
Physico-Chemical and Metagenomic Profile Analyses of Animal Manures Routinely Used as Inocula in Anaerobic Digestion for Biogas Production




Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Feed and Manure Sample Collection
2.2. Physico-Chemical Analysis of Animal Feeds and Manures
2.3. Microbiome Characterisation
DNA Extraction and Bioinformatics Analysis
3. Results
3.1. Elemental Composition of Animal Feeds and Manures
3.2. Taxonomic Diversity of Bacterial Communities in Manure
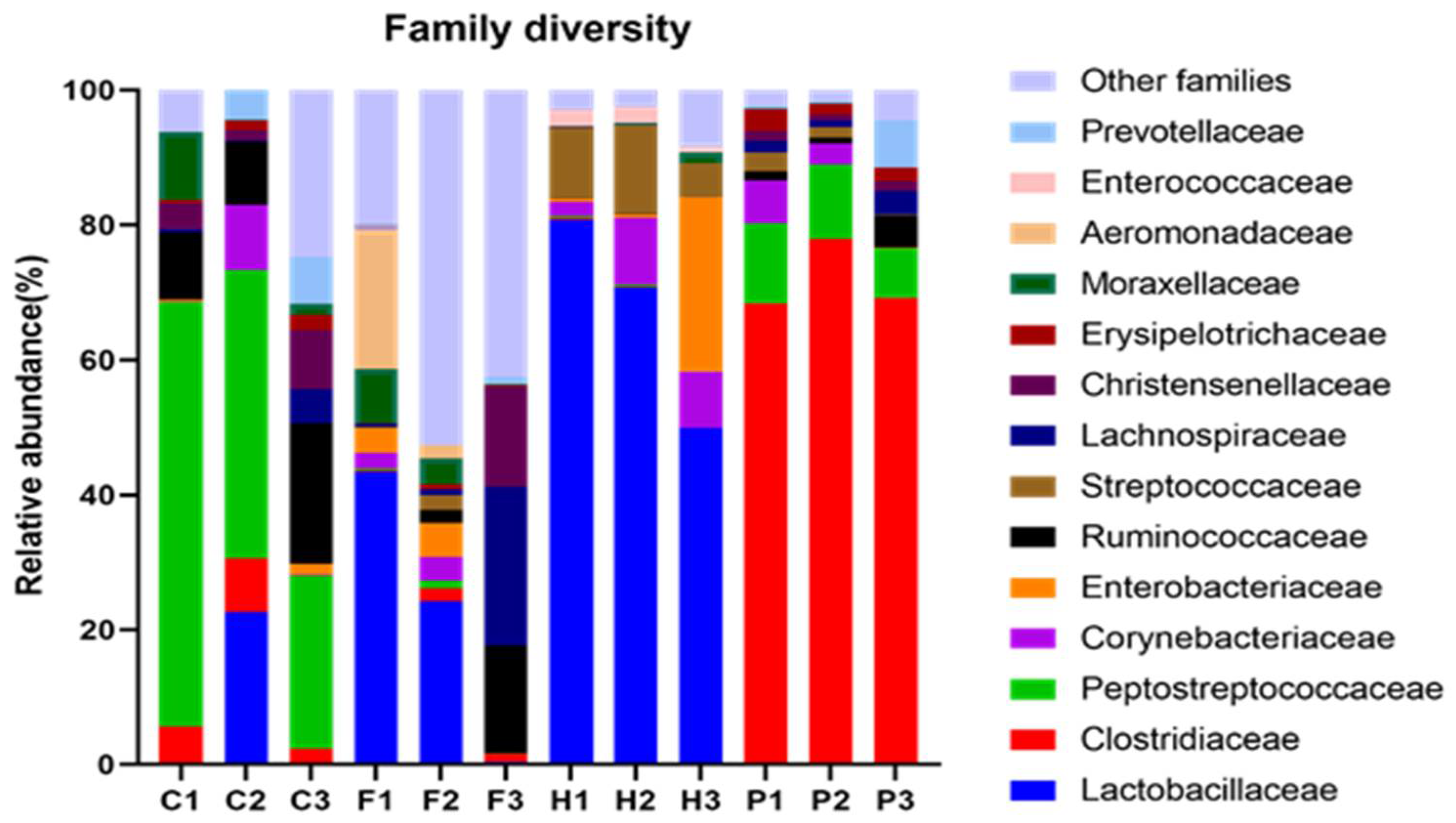
3.3. Phylum and Family Diversity
3.4. Genus Diversity in Manure Samples
4. Discussion
4.1. Taxonomical Similarities and Differences in Biological Replicates of Manures from the Same Animal Species
4.2. Feed Composition Influence on Faecal Microbiota of Cow, Chicken, Horse, and Pigs
4.3. Feed versus Manure Compositions, and Their Likely Influence on Biogas Production
5. Conclusions
6. Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Animal | Location | Sample ID | Feed Type | %C | %H | %N | %S | %O |
|---|---|---|---|---|---|---|---|---|
| Cow | Boerdery Farm | C1 | Cattle feed | 42.7 ± 0.2 | 6.40 ± 0.1 | 4.47 ± 0.03 | 0.145 ± 0.01 | 46.3 ± 0.1 |
| Chicken | Country Portion Farm | F2 | Chicken feed | 42.2 ± 0.2 | 6.12 ± 0.1 | 2.31 ± 0.05 | 0 | 49.4 ± 0.2 |
| Layers’ feed | 38.9 ± 0.1 | 5.74 ± 0.1 | 2.53 ± 0.05 | 0.222 ± 0.01 | 52.6 ± 0.2 | |||
| Pig | P3 | Bran and white maize meal | 38.3 ± 0.2 | 5.82 ± 0.1 | 1.72 ± 0.05 | 0.155 ± 0.01 | 54.0 ± 0.2 | |
| Crushed yellow maize | 39.9 ± 0.2 | 6.03 ± 0.2 | 1.42 ± 0.05 | 0 | 52.6 ± 0.2 | |||
| Horse | EARTH Centre | H1 | Equus Nice and Easy | 43.0 ± 0.1 | 6.34 ± 0.1 | 3.36 ± 0.03 | 0.167 ± 0.01 | 47.1 ± 0.1 |
| Eragrostis horse feed | 47.0 ± 0.3 | 6.06 ± 0.1 | 0.928 ± 0.01 | 0.171 ± 0.01 | 45.8 ± 0.1 | |||
| Active bacteria | 36.6 ± 0.2 | 5.21 ± 0.1 | 2.68 ± 0.03 | 2.25 ± 0.02 | 53.2 ± 0.2 | |||
| Equus all-time balancer | 39.1 ± 0.3 | 5.79 ± 0.2 | 3.20 ± 0.01 | 0.337 ± 0.01 | 51.6 ± 0.2 | |||
| Equus Train and Leisure | 40.1 ± 0.2 | 5.93 ± 0.1 | 2.33 ± 0.03 | 0.249 ± 0.01 | 51.4 ± 0.2 | |||
| Horse | Harveston Stables | H2 | Horse feed | 42.5 ± 0.2 | 6.08 ± 0.1 | 2.16 ± 0.02 | 0.220 ± 0.01 | 49.0 ± 0.2 |
| Cow | Bosheuvel Country Estates | C3 | Mixed vegetables | 30.7 ± 0.1 | 4.53 ± 0.1 | 3.53 ± 0.03 | 0.488 ± 0.01 | 60.7 ± 0.2 |
| Chicken | F3 | |||||||
| Pig | P1, P2 |
| Manure Type | Location | Sample ID | %C | %H | %N | %S | %O |
|---|---|---|---|---|---|---|---|
| Cow dung | Boerdery Farm | C1 | 28.4 ± 0.1 | 4.04 ± 0.03 | 5.72 ± 0.05 | 0.223 ± 0.01 | 61.6 ± 0.3 |
| Cow dung | Kates Farm | C2 | 38.5 ± 0.1 | 5.32 ± 0.03 | 3.74 ± 0.03 | 0.217 ± 0.01 | 52.2 ± 0.3 |
| Cow dung | Bosheuvel Country Estates | C3 | 20.0 ± 0.2 | 3.56 ± 0.02 | 7.17 ± 0.03 | 0.313 ± 0.01 | 69.0 ± 0.2 |
| Chicken manure | Kates Farm | F1 | 38.8 ± 0.1 | 5.35 ± 0.04 | 1.75 ± 0.02 | 0.219 ± 0.01 | 53.9 ± 0.3 |
| Chicken manure | Country Portion Farm | F2 | 23.7 ± 0.2 | 3.11 ± 0.03 | 9.80 ± 0.04 | 0.251 ± 0.01 | 63.1 ± 0.2 |
| Chicken manure | Bosheuvel Country Estates | F3 | 33.0 ± 0.1 | 4.82 ± 0.01 | 1.69 ± 0.03 | 0.205 ± 0.01 | 57.3 ± 0.2 |
| Pig manure | Bosheuvel Country Estates | P1 | 26.7 ± 0.1 | 4.26 ± 0.02 | 2.67 ± 0.02 | 0.292 ± 0.01 | 66.1 ± 0.2 |
| Pig manure | Bosheuvel Country Estates | P2 | 29.9 ± 0.2 | 4.96 ± 0.02 | 3.63 ± 0.02 | 0.234 ± 0.01 | 61.3 ± 0.3 |
| Pig manure | Country Portion Farm | P3 | 42.8 ± 0.2 | 5.52 ± 0.03 | 1.34 ± 0.02 | 0.271 ± 0.01 | 50.1 ± 0.2 |
| Horse manure | EARTH Centre | H1 | 38.0 ± 0.2 | 5.56 ± 0.02 | 2.17 ± 0.02 | 0.313 ± 0.01 | 54.0 ± 0.2 |
| Horse manure | Harveston Stables | H2 | 40.2 ± 0.1 | 5.25 ± 0.01 | 1.26 ± 0.02 | 0.176 ± 0.01 | 53.1 ± 0.2 |
| Horse manure | Barent Horse Stables | H3 | 27.7 ± 0.2 | 3.63 ± 0.02 | 1.85 ± 0.03 | 0.219 ± 0.01 | 66.6 ± 0.3 |
Appendix B
| Cow Dung | |||
|---|---|---|---|
| Genera Common in All Manure Samples | Uniquely Prevalent Genera Specific to Locations | ||
| C1, C2, C3 | Boerdery Farm Produce (C1) | Kates Farm (C2) | Bosheuvel Country Estates (C3) |
| Lachnospiracaeae_uc Ruminoccocaceae_uc Sporobacter Christensenellaceae_uc Treponema Oscillibacter Terrisporobacter Turicibacter Clostridium Romboutsia Acinetobacter Corynebacterium | Glutamicibacter Enterococcus Jeotgalicoccus | Prevotella Lactobacillaceae_uc | Cellulosilyticum Enterobacter Lysinibacillus Psychrobacter |
| Chicken droppings | |||
| F1, F2, F3 | Kates Farm (F1) | Country Portion Farm (F2) | Bosheuvel Country Estates (F3) |
| Clostridium Acinetobacter Pseudomonas Lactobacillus | Oceanisphaera Glutamicibacter Escherichia Enterococcus Jeotgalicoccus | Oscillibacter Streptococcus | Christensenellaceae_uc Sporobacter |
| Horse manure | |||
| H1, H2, H3 | Earth Centre (H1) | Harveston Stables (H2) | Barent Horse Stables (H3) |
| Romboutsia Acinetobacter Pseudomonas Lactobacillus Corynebacterium Aerococcus Lactobacillaceae_uc Escherichia Enterococcus | Prevotella Christensenellaceae_uc Treponema | Lachnospiracaeae_uc Sporobacter Alistipes Oceanisphaera | Enterobacter Weissella Kosakonia |
| Pig manure | |||
| P1, P2, P3 | Bosheuvel Country Estates (P1) | Bosheuvel Country Estates (P2) | Country Portion Farm (P3) |
| Prevotella Cellulosilyticum Peptostreptococaceae_uc Clostridiaceae_uc Lachnospiracaeae_uc Ruminoccocaceae_uc Sporobacter Christensenellaceae_uc Treponema Ruminococcus Oscillibacter Terrisporobacter Turicibacter Clostridium Romboutsia Pseudomonas Streptococcus Lactobacillus Corynebacterium | Acinetobacter Lactobacillaceae_uc Enterococcus | --- | Enterobacter Escherichia |
| Cow Dung | Chicken Droppings | Horse Manure | Pig Manure |
|---|---|---|---|
| Lachnospiracaeae_uc | Clostridium | Romboutsia | Prevotella |
| Ruminoccocaceae_uc | Lactobacillus | Lactobacillus | Cellulosilyticum |
| Sporobacter | Escherichia | Aerococcus | Peptostreptococaceae_uc |
| Christensenellaceae_uc | Enterococcus | Lactobacillaceae_uc | Clostridiaceae_uc |
| Treponema | Jeotgalicoccus | Escherichia | Lachnospiracaeae_uc |
| Oscillibacter | Oscillibacter | Enterococcus | Ruminoccocaceae_uc |
| Terrisporobacter | Streptococcus | Prevotella | Sporobacter |
| Turicibacter | Christensenellaceae_uc | Christensenellaceae_uc | Christensenellaceae_uc |
| Clostridium | Sporobacter | Treponema | Treponema |
| Romboutsia | Lachnospiracaeae_uc | Ruminococcus | |
| Enterococcus | Sporobacter | Oscillibacter | |
| Jeotgalicoccus | Alistipes | Terrisporobacter | |
| Prevotella | Weissella | Turicibacter | |
| Lactobacillaceae_uc | Clostridium | ||
| Cellulosilyticum | Romboutsia | ||
| Streptococcus | |||
| Lactobacillus | |||
| Lactobacillaceae_uc | |||
| Enterococcus | |||
| Escherichia |
References
- Madigan, M.T.; Martinko, J.M.; Dunlap, P.V.; Clark, D.P. Brock Biology of microorganisms 12th edn. Int. Microbiol. 2008, 11, 65–73. [Google Scholar]
- Ali Shah, F.; Mahmood, Q.; Maroof Shah, M.; Pervez, A.; Ahmad Asad, S. Microbial ecology of anaerobic digesters: The key players of anaerobiosis. Sci. World J. 2014, 183752. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I. Production of biogas: Relationship between methanogenic and sulfate-reducing microorganisms. Open Life Sci. 2017, 12, 82–91. [Google Scholar] [CrossRef]
- Mutungwazi, A.; Ijoma, G.N.; Matambo, T.S. The significance of microbial community functions and symbiosis in enhancing methane production during anaerobic digestion: A review. Symbiosis 2020, 1, 1–24. [Google Scholar] [CrossRef]
- Ijoma, G.N.; Nkuna, R.; Mutungwazi, A.; Rashama, C.; Matambo, T.S. Applying PICRUSt and 16S rRNA functional characterisation to predicting co-digestion strategies of various animal manures for biogas production. Sci. Rep. 2021, 11, 19913. [Google Scholar] [CrossRef]
- Shin, J.; Cho, S.K.; Lee, J.; Hwang, K.; Chung, J.W.; Jang, H.N.; Shin, S.G. Performance and microbial community dynamics in anaerobic digestion of waste activated sludge: Impact of immigration. Energies 2019, 12, 573. [Google Scholar] [CrossRef] [Green Version]
- Ohemeng-Ntiamoah, J.; Datta, T. Perspectives on variabilities in biomethane potential test parameters and outcomes: A review of studies published between 2007 and 2018. Sci. Total Environ. 2019, 664, 1052–1062. [Google Scholar] [CrossRef]
- Majd, S.; Abdoli, M.A.; Karbassi, A.; Pourzamani, H.R.; Rezaee, M. Effect of Physical and Chemical Operating Parameters on Anaerobic Digestion of Manure and Biogas Production: A Review. J. Environ. Health Sustain. Dev. 2017, 2, 235–247. [Google Scholar]
- Sibiya, N.T.; Muzenda, E.; Mbohwa, C. Evaluation of potential substrates for biogas production via anaerobic digestion: A review. Lect. Notes Eng. Comput. Sci. 2017, 2, 583–588. [Google Scholar]
- He, Q.; Li, L.; Zhao, X.; Qu, L.; Wu, D.; Peng, X. Investigation of foaming causes in three mesophilic food waste digesters: Reactor performance and microbial analysis. Sci. Rep. 2017, 7, 13701. [Google Scholar] [CrossRef] [Green Version]
- Illumina Targeted Next-Generation Sequencing vs qPCR and Sanger Sequencing. Available online: https://emea.illumina.com/content/dam/illumina-marketing/documents/products/other/infographic-targeted-ngs-vs-sanger-qpcr.pdf (accessed on 17 November 2019).
- Verberkmoes, N.C.; Russell, A.L.; Shah, M.; Godzik, A.; Rosenquist, M.; Halfvarson, J.; Lefsrud, M.G.; Apajalahti, J.; Tysk, C.; Hettich, R.L.; et al. Shotgun metaproteomics of the human distal gut microbiota. ISME J. 2009, 3, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukumba, P.; Makaka, G. Biogas Production From a Field Batch Biogas Digester Using Cow Dung As a substrate. Int. J. Eng. Res. Sci. Technol. 2015, 4, 1–10. [Google Scholar]
- Agarry, S.E. Comparison of Biogas production from Cow dung and Pig dung under Mesophilic condition. Int. Ref. J. Eng. Sci. 2012, 1, 16–21. [Google Scholar]
- Prasad, R.D. Empirical Study on Factors Affecting Biogas Production. ISRN Renew. Energy 2012, 2012, 136959. [Google Scholar] [CrossRef] [Green Version]
- Prapinagsorn, W.; Sittijunda, S.; Reungsang, A. Co-digestion of napier grass and its silage with cow dung for methane production. Energies 2017, 10, 1654. [Google Scholar] [CrossRef] [Green Version]
- Priadi, C.; Wulandari, D.; Rahmatika, I.; Moersidik, S.S. Biogas Production in the Anaerobic Digestion of Paper Sludge. APCBEE Procedia 2014, 9, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Olowoyeye, J. Comparative studies on biogas production using six different animal dungs. J. Energy Technol. Policy 2013, 3, 1–6. [Google Scholar]
- Mönch-Tegeder, M.; Lemmer, A.; Oechsner, H. Enhancement of methane production with horse manure supplement and pretreatment in a full-scale biogas process. Energy 2014, 73, 523–530. [Google Scholar] [CrossRef]
- Mönch-Tegeder, M.; Lemmer, A.; Oechsner, H.; Jungbluth, T. Investigation of the methane potential of horse manure. Agric. Eng. Int. CIGR J. 2013, 15, 161–172. [Google Scholar]
- Wang, X.; Lu, X.; Li, F.; Yang, G. Effects of Temperature and Carbon-Nitrogen (C/N) Ratio on the Performance of Anaerobic Co-Digestion of Dairy Manure, Chicken Manure and Rice Straw: Focusing on Ammonia Inhibition. PLoS ONE 2014, 9, e97265. [Google Scholar] [CrossRef] [Green Version]
- Scheftelowitz, M.; Thrän, D. Unlocking the Energy Potential of Manure—An Assessment of the Biogas Production Potential at the Farm Level in Germany. Agriculture 2016, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Suresh, A.; Choi, H.L.; Kannan, N.; Rajagopal, K. Biochemical Methane Potentials and Organic Matter Degradation of Swine Slurry under Mesophilic Anaerobic Digestion. Indian J. Sci. Technol. 2016, 9, 1–6. [Google Scholar] [CrossRef]
- Bahrndorff, S.; Alemu, T.; Alemneh, T.; Nielsen, J.L. The Microbiome of Animals: Implications for Conservation Biology. Int. J. Genom. 2016, 2016, 5304028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spergser, J.; Loncaric, I.; Tichy, A.; Fritz, J.; Scope, A. The cultivable autochthonous microbiota of the critically endangered Northern bald ibis (Geronticus eremita). PLoS ONE 2018, 13, e0195255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Lozupone, C.A.; Hamady, M.; Knight, R.; Jeffrey, I.G. Worlds within worlds: Evolution of the vertebrate gut microbiota. Natl. Inst. Health 2008, 6, 776–788. [Google Scholar] [CrossRef] [Green Version]
- Zoetendal, E.G.; Collier, C.T.; Koike, S.; Mackie, R.I.; Gaskins, H.R. Diet Induced Changes in the Colonic Environment and Colorectal Cancer Molecular Ecological Analysis of the Gastrointestinal Microbiota: A Review 1. J. Nutr. 2004, 134, 479–482. [Google Scholar] [CrossRef]
- Barbosa, A.; Balagué, V.; Valera, F.; Martínez, A.; Benzal, J. Age-Related Differences in the Gastrointestinal Microbiota of Chinstrap Penguins (Pygoscelis antarctica). PLoS ONE 2016, 11, e0153215. [Google Scholar] [CrossRef] [Green Version]
- Krotz, L.; Giazzi, G. Thermo Scientific FLASH 2000 CHNS Analyzer: Stability, Linearity, Repeatability and Accuracy. Thermo Fish. Sci. Inc. 2014, 2–3. Available online: https://www.brechbuehler.ch/fileadmin/redacteur/pdf/oea/FLASHSmart/app-notes/AN42213_Flash_2000_CHNS_Analyzer_Stability-LInearity-Repeatability_and_Accuracy.pdf (accessed on 3 March 2021).
- Selvarajan, R.; Sibanda, T.; Venkatachalam, S.; Ogola, H.J.O.; Christopher Obieze, C.; Msagati, T.A. Distribution, Interaction and Functional Profiles of Epiphytic Bacterial Communities from the Rocky Intertidal Seaweeds, South Africa. Sci. Rep. 2019, 9, 19835. [Google Scholar] [CrossRef]
- Nkuna, R.; Roopnarain, A.; Adeleke, R. Effects of organic loading rates on microbial communities and biogas production from water hyacinth: A case of mono- and co-digestion. J. Chem. Technol. Biotechnol. 2019, 94, 1294–1304. [Google Scholar] [CrossRef]
- Ni, J.; Li, X.; He, Z.; Xu, M. A novel method to determine the minimum number of sequences required for reliable microbial community analysis. J. Microbiol. Methods. 2017, 4, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the Environment. Microbiol. Mol. Biol. Rev. 2012, 76, 685–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, W.W.; Wang, T.T.; Bai, J.L.; Ding, P.; Xing, K.; Jiang, J.H.; Peng, X.; Qin, S. Glutamicibacter halophytocola sp. nov., an endophytic actinomycete isolated from the roots of a coastal halophyte, Limonium sinense. Int. J. Syst. Evol. Microbiol. 2017, 67, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Feiner, G. The microbiology of specific bacteria. In Technology and Nutrition, Meat Products Handbook; Feiner, G., Ed.; Woodhead Publishing: Sawston, UK, 2006; Volume 53, pp. 595–615. ISBN 9781845690502. [Google Scholar]
- Flint, H.J.; Duncan, S.H. Bacteroides and Prevotella. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 203–208. [Google Scholar] [CrossRef]
- Iljazovic, A.; Roy, U.; Gálvez, E.J.C.; Lesker, T.R.; Zhao, B.; Gronow, A.; Amend, L.; Will, S.E.; Hofmann, J.D.; Pils, M.C.; et al. Perturbation of the gut microbiome by Prevotella spp. enhances host susceptibility to mucosal inflammation. Mucosal Immunol. 2020, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Margolis, E.; Fredricks, D.N. Chapter 83-Bacterial Vaginosis-Associated Bacteria. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Ian Poxton, J.S., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 1487–1496. ISBN 9780123971692. [Google Scholar]
- Cai, S.; Dong, X. Cellulosilyticum ruminicola gen. nov., sp. nov., isolated from the rumen of yak, and reclassification of Clostridium lentocellum as Cellulosilyticum lentocellum comb. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 845–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, K. Enterobacter. Encyclopedia Britannica. 2020. Available online: https://www.britannica.com/science/Enterobacter (accessed on 7 March 2022).
- Cai, S.; Li, J.; Ze Hu, F.; Zhang, K.; Luo, Y.; Janto, B.; Boissy, R.; Ehrlich, G.; Dong, X. Cellulosilyticum ruminicola, a newly described rumen bacterium that possesses redundant Fibrolytic-protein-encoding genes and degrades lignocellulose with multiple carbohydrateborne fibrolytic enzymes. Appl. Environ. Microbiol. 2010, 76, 3818–3824. [Google Scholar] [CrossRef] [Green Version]
- Difford, G.F.; Plichta, D.R.; Løvendahl, P.; Lassen, J.; Noel, S.J.; Højberg, O.; Wright, A.D.G.; Zhu, Z.; Kristensen, L.; Nielsen, H.B.; et al. Host genetics and the rumen microbiome jointly associate with methane emissions in dairy cows. PLoS Genet. 2018, 14, e1007580. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes. 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Hammond, J.N.C. Cow Manure as a Source of Certain Vitamins for Growing Chickens. Poult. Sci. 1942, 21, 554–559. [Google Scholar] [CrossRef]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef]
- Bomba, A.; Jonecová, Z.; Gancarčíková, S.; Nemcová, R. The gastrointestinal microbiota of farm animals. Gastrointest. Microbiol. 2006, 381–400. [Google Scholar] [CrossRef]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Morton, J.; Jackson, B.; Freeman, M.; Eckard, R.; Nettle, R.; Doonan, B.; Hough, G. Nutrition Management for Tasmanian Dairy Farmers Acknowledgments 2007. Available online: https://www.utas.edu.au/__data/assets/pdf_file/0014/1113404/Nutrition-Management-for-Tasmanian-Dairy-Farmers.pdf (accessed on 3 March 2021).
- Miller, E.L. Protein nutrition requirements of farmed livestock and dietary supply. In Proceedings of the Protein Sources for the Animal Feed Industry. FAO Expert Consultation and Workshop, Bangkok, Thailand, 29 April–3 May 2002; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2004; pp. 29–75. [Google Scholar]
- Bambi, G.; Rossi, G.; Barbari, M. Comparison between different types of bedding materials for horses. Agron. Res. 2018, 16, 646–655. [Google Scholar] [CrossRef]
- Saastamoinen, M.; Särkijärvi, S.; Hyyppä, S. Reducing respiratory health risks to horses and workers: A comparison of two stall bedding materials. Animals 2015, 5, 965–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, J.R. Minerals for Livestock; The Oregon Agricultural Experiment Station (OAES): Corvallis, OR, USA, 1951. [Google Scholar]
- Department of Agriculture Intermediate Animal Nutrition 2006. pp. 1–79. Available online: https://www.gov.za/sites/default/files/gcis_document/201409/28711b.pdf (accessed on 7 November 2021).
- Thornton, P.K. Livestock production: Recent trends, future prospects. Phil. Trans. R. Soc. B 2010, 365, 2853–2867. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Soni, A. Elemental Ratio and Their Importance in Feed and Fodder. Int. J. Pure App. Biosci. 2014, 2, 154–160. [Google Scholar]
- Perry, T.W. Nutrient Requirements of Beef Cattle. In Beef Cattle Feeding and Nutrition; National Academy Press: Washington, DC, USA, 1995; pp. 351–369. [Google Scholar] [CrossRef]
- Chase, L.E. Understanding Nitrogen Utilization in Dairy Cattle; Cornell University: Ithaca, NY, USA, 2007; Available online: https://s3.wp.wsu.edu/uploads/sites/346/2014/11/UnderstandingNUtilization.pdf (accessed on 3 March 2019).
- Erickson, P.S.; Kalscheur, K.F. Nutrition and feeding of dairy cattle. In Animal Agriculture; Academic Press: Cambridge, MA, USA, 2020; pp. 157–180. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Minamoto, Y. Gastrointestinal microorganisms in cats and dogs: A brief review. Arch. Med. Vet. 2013, 45, 111–124. [Google Scholar] [CrossRef]
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the gut microbiome using 16S or shotgun metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, G.; Osman, S.; Kukkadapu, R.; Engelhard, M.; Vaishampayan, P.A.; Andersen, G.L.; Sani, R.K. Microbial and Mineralogical Characterizations of Soils Collected from the Deep Biosphere of the Former Homestake Gold Mine, South Dakota. Microb. Ecol. 2010, 60, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Selvarajan, R.; Sibanda, T.; Tekere, M. Thermophilic bacterial communities inhabiting the microbial mats of “indifferent” and chalybeate (iron-rich) thermal springs: Diversity and biotechnological analysis. Microbiologyopen 2018, 7, e00560. [Google Scholar] [CrossRef]
- Anandan, R.; Dharumadurai, D.; Manogaran, G.P. An Introduction to Actinobacteria. In Actinobacteria-Basics and Biotechnological Applications; IntechOpen: London, UK, 2016; pp. 1–37. [Google Scholar]
- Shivlata, L.; Tulasi, S. Thermophilic and alkaliphilic Actinobacteria: Biology and potential applications. Front. Microbiol. 2015, 6, 1014. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezeld, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Am. Soc. Microbiol. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut Bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.L.; Heaver, S.L.; Walters, W.A.; Ley, R.E. Microbiome and metabolic disease: Revisiting the bacterial phylum Bacteroidetes. J. Mol. Med. 2017, 95, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Mahmood, S.; Adeolu, M. A phylogenomic and molecular signature based approach for characterization of the phylum spirochaetes and its major clades: Proposal for a taxonomic revision of the phylum. Front. Microbiol. 2013, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- VanHorn, H. Factors affecting manure quantity, quality, and use. In Proceedings of the Mid-South Ruminant Nutrition Conference, Dallas, TX, USA, 7–8 May 1998; pp. 9–20. [Google Scholar]
- Manitoba. Agriculture Properties of Manure. Manitoba Agric. Food Rural Dev. 2015, 42. Available online: https://www.gov.mb.ca/agriculture/environment/nutrient-management/pubs/properties-of-manure.pdf (accessed on 3 March 2020).
- Barnes, C.; Sweeting, C.J.; Jennings, S.; Barry, J.T.; Polunin, N.V.C. Effect of temperature and ration size on carbon and nitrogen stable isotope trophic fractionation. Functional Ecology 2007, 21, 356–362. [Google Scholar] [CrossRef]
- Macias-Corral, M.A.; Cueto-Wong, J.A.; Morán-Martínez, J.; Reynoso-Cuevas, L. Effect of different initial C/N ratio of cow manure and straw on microbial quality of compost. Int. J. Recycl. Org. Waste Agric. 2019, 8, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Ogunwande, G.A.; Osunade, J.A.; Adeagbo, A.O.; Fakuyi, O.F. Effects of co-digesting swine manure with chicken Manure on biogas production. IFE J. Sci. 2013, 15, 1–8. [Google Scholar]
- Pham, C.H.; Vu, C.C.; Sommer, S.G.; Bruun, S. Factors affecting process temperature and biogas production in small-scale rural biogas digesters in winter in northern vietnam. Asian-Australas. J. Anim. Sci. 2014, 27, 1050–1056. [Google Scholar] [CrossRef] [Green Version]
- Mulka, R.; Szulczewski, W.; Szlachta, J.; Prask, H. The influence of carbon content in the mixture of substrates on methane production. Clean Technol. Environ. Policy 2016, 18, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Barnett, A.; International Development Research Centre; Pyle, L.; Subramanian, S.K. Biogas Technology in the Third World; Management Institute: New Delhi, India, 1978; ISBN 0-88936-162-2. [Google Scholar]
- Maramba, F. Biogas and Waste Recycling; Liberty Flour Mills Inc.: Makati, Philippines, 2019; Volume 22, ISBN 9788578110796. [Google Scholar]
- Khan, A.W.; Trottier, T.M. Effect of sulfur-containing compounds on anaerobic degradation of cellulose to methane by mixed cultures obtained from sewage sludge. Appl. Environ. Microbiol. 1978, 35, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegarty, R.S.; Gerdes, R. Hydrogen production and transfer in the rumen. Recent Adv. Anim. Nutr. Aust. 1999, 12, 37–44. [Google Scholar]
- Salam, B.; Biswas, S.; Rabbi, M.S. Biogas from mesophilic anaerobic digestion of cow dung using silica gel as catalyst. Procedia Eng. 2015, 105, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Ulusoy, Y.; Ulukardesler, A.H.; Arslan, R.; Tekin, Y. Energy and emission benefits of chicken manure biogas production: A case study. In Proceedings of the 2017 IEEE 6th International Conference on Renewable Energy Research and Applications (ICRERA), San Diego, CA, USA, 5–8 November 2017; pp. 648–652. [Google Scholar] [CrossRef]
- Okewale, A.O.; Adesina, O.A. Evaluation of biogas production from co-digestion of pig dung, water hyacinth and poultry droppings. Waste Dispos. Sustain. Energy 2019, 1, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Agayev, E.; Ugurlu, A. Biogas production from co-digestion of horse manure and waste sewage sludge. TechConnect Briefs 2011, 3, 657–660. [Google Scholar]
- Bermingham, E.N.; Maclean, P.; Thomas, D.G.; Cave, N.J.; Young, W. Key bacterial families (Clostridiaceae, Erysipelotrichaceae and Bacteroidaceae) are related to the digestion of protein and energy in dogs. PeerJ 2017, 5, e3019. [Google Scholar] [CrossRef]
- Schingoethe, D.J. Whey Utilization in Animal Feeding: A Summary and Evaluation. J. Dairy Sci. 1976, 59, 556–570. [Google Scholar] [CrossRef]
- Campbell, B.; Kreider, R.B.; Ziegenfuss, T.; La Bounty, P.; Roberts, M.; Burke, D.; Landis, J.; Lopez, H.; Antonio, J. International Society of Sports Nutrition position stand: Protein and exercise. J. Int. Soc. Sports Nutr. 2007, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Mastellar, S.L.; Moffet, A.; Harris, P.A.; Urschel, K.L. Effects of threonine supplementation on whole-body protein synthesis and plasma metabolites in growing and mature horses. Vet. J. 2016, 207, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Mok, C.H.; Urschel, K.L. Amino acid requirements in horses. Asian-Australas. J. Anim. Sci. 2020, 33, 679–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, A.; Iyer, A.B.; Arif, S.; Junaid, M.; Khan, R.S.; Nazir, W.; Khalid, N. Whey Protein-Based Functional Energy Drinks Formulation and Characterization. In Sports and Energy Drinks; Alexandru Mihai Grumezescu, A.M.H., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 161–181. ISBN 9780128158517. [Google Scholar]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by lachnospiraceae and ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Stanton, C.; Lane, J.A.; Smith, G.J.; Ross, R.P. Precision nutrition and the microbiome, part I: Current state of the science. Nutrients 2019, 11, 923. [Google Scholar] [CrossRef] [Green Version]
- Bascomb, S.; Manafi, M. Use of enzyme tests in characterization and identification of aerobic and facultatively anaerobic gram-positive cocci. Clin. Microbiol. Rev. 1998, 11, 318–340. [Google Scholar] [CrossRef] [Green Version]
- Pompilio, A.; Di Bonaventura, G.; Gherardi, G. An overview on streptococcus bovis/streptococcus equinus complex isolates: Identification to the species/subspecies level and antibiotic resistance. Int. J. Mol. Sci. 2019, 20, 480. [Google Scholar] [CrossRef] [Green Version]
- Dunican, L.K.; Seeley, H.W. Starch hydrolysis by Streptococcus equinus. J. Bacteriol. 1962, 83, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Spinler, J. Encyclopedia of Metagenomics: Environmental Metagenomics; Springer: Berlin, Germany, 2015. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yang, X. Biofuel production from food wastes. In Handbook of Biofuels Production, 2nd ed.; Luque, R., Sze Ki Lin, C., Karen Wilson, J.C., Eds.; Woodhead Publishing: Sawston, UK, 2016; Volume 53, pp. 617–653. ISBN 9780081004555. [Google Scholar]
- Jang, H.B.; Choi, M.K.; Kang, J.H.; Park, S.I.; Lee, H.J. Association of dietary patterns with the fecal microbiota in Korean adolescents. BMC Nutr. 2017, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Murray, P. Manual of Clinical Microbiology, 6th ed.; American Society for Microbiology: Washington, DC, USA, 1995. [Google Scholar]
- Fusco, V.; Franz, C.M.A.P.; Safety, F. Opportunistic Food-Borne Pathogens. In Food Safety and Preservation; Mihai Grumezescu, A., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 269–306. ISBN 9780128149560. [Google Scholar]
- Becker, M.; Patz, S.; Becker, Y.; Berger, B.; Drungowski, M.; Bunk, B.; Overmann, J.; Spröer, C.; Reetz, J.; Tchuisseu Tchakounte, G.V.; et al. Comparative Genomics Reveal a Flagellar System, a Type VI Secretion System and Plant Growth-Promoting Gene Clusters Unique to the Endophytic Bacterium Kosakonia radicincitans. Front. Microbiol. 2018, 9, 1997. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Ma, W.; Jin, M.; Chen, J.; Li, S.; Zou, X. Complete Genome Sequence of Kosakonia sp. Strain CCTCC M2018092, a Fucose-Rich Exopolysaccharide Producer. Microbiol. Resour. Announc. Am. Soc. Microbiol. 2019, 8, e00567-19. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.C.; Frick, I. Gram-positive anaerobic cocci–commensals and opportunistic pathogens. FEMS Microbiol. Rev. 2013, 37, 520–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, R.; Fouhse, J.M.; Tiwari, U.P.; Li, L.; Willing, B.P. Dietary fiber and intestinal health of monogastric animals. Front. Vet. Sci. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Sample | Sample ID | Sources | Location | Description of Farm (Location) Where Samples Were Collected from |
|---|---|---|---|---|
| Cow dung | C1 | Boerdery Farm Produce | −26.21779, 27.6414784 | An animal feeds producing and cattle rearing farm in Randfontein, Gauteng, South Africa, that feeds livestock on grains and concentrates |
| C2 | Kates Farm | −26.0990952, 27.9103166 | A bed and breakfast facility in the periphery of Johannesburg, Gauteng serving free-range chicken and beef | |
| C3 | Bosheuvel Country Estates | −26.0249182, 27.8929324 | A multi-purpose farm in the Muldersdrift, Johannesburg with an on-site vintage hotel, restaurant, event venues, and conference rooms where Pinzgauer cattle, chickens, sheep, and pigs fed mostly on mixed vegetables and tubers are reared and served to guests | |
| Chicken droppings | F1 | Kates Farm | −26.0990952, 27.9103166 | A bed and breakfast facility in the periphery of Johannesburg, Gauteng serving free-range chicken and beef |
| F2 | Country Portion Farm | −26.224517, 27.6325603 | A farm in Randfontein that produces vegetables, poultry, and pork products to supply local restaurants and consumers. The poultry and pigs are fed on corn and sorghum and nutrient concentrates | |
| F3 | Bosheuvel Country Estates | −26.0249182, 27.8929324 | A multi-purpose farm in the Muldersdrift, Johannesburg with an on-site vintage hotel, restaurant, event venues, and conference rooms where Pinzgauer cattle, chickens, sheep, and pigs fed mostly on mixed vegetables and tubers are reared and served to guests | |
| Horse manure | H1 | Earth Centre | −26.080926, 27.8747353 | A non-profit making organisation in Ruimsig, Gauteng that caters to disabilities such as Autism, ADD / ADHD, Cerebral Palsy, Downs Syndrome, and the Deaf using locally reared horses in Equine Assisted Programmes. The horses are fed with a wide range of feed concentrates and probiotics. They also feed on hay where they sleep. |
| H2 | Harveston Stables | −26.0990952, 27.9103167 | A horse riding facility in Honeydew, Gauteng that offers lessons, pony rides, stabling, kids’ pony parties, and picnics. The horses are fed mainly on feed concentrates and they gave grass as their bedding. | |
| H3 | Barent Horse Stables | −26.21779, 27.6414783 | A plot rearing horses for family leisure in Randfontein, Gauteng. The horses are fed mostly on lucerne | |
| Pig manure | P1 | Bosheuvel Country Estates | −26.0249182, 27.8929324 | A multi-purpose farm in the Muldersdrift, Johannesburg with an on-site vintage hotel, restaurant, event venues, and conference rooms where Pinzgauer cattle, chickens, sheep, and pigs fed mostly on mixed vegetables and tubers are reared and served to guests |
| P2 | Bosheuvel Country Estates | −26.0249182, 27.8929324 | ||
| P3 | Country Portion Farm | −26.224517, 27.6325603 | A farm in Randfontein that produces vegetables, poultry, and pork products to supply local restaurants and consumers. The poultry and pigs are fed on corn and sorghum and nutrient concentrates |
| Sample ID | C-Utilisation Rate (%) | H-Utilisation Rate (%) | N-Utilisation Rate (%) | S-Utilisation Rate (%) |
|---|---|---|---|---|
| C1 | 33.5 ± 0.3 | 36.9 ± 0.13 | 7.96 ± 0.08 | 38.9 ± 0.02 |
| C3 | 35.1 ± 0.3 | 31.3 ± 0.12 | 6.20 ± 0.06 | 35.8 ± 0.02 |
| F2 | 4.52 ± 0.35 | 47.6 ± 0.13 | 5.63 ± 0.09 | 13.2 ± 0.02 |
| F3 | 7.43 ± 0.2 | 46.4 ± 0.11 | 5.06 ± 0.06 | 18.0 ± 0.02 |
| P1 | 13.3 ± 0.2 | 5.86 ± 0.12 | 1.31 ± 0.05 | 40.1 ± 0.02 |
| P2 | 12.8 ± 0.3 | 9.53 ± 0.12 | 0.928 ± 0.05 | 52.1 ± 0.02 |
| P3 | 9.36 ± 0.4 | 6.92 ± 0.18 | 2.44 ± 0.07 | 47.4 ± 0.01 |
| H1 | 17.8 ± 0.4 | 15.2 ± 0.12 | 43.1 ± 0.04 | 21.7 ± 0.02 |
| H2 | 15.4 ± 0.3 | 13.6 ± 0.11 | 41.4 ± 0.04 | 19.9 ± 0.02 |
| H3 | 15.0 ± 0.3 | 13.9 ± 0.12 | 41.2 ± 0.05 | 21.0 ± 0.02 |
| Variable | C1 | C2 | C3 | F1 | F2 | F3 |
| Valid reads | 12,310 | 10,110 | 9469 | 12,263 | 19,320 | |
| OTUs | 1316 | 408 | 1475 | 352 | 103 | 1255 |
| Simpson_1-D | 0.94 | 0.89 | 0.94 | 0.95 | 0.88 | |
| Shannon_H | 4.90 | 3.17 | 6.07 | 3.72 | 3.94 | 4.08 |
| Chao_1 | 1331.05 | 419.13 | 1529.51 | 432.29 | 188.55 | 1274.85 |
| ACE | 1377.64 | 442.42 | 1609.21 | 458.76 | 198.70 | 1330.69 |
| P1 | P2 | P3 | H1 | H2 | H3 | |
| Valid reads | 55,482 | 30,943 | 17,585 | 5152 | 71,805 | 17,500 |
| OTUs | 1628 | 941 | 234 | 971 | 333 | 339 |
| Simpson_1-D | 0.88 | 0.89 | 0.30 | 0.30 | 0.88 | |
| Shannon_H | 3.62 | 3.42 | 1.07 | 5.57 | 1.00 | 2.80 |
| Chao_1 | 1643.50 | 951.90 | 297.87 | 979.26 | 365.88 | 344.01 |
| ACE | 1689.24 | 983.54 | 318.43 | 1010.30 | 390.47 | 360.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutungwazi, A.; Ijoma, G.N.; Ogola, H.J.O.; Matambo, T.S. Physico-Chemical and Metagenomic Profile Analyses of Animal Manures Routinely Used as Inocula in Anaerobic Digestion for Biogas Production. Microorganisms 2022, 10, 671. https://doi.org/10.3390/microorganisms10040671
Mutungwazi A, Ijoma GN, Ogola HJO, Matambo TS. Physico-Chemical and Metagenomic Profile Analyses of Animal Manures Routinely Used as Inocula in Anaerobic Digestion for Biogas Production. Microorganisms. 2022; 10(4):671. https://doi.org/10.3390/microorganisms10040671
Chicago/Turabian StyleMutungwazi, Asheal, Grace N. Ijoma, Henry J. O. Ogola, and Tonderayi S. Matambo. 2022. "Physico-Chemical and Metagenomic Profile Analyses of Animal Manures Routinely Used as Inocula in Anaerobic Digestion for Biogas Production" Microorganisms 10, no. 4: 671. https://doi.org/10.3390/microorganisms10040671