRef] [Green Version]
- Lundqvist, M.; Carlsson, P.; Sjödahl, R.; Theodorsson, E.; Levin, L.Å. Patient benefit of dog-assisted interventions in health care: A systematic review. BMC Complementary Altern. Med. 2017, 17, 358. [Google Scholar] [CrossRef]
- Hediger, K.; Wagner, J.; Künzi, K.; Haefeli, A.; Theis, F.; Grob, C.; Pauli, J.; Gerger, H. Effectiveness of animal-assisted interventions for children and adults with post-traumatic stress disorder symptoms: A systematic review and meta-analysis. Eur. J. Psychotraumatol. 2021, 12, 1879713. [Google Scholar] [CrossRef]
- Lefebvre, S.L.; Waltner-Toews, D.; Peregrine, A.S.; Reid-Smith, R.; Hodge, L.; Arroyo, L.; Weese, J.S. Prevalence of zoonotic agents in dogs visiting hospitalized people in Ontario: Implications for infection control. J. Hosp. Infect. 2006, 62, 458–466. [Google Scholar] [CrossRef]
- Enoch, D.A.; Karas, J.A.; Slater, J.D.; Emery, M.M.; Kearns, A.M.; Farrington, M. MRSA carriage in a pet therapy dog. J. Hosp. Infect. 2005, 60, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Akmatov, M.K.; Mehraj, J.; Gatzemeier, A.; Strömp, J.; Witte, W.; Krause, G.; Pessler, F. Serial home-based self-collection of anterior nasal swabs to detect Staphylococcus aureus carriage in a randomized population-based study in Germany. Int. J. Infect. Dis. 2014, 25, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martineau, F.; Picard, F.J.; Ke, D.; Paradis, S.; Roy, P.H.; Ouellette, M.; Bergeron, M.G. Development of a PCR assay for identification of staphylococci at genus and species levels. J. Clin. Microbiol. 2001, 39, 2541–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Tsubakishita, S.; Tanaka, Y.; Sakusabe, A.; Ohtsuka, M.; Hirotaki, S.; Kawakami, T.; Fukata, T.; Hiramatsu, K. Multiplex-PCR method for species identification of coagulase-positive staphylococci. J. Clin. Microbiol. 2010, 48, 765–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuny, C.; Layer, F.; Hansen, S.; Werner, G.; Witte, W. Nasal Colonization of humans with occupational exposure to raw meat and to raw meat products with methicillin-susceptible and methicillin-resistant Staphylococcus aureus. Toxins 2019, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Goyal, M.; Javerliat, F.; Palmieri, M.; Mirande, C.; Van Wamel, W.; Tavakol, M.; Verkaik, N.; Van Belkum, A. Genomic evolution of Staphylococcus aureus during artificial and natural colonization of the human nose. Front. Microbiol. 2019, 10, 1525. [Google Scholar] [CrossRef]
- Mehraj, J.; Akmatov, M.K.; Strömpl, J.; Gatzemeier, A.; Layer, F.; Werner, G.; Pieper, D.H.; Medina, E.; Witte, W.; Pessler, F.; et al. Methicillin-sensitive and methicillin-resistant Staphylococcus aureus nasal carriage in a random sample of non-hospitalized adult population in northern Germany. PLoS ONE 2014, 9, e107937. [Google Scholar] [CrossRef]
- Holtfreter, S.; Grumann, D.; Balau, V.; Barwich, A.; Kolata, J.; Goehler, A.; Weiss, S.; Holtfreter, B.; Bauerfeind, S.S.; Döring, P.; et al. Molecular epidemiology of Staphylococcus aureus in the general population in Northeast Germany: Results of the study of health in Pomerania (SHIP-TREND-0). J. Clin. Microbiol. 2016, 54, 2774–2785. [Google Scholar] [CrossRef] [Green Version]
- Misic, A.M.; Davis, M.F.; Tyldsley, A.S.; Hodkinson, B.P.; Tolomeo, P.; Hu, B.; Nachamkin, I.; Lautenbach, E.; Morris, D.O.; Grice, E.A. The shared microbiota of humans and companion animals as evaluated from Staphylococcus carriage sites. Microbiome 2015, 3, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Hanson, B.M.; Kates, A.E.; O’Malley, S.M.; Mills, E.; Herwaldt, L.A.; Torner, J.C.; Dawson, J.D.; Farina, S.A.; Klostermann, C.; Wu, J.Y.; et al. Staphylococcus aureus in the nose and throat of Iowan families. Epidemiol. Infect. 2018, 14, 1777–1784. [Google Scholar] [CrossRef] [Green Version]
- Knox, J.; Uhlemann, A.C.; Lowy, F.D. Staphylococcus aureus infections: Transmission within households and the community. Trends Microbiol. 2015, 7, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Torres, C.; Lozano, C.; Zarazaga, M. High diversity of Staphylococcus aureus and Staphylococcus pseudintermedius lineages and toxigenic traits in healthy pet-owning household members. Underestimating normal household contact? Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Kottler, S.; Middleton, J.R.; Perry, J.; Weese, J.S.; Cohn, L.A. Prevalence of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus carriage in three populations. J. Vet. Intern. Med. 2010, 24, 132–139. [Google Scholar] [CrossRef]
- Layer, F.; Stromenger, B.; Cuny, C.; Werner, G. Eigenschaften, Häufigkeit und Verbreitung von MRSA in Deutschland—Update 2013/2014. Epidemiol. Bull. 2015. Available online: https://www.rki.de/DE/Content/Infekt/EpidBull/Archiv/2015/31/Art_01.html (accessed on 21 February 2022).
- Paterson, G.K.; Harrison, E.M.; Holmes, M.A. The emergence of mecC methicillin-resistant Staphylococcus aureus. Trends Microbiol. 2014, 22, 42–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenni, M.; El Garch, F.; Miossec, C.; Madec, J.Y.; Hocquet, D.; Valot, B. High genetic diversity among methicillin-susceptible Staphylococcus pseudintermedius in dogs in Europe. J. Glob. Antimicrob. Resist. 2020, 21, 57–59. [Google Scholar] [CrossRef]
- Phumthanakorn, N.; Schwendener, S.; Donà, V.; Chanchaithong, P.; Perreten, V.; Prapasarakul, N. Genomic insights into methicillin-resistant Staphylococcus pseudintermedius isolates from dogs and humans of the same sequence types reveals diversity in prophages and pathogenicity islands. PLoS ONE 2021, 16, e0254382. [Google Scholar] [CrossRef]
- Kjellman, E.E.; Slettemeås, J.S.; Small, H.; Sunde, M. Methicillin-resistant Staphylococcus pseudintermedius (MRSP) from healthy dogs in Norway-occurrence, genotypes and comparison to clinical MRSP. Microbiologyopen 2015, 4, 857–866. [Google Scholar] [CrossRef]
- Haenni, M.; de Moraes, N.A.; Châtre, P.; Médaille, C.; Moodley, A.; Madec, J.Y. Characterisation of clinical canine methicillin-resistant and methicillin-susceptible Staphylococcus pseudintermedius in France. J. Glob. Antimicrob. Resist. 2014, 2, 119–123. [Google Scholar] [CrossRef]
- Duim, B.; Verstappen, K.M.; Broens, E.M.; Laarhoven, L.M.; van Duijkeren, E.; Hordijk, J.; de Heus, P.; Spaninks, M.; Timmerman, A.J.; Wagenaar, J.A. Changes in the population of methicillin-resistant Staphylococcus pseudintermedius and dissemination of antimicrobial-resistant phenotypes in the Netherlands. J. Clin. Microbiol. 2016, 54, 283–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viau, R.; Hujer, A.M.; Hujer, K.M.; Bonomo, R.A.; Jump, R.L. Are Staphylococcus intermedius Infections in Humans Cases of Mistaken Identity? A Case Series and Literature Review. Open Forum Infect. Dis. 2015, 2, ofv110. [Google Scholar] [CrossRef] [Green Version]
- Bert, F.; Gualano, M.R.; Camussi, E.; Pieve, G.; Voglino, G.; Siliquini, R. Animal assisted intervention: A systematic review of benefits and risks. Eur. J. Integr. Med. 2016, 8, 695–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, K.R.; Ruble, K.; Redding, L.E.; Morris, O.D.; Mueller-Roland, N.T.; Thorpe, J.; Agnew, J.; Carroll, K.C.; Planet, P.J.; Rubenstein, R.C.; et al. Microbial sharing between pediatric patients and therapy dogs during hospital animal-assisted intervention programs. Microorganisms 2021, 9, 1054. [Google Scholar] [CrossRef]
- Murthy, R.; Bearman, G.; Brown, S.; Bryant, K.; Chinn, R.; Hewlett, A.; George, B.G.; Goldstein, E.J.C.; Holzmann-Pazgal, G.; Rupp, M.E.; et al. Animals in healthcare facilities: Recommendations to minimize potential risks. Infect. Control. Hosp. Epidemiol. 2015, 36, 495–516. [Google Scholar] [CrossRef] [Green Version]
- Deutsche Gesellschaft für Krankenhaushygiene. Empfehlung zum hygienegerechten Umgang mit Therapiehunden in Krankenhäusern und vergleichbaren Einrichtungen. Hyg. Med. 2017, 42-10. Available online: https://www.krankenhaushygiene.de/pdfdata/hm/2017_10_DGKH_Therapiehunde.pdf (accessed on 21 February 2022).
- Caffrey, A.R.; Quilliam, B.J.; LaPlante, K. Risk factors associated with mupirocin resistance in methicillin-resistant Staphylococcus aureus. J. Hosp. Infect. 2010, 76, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Kizerwetter-Świda, M.; Chrobak-Chmiel, D.; Rzewuska, M. High-level mupirocin resistance in methicillin-resistant staphylococci isolated from dogs and cats. BMC Vet. Res. 2019, 15, 238. [Google Scholar] [CrossRef]




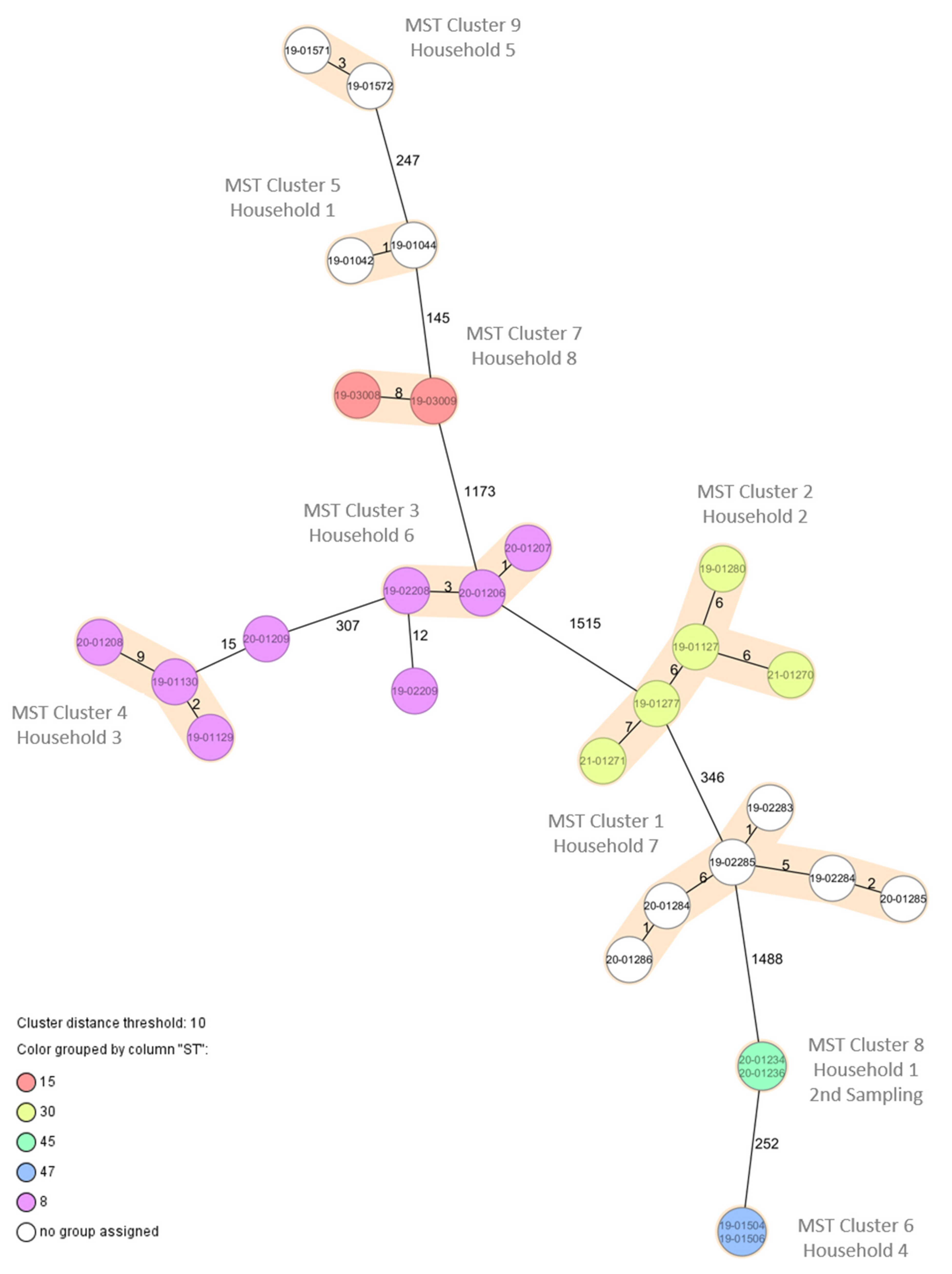{kind=link}
{kind=link}
{kind=link}
| Characteristics | All 179 Persons in 84 Households 1 | 14 Persons in 8 Households with S. aureus in Humans and Dogs | 97 Persons in 34 Households with S. aureus only in Humans | 68 Persons in 42 Households without S. aureus | |||
|---|---|---|---|---|---|---|---|
| carriage | SA positive | SA negative | SA positive | SA negative | SA positive | SA negative | SA negative |
| No. and % among carriers and non-carriers, respectively | |||||||
| all | 66 (36.9) | 113 (63.1) | 12 | 2 | 54 (55.6) | 43 (44.3) | 68 (100.0) |
| female | 34 (51.5) | 61 (54.0) | 8 | 1 | 26 (48.1) | 25 (58.1) | 35 (51.4) |
| male | 32 (48.5) | 52 (46.0) | 4 | 1 | 28 (51.9) | 18 (41.9) | 33 (48.5) |
| previous hospital stay 2 | 11 (16.7) | 15 (13.3) | 3 | 2 | 8 (14.8) | 4 (9.3) | 9 (13.2) |
| antibiotic consumption 2 | 12 (18.2) | 12 (10.6) | 3 | 0 | 9 (16.7) | 4 (9.3) | 8 (11.8) |
| diabetes mellitus | 4 (6.1) | 3 (2.7) | 0 | 0 | 4 (7.4) | 0 | 3 (4.4) |
| skin lesions | 2 (3.0) | 2 (1.8) | 0 | 0 | 2 3.7) | 2 (4.7) | 0 |
| atopic eczema | 10 (15.2) | 4 (3.5%) | 1 | 0 | 0 | 0 | 4 (5.9) |
| ≥2 dogs in the household | 25 (37.8%) | 36 (31.8%) | 5 | 3 | 20 (37) | 16 (37.2%) | 17 (25%) |
| Persons/ Household | No. of Households (n = 84) | Households with Carriers of S. aureus (n =42) | Households with 1 Person Colonized | Households with ≥ 2 Persons Colonized | ||||
|---|---|---|---|---|---|---|---|---|
| Households | Carrier (%) | All persons | Households | Carrier (%) | All persons | |||
| 1 | 18 | 4 | 4 | 4 (22) | 18 | |||
| ≥2 | 66 | 38 1 | 21 | 21 (50) | 42 | 17 | 41 (82) 2 | 51 |
| First Sampling | Second Sampling | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Household | Individual 1 | Species 2 | Isolate 3 | Spa-Type | CC 4 | Resistance Phenotype 5 | Individual 1 | Species 2 | Isolate 3 | Spa-Type | CC 4 | Resistance Phenotype 5 |
| 1 | H-28 | SA | 19-01042 | t2696 | 15 | PEN | H-28.1 | SA | 20-01234 | t779 | 45 | susceptible |
| H-29 | SA | t091 | 7 | susceptible | H-29.1 | SA | t091 | 7 | susceptible | |||
| D-30 | SA | 19-01044 | t2696 | 15 | susceptible | D-30.1 | negative | |||||
| D-400 | SA | 20-01236 | t779 | 45 | susceptible | |||||||
| 2 | H-62 | SA | 19-01277 | t1577 | 30 | PEN, ERY | H-62.1 | SA | 21-01270 | t1577 | 30 | PEN, ERY |
| H-63 | SA | 19-01127 | t1577 | 30 | PEN, ERY | H-63.1 | SA | 21-01271 | t1577 | 30 | PEN, ERY | |
| D-64 | SP | PEN, OXA, TET, CIP, MOX | D-64.1 | negative | ||||||||
| D-65 | SA | 19-01280 | t1577 | 30 | PEN, ERY | D-65.1 | negative | |||||
| 3 | H-66 | SA | 19-01129 | t9325 | 8 | PEN | H-66.1 | SA | 20-01208 | t9325 | 8 | PEN |
| D-67 | SA | 19-01130 | t9325 | 8 | PEN | D-67.1 | SA, SP | 20-01209 | t9325 | 8 | PEN | |
| 4 | H-176 | SA | 19-01504 | t026 | 45 | susceptible | H-176.1 | negative | ||||
| D-178 | SP | D-178.1 | negative | |||||||||
| D-179 | SA | 19-01506 | t026 | 45 | susceptible | D-179.1 | negative | |||||
| 5 | H-206 | SA | 19-01571 | t084 | 15 | PEN | H-206.1 | No feedback | ||||
| H-207 | negative | H-207.1 | ||||||||||
| D-208 | SA | 19-01572 | t084 | 15 | PEN | |||||||
| 6 | H-312 | SA | 19-02208 | t008 | 8 | PEN, FUS | D-312.1 | SA | 20-01206 | t008 | 8 | PEN, FUS |
| D-313 | SA | 19-02209 | t008 | 8 | PEN, FUS | D-313.1 | SA | 20-01207 | t008 | 8 | PEN, FUS | |
| D-314 | SP | susceptible | deceased | |||||||||
| 7 | H-318 | SA | 19-02283 | t6997 | 30 | PEN | H-318.1 | SA | 20-01284 | t6997 | 30 | PEN |
| H-319 | SA | 19-02284 | t6997 | 30 | PEN | H-319.1 | SA | 20-01286 | t6997 | 30 | PEN | |
| D-320 | SA | 19-02285 | t6997 | 30 | PEN | D-320.1 | SA | t6997 | 30 | PEN | ||
| 8 | H-350 | negative | No feedback | |||||||||
| H-351 | SA | 19-03008 | t935 | 15 | susceptible | H-351.1 | ||||||
| D-352 | SP | TET | D-352.1 | |||||||||
| D-353 | SA | 19-03009 | t935 | 15 | susceptible | D-353.1 | ||||||
| Total No. of Households | No. of Households with SP 1 Positive Dogs | Total No. of Dogs | No. of SP 1 Positive Dogs | ||
|---|---|---|---|---|---|
| All | 84 | 36 | 112 | 42 (37.5%) | |
| Households with 1 dog | 61 | 21 (34.4%) | 61 | 21 (34.4%) | |
| Households with ≥2 dogs | 23 | 15 (65.2%) p (0.078) | 51 | 21 (41.2%) p (0.557) | |
| 2 dogs | 19 | 11 | 38 | 14 (36.8%) | |
| 3 dogs | 3 | 3 | 9 | 4 | |
| 4 dogs | 1 | 1 | 4 | 3 | |
| Antibiotic Resistance Phenotypes 1 | All | Cohort I | Cohort II | Cohort III |
|---|---|---|---|---|
| PEN | 8 | 7 | 1 | |
| TET | 4 | 4 | ||
| PEN, TET | 5 | 5 | ||
| ERY, CLI | 3 | 1 | 1 | 1 |
| ERY, CLI, TET | 3 | 2 | 1 | |
| PEN, ERY, CLI, TET | 3 | 3 | ||
| PEN, GEN, ERY, CLI, TET, CIP | 1 | 1 | ||
| PEN, OXA, TET, CIP, MOX | 1 | 1 | ||
| PEN, OXA, ERY, CLI, TET, CIP, MOX, TRS | 3 | 3 | ||
| PEN, OXA, GEN, ERY, CLI, CIP, MOX, TRS, FUS | 1 | 1 | ||
| Susceptible to all antibiotics tested | 23 | 18 | 1 | 4 |
| Total | 55 | 42 | 6 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuny, C.; Layer-Nicolaou, F.; Weber, R.; Köck, R.; Witte, W. Colonization of Dogs and Their Owners with Staphylococcus aureus and Staphylococcus pseudintermedius in Households, Veterinary Practices, and Healthcare Facilities. Microorganisms 2022, 10, 677. https://doi.org/10.3390/microorganisms10040677
Cuny C, Layer-Nicolaou F, Weber R, Köck R, Witte W. Colonization of Dogs and Their Owners with Staphylococcus aureus and Staphylococcus pseudintermedius in Households, Veterinary Practices, and Healthcare Facilities. Microorganisms. 2022; 10(4):677. https://doi.org/10.3390/microorganisms10040677
Chicago/Turabian StyleCuny, Christiane, Franziska Layer-Nicolaou, Robert Weber, Robin Köck, and Wolfgang Witte. 2022. "Colonization of Dogs and Their Owners with Staphylococcus aureus and Staphylococcus pseudintermedius in Households, Veterinary Practices, and Healthcare Facilities" Microorganisms 10, no. 4: 677. https://doi.org/10.3390/microorganisms10040677