
Antimicrobial Activity of Ohelo Berry (Vaccinium calycinum) Juice against Listeria monocytogenes and Its Potential for Milk Preservation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ohelo Berry Juice Preparation
2.2. Chemical Analysis of Ohelo Berry Juice
2.2.1. Quantification of Sugar and Organic Acids in Ohelo Berry Juice
2.2.2. Total Phenolic Concentration
2.2.3. Anthocyanin Concentration
2.3. Bacterial Strain and Growth Conditions
2.4. Determination of the Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentrations (MBC) of Ohelo Berry Juice
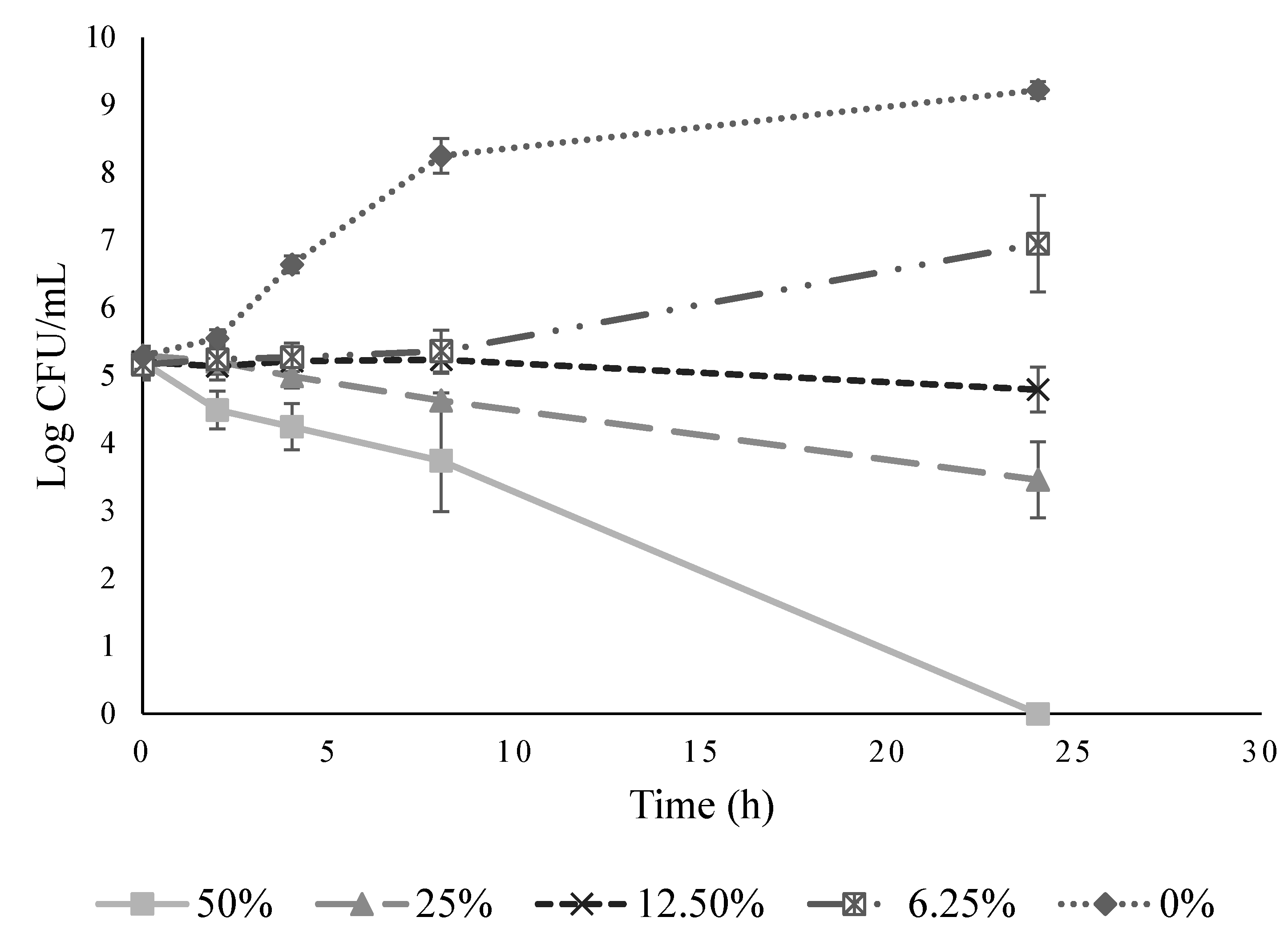
2.5. Growth Inhibition Kinetics of Ohelo Berry Juice against L. monocytogenes
2.6. Evaluation of the Physicochemical Properties and Biofilm Formation Capability of L. monocytogenes Treated with Ohelo Berry Juice
2.6.1. Hydrophobicity
2.6.2. Auto-Aggregation
2.6.3. Motility
2.6.4. Biofilm Formation
2.7. RT-qPCR
2.8. Effect of Ohelo Berry Juice on the Growth of L. monocytogenes in Milk
2.9. Statistical Analysis
3. Results
3.1. Chemical Analysis of Ohelo Berry Juice
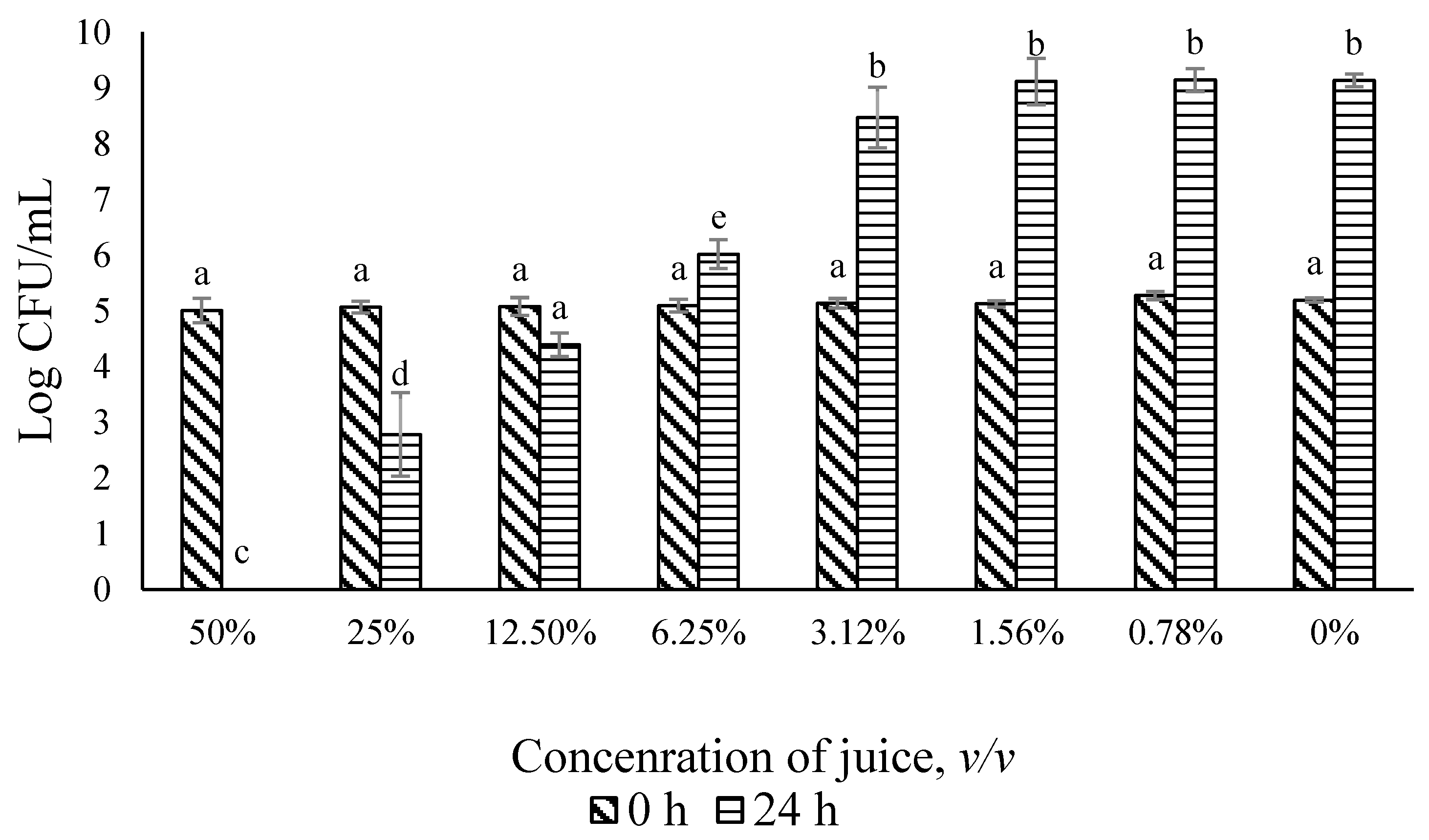
3.2. Antimicrobial Effect of Ohelo Berry Juice on L. monocytogenes
3.3. Effect of Ohelo Berry Juice on the Physicochemical Properties and Biofilm Formation Capability of L. monocytogenes
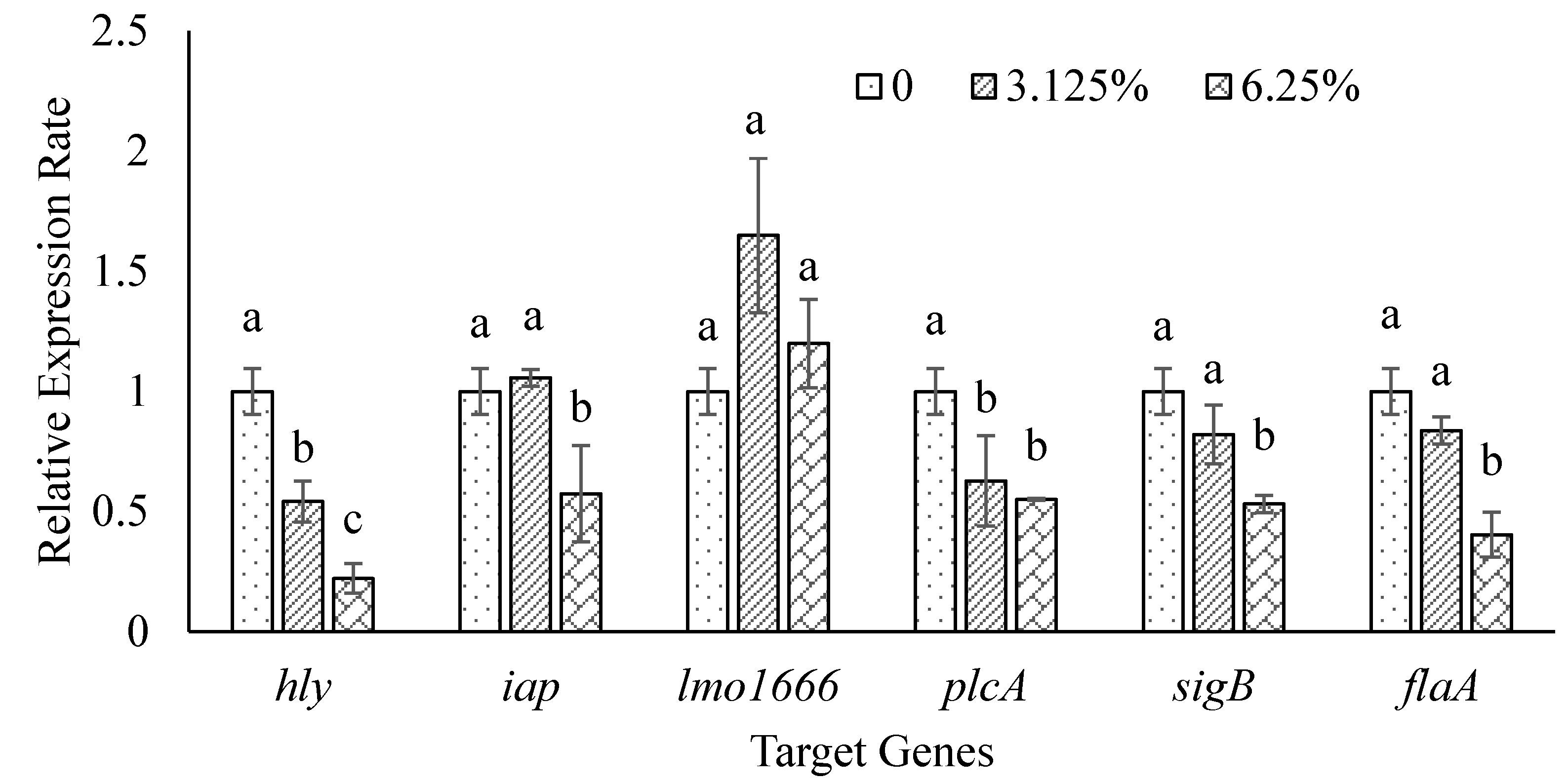
3.4. Effect of Ohelo Berry Juice on Gene Expression of L. monocytogenes
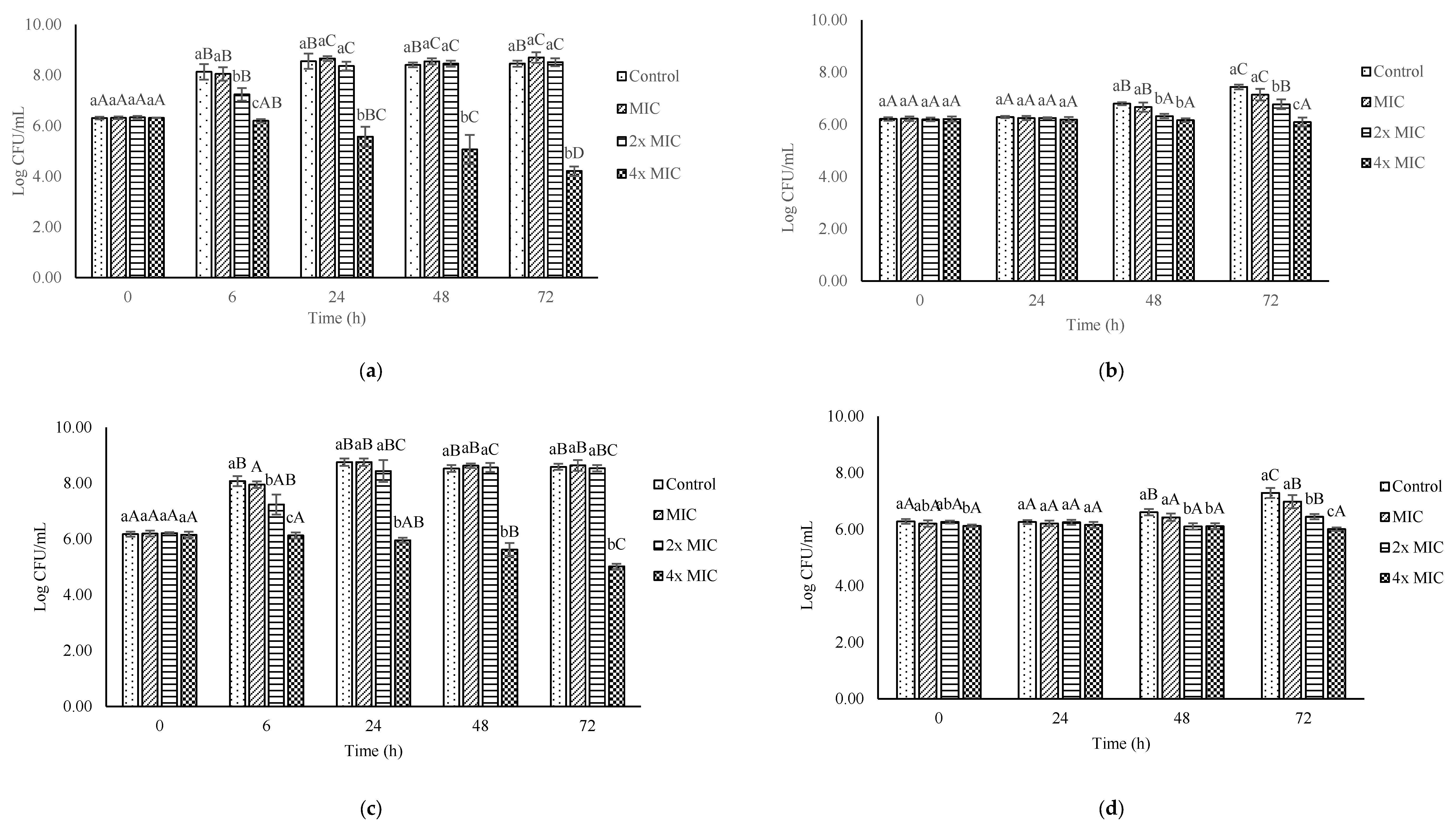
3.5. Effect of Ohelo Berry Juice on the Growth of L. monocytogenes in Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Listeria (Listeriosis). Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/listeria/risk.html (accessed on 27 May 2021).
- Chen, H.; Zhong, Q. Lactobionic acid enhances the synergistic effect of nisin and thymol against Listeria monocytogenes Scott A in tryptic soy broth and milk. Int. J. Food Microbiol. 2017, 260, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Cossart, P.; Bierne, H. The use of host cell machinery in the pathogenesis of Listeria monocytogenes. Curr. Opin. Immunol. 2001, 13, 96–103. [Google Scholar] [CrossRef]
- Interagency Food Safety Analytics Collaboration (IFSAC). Foodborne Illness Source Attribution Estimates for 2019 for Salmonella, Escherichia Coli O157, Listeria Monocytogenes and Campylobacter Using Multi-Year Outbreak Surveillance Data, United States; GA and D.C. U.S. Department of Health and Human Services’ Centers for Disease Control and Prevention and U.S. Food and Drug Administration, U.S. Department of Agriculture’s Food Safety and Inspection Service: Atlanta, GA, USA, 2021. Available online: https://www.cdc.gov/foodsafety/ifsac/annual-reports.html (accessed on 29 January 2022).
- Das, Q.; Islam, M.R.; Marcone, M.F.; Warriner, K.; Diarra, M.S. Potential of berry extracts to control foodborne pathogens. Food Control 2017, 73, 650–662. [Google Scholar] [CrossRef]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef]
- Dellavalle, C.T.; Daniel, C.R.; Aschebrook-Kilfoy, B.; Hollenbeck, A.R.; Cross, A.J.; Sinha, R.; Ward, M.H. Dietary intake of nitrate and nitrite and risk of renal cell carcinoma in the NIH-AARP Diet and Health Study. Br. J. Cancer 2013, 108, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Baenas, N.; Ruales, J.; Moreno, D.A.; Barrio, D.A.; Stinco, C.M.; Martínez-Cifuentes, G.; Meléndez-Martínez, A.J.; García-Ruiz, A. Characterization of Andean Blueberry in Bioactive Compounds, Evaluation of Biological Properties, and in vitro Bioaccessibility. Foods 2020, 9, 1483. [Google Scholar] [CrossRef]
- Lacombe, A.; McGivney, C.; Tadepalli, S.; Sun, X.; Wu, V.C. The effect of American cranberry (Vaccinium macrocarpon) constituents on the growth inhibition, membrane integrity, and injury of Escherichia coli O157:H7 and Listeria monocytogenes in comparison to Lactobacillus rhamnosus. Food Microbiol. 2013, 34, 352–359. [Google Scholar] [CrossRef]
- Lau, A.T.Y.; Barbut, S.; Ross, K.; Diarra, M.S.; Balamurugan, S. The effect of cranberry pomace ethanol extract on the growth of meat starter cultures, Escherichia coli O157: H7, Salmonella enterica serovar Enteritidis and Listeria monocytogenes. LWT 2019, 115, 108452. [Google Scholar] [CrossRef]
- Sun, X.H.; Zhou, T.T.; Wei, C.H.; Lan, W.Q.; Zhao, Y.; Pan, Y.J.; Wu, V.C. Antibacterial effect and mechanism of anthocyanin rich Chinese wild blueberry extract on various foodborne pathogens. Food Control 2018, 94, 155–161. [Google Scholar] [CrossRef]
- Côté, J.; Caillet, S.; Doyon, G.; Dussault, D.; Sylvain, J.F.; Lacroix, M. Antimicrobial effect of cranberry juice and extracts. Food Control 2011, 22, 1413–1418. [Google Scholar] [CrossRef]
- Zhao, Y.; Jiang, Q. Roles of the polyphenol-gut microbiota interaction in alleviating colitis and preventing colitis-associated colorectal cancer. Adv. Nutr. 2021, 12, 546–565. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Su, X.; Zubik, L.; Bose, P. Phenol antioxidant quantity and quality in foods: Fruits. J. Agric. Food Chem. 2001, 49, 5315–5321. [Google Scholar] [CrossRef] [PubMed]
- Alshaibani, D.; Zhang, R.; Wu, V.C. Antibacterial characteristics and activity of Vaccinium macrocarpon proanthocyanidins against diarrheagenic Escherichia coli. J. Funct. Foods 2017, 39, 133–138. [Google Scholar] [CrossRef]
- Degener, O. Plants of Hawaii National Park Illustrative of Plants and Customs of the South Seas; Food and Agriculture Organization of the United Nations: Rome, Italy; Braun-Brumfield, Inc.: Ann Arbor, MI, USA, 1973; pp. 240–245. [Google Scholar]
- Zee, F.; Keith, L.; Follett, P.; Bassil, N.; Reed, B.; Strauss, A.; Arakawa, C.; Kawabata, A.; Hamasaki, R.; Nakamoto, S.T.; et al. Kīlauea and Red Button: Two ʻŌhelo, Vaccinium Reticulatum, Cultivars from Hawaiʻi; College of Tropical Agriculture and Human Resources, University of Hawaiʻi at Mānoa: Honolulu, Hawaii, 2011; Available online: https://www.ctahr.hawaii.edu/oc/freepubs/pdf/F_N-18.pdf (accessed on 29 January 2022).
- Hummer, K.; Durst, R.; Zee, F.; Atnip, A.; Giusti, M.M. Phytochemicals in fruits of Hawaiian wild cranberry relatives. J. Sci. Food Agric. 2014, 94, 1530–1536. [Google Scholar] [CrossRef]
- Hong, V.; Wrolstad, R.E. Cranberry juice composition. J. Assoc. Off. Anal. Chem. 1986, 69, 199–207. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Acree, T.E.; Decker, E.A.; Penner, M.H.; Reid, D.S.; Schwartz, S.J.; Shoemaker, C.F.; Smith, D.M.; Sporns, P. Handbook of Food Analytical Chemistry, Volume 2: Pigments, Colorants, Flavors, Texture, and Bioactive Food Components; John Wiley & Sons: Hoboken, NJ, USA, 2005; pp. 457–537. [Google Scholar]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically. Approved Standard —Eighth Edition; CLSI publication M07-A8; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2009; Available online: https://simpleshowoflove.weebly.com/uploads/1/4/0/7/14073276/agar_dilution_assay.pdf (accessed on 29 January 2022).
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef] [Green Version]
- Lacombe, A.; Wu, V.C.; White, J.; Tadepalli, S.; Andre, E.E. The antimicrobial properties of the lowbush blueberry (Vaccinium angustifolium) fractional components against foodborne pathogens and the conservation of probiotic Lactobacillus rhamnosus. Food Microbiol. 2012, 30, 124–131. [Google Scholar] [CrossRef]
- Salaheen, S.; Nguyen, C.; Hewes, D.; Biswas, D. Cheap extraction of antibacterial compounds of berry pomace and their mode of action against the pathogen Campylobacter jejuni. Food Control 2014, 46, 174–181. [Google Scholar] [CrossRef]
- O’May, C.; Tufenkji, N. The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Environ. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. JoVE 2011, 47, 2437. [Google Scholar] [CrossRef]
- Wu, V.C.; Qiu, X.; de los Reyes, B.G.; Lin, C.S.; Pan, Y. Application of cranberry concentrate (Vaccinium macrocarpon) to control Escherichia coli O157:H7 in ground beef and its antimicrobial mechanism related to the downregulated slp, hdeA and cfa. Food Microbiol. 2009, 26, 32–38. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Upadhyay, A.; Johny, A.K.; Amalaradjou, M.A.; Baskaran, S.A.; Kim, K.S.; Venkitanarayanan, K. Plant-derived antimicrobials reduce Listeria monocytogenes virulence factors in vitro, and down-regulate expression of virulence genes. Int. J. Food Microbiol. 2012, 157, 88–94. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Li, H.; Guo, X.; Guo, D.; Yang, Y.; Wang, X.; Zhang, C.; Shan, Z.; Xia, X.; et al. Antibiofilm activity of shikonin against Listeria monocytogenes and inhibition of key virulence factors. Food Control 2021, 120, 107558. [Google Scholar] [CrossRef]
- Yan, Z.; Gurtler, J.B.; Kornacki, J.L. A solid agar overlay method for recovery of heat-injured Listeria monocytogenes. J. Food Prot. 2006, 69, 428–431. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research.R Package Version 1.4.0; 2020. Available online: https://myaseen208.github.io/agricolae/https://cran.r-project.org/package=agricolae (accessed on 21 January 2022).
- Lee, S.H.I.; Cappato, L.P.; Guimarães, J.T.; Balthazar, C.F.; Rocha, R.S.; Franco, L.T.; da Cruz, A.G.; Corassin, C.H.; de Oliveira, C.A.F. Listeria monocytogenes in milk: Occurrence and recent advances in methods for inactivation. Beverages 2019, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.C.; de Sousa, C.P.; Fernandez-Prada, C.; Harel, J.; Dubreuil, J.D.; de Souza, E.L. A review of the current evidence of fruit phenolic compounds as potential antimicrobials against pathogenic bacteria. Microb. Pathog. 2019, 130, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.B.; Kyle, J.; Jenkinson, A.M.; Gardner, P.T.; McPhail, D.B.; Duthie, G.G. Effects of blueberry and cranberry juice consumption on the plasma antioxidant capacity of healthy female volunteers. Eur. J. Clin. Nutr. 2000, 54, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Vinson, J.A.; Bose, P.; Proch, J.; Al Kharrat, H.; Samman, N. Cranberries and cranberry products: Powerful in vitro, ex vivo, and in vivo sources of antioxidants. J. Agric. Food Chem. 2008, 56, 5884–5891. [Google Scholar] [CrossRef]
- Lee, J.; Rennaker, C.; Wrolstad, R.E. Correlation of two anthocyanin quantification methods: HPLC and spectrophotometric methods. Food Chem. 2008, 110, 782–786. [Google Scholar] [CrossRef]
- Nohynek, L.J.; Alakomi, H.L.; Kähkönen, M.P.; Heinonen, M.; Helander, I.M.; Oksman-Caldentey, K.M.; Puupponen-Pimiä, R.H. Berry phenolics: Antimicrobial properties and mechanisms of action against severe human pathogens. Nutr. Cancer 2006, 54, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Perez, C.; Wei, Y.; Rapoza, E.; Su, G.; Bou-Abdallah, F.; Chasteen, N.D. Iron-binding properties of plant phenolics and cranberry’s bio-effects. Dalton Trans. 2007, 43, 4951–4961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond risk: Bacterial biofilms and their regulating approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Song, X.; Zhang, Z.; Fu, J.; Wang, X.; Malakar, P.K.; Liu, H.; Pan, Y.; Zhao, Y. Removal of foodborne pathogen biofilms by acidic electrolyzed water. Front. Microbiol. 2017, 8, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dushku, E.; Kotzamanidis, C.; Avgousti, K.; Zdragas, A.; Vafeas, G.; Giantzi, V.; Staikou, A.; Yiangou, M. Listeria monocytogenes induced dysbiosis in snails and rebiosis achieved by administration of the gut commensal Lactobacillus plantarum Sgs14 strain. Fish Shellfish. Immunol. 2020, 104, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Qiao, J.; Lu, Z.; Fen, Z.; Tao, Y.; Lv, F.; Zhao, H.; Zhang, C.; Bie, X. Influence of different factors on biofilm formation of Listeria monocytogenes and the regulation of cheY gene. Food Res. Int. 2020, 137, 109405. [Google Scholar] [CrossRef]
- Choi, N.Y.; Bae, Y.M.; Lee, S.Y. Cell surface properties and biofilm formation of pathogenic bacteria. Food Sci. Biotechnol. 2015, 24, 2257–2264. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]
- Wang, Y.; Baptist, J.A.; Dykes, G.A. Garcinia mangostana extract inhibits the attachment of chicken isolates of Listeria monocytogenes to cultured colorectal cells potentially due to a high proanthocyanidin content. J. Food Saf. 2021, 41, e12889. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yu, H.H.; Song, Y.J.; Park, Y.J.; Lee, N.K.; Paik, H.D. Anti-biofilm effect of the cell-free supernatant of probiotic Saccharomyces cerevisiae against Listeria monocytogenes. Food Control 2021, 121, 107667. [Google Scholar] [CrossRef]
- Lemon, K.P.; Higgins, D.E.; Kolter, R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J. Bacteriol. 2007, 189, 4418–4424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dons, L.; Eriksson, E.; Jin, Y.; Rottenberg, M.E.; Kristensson, K.; Larsen, C.N.; Bresciani, J.; Olsen, J.E. Role of flagellin and the two-component CheA/CheY system of Listeria monocytogenes in host cell invasion and virulence. Infect. Immun. 2004, 72, 3237–3244. [Google Scholar] [CrossRef] [Green Version]
- Hamon, M.A.; Ribet, D.; Stavru, F.; Cossart, P. Listeriolysin O: The Swiss army knife of Listeria. Trends Microbiol. 2012, 20, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Hadjilouka, A.; Molfeta, C.; Panagiotopoulou, O.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Expression of Listeria monocytogenes key virulence genes during growth in liquid medium, on rocket and melon at 4, 10 and 30 C. Food Microbiol. 2016, 55, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.A.; Wilke, T.; Erdmann, R. Efficacy of bacteriocin-containing cell-free culture supernatants from lactic acid bacteria to control Listeria monocytogenes in food. Int. J. Food Microbiol. 2011, 146, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hewes, D.; Salaheen, S.; Federman, C.; Biswas, D. Effects of blackberry juice on growth inhibition of foodborne pathogens and growth promotion of Lactobacillus. Food Control 2014, 37, 15–20. [Google Scholar] [CrossRef]
- Biswas, D.; Wideman, N.E.; O’Bryan, C.A.; Muthaiyan, A.; Lingbeck, J.M.; Crandall, P.G.; Ricke, S.C. Pasteurized blueberry (Vaccinium corymbosum) juice inhibits growth of bacterial pathogens in milk but allows survival of probiotic bacteria. J. Food Saf. 2012, 32, 204–209. [Google Scholar] [CrossRef]
- Jung, D.S.; Bodyfelt, F.W.; Daeschel, M.A. Influence of fat and emulsifiers on the efficacy of nisin in inhibiting Listeria monocytogenes in fluid milk. J. Dairy Sci. 1992, 75, 387–393. [Google Scholar] [CrossRef]
- Sun, L.; Forauer, E.C.; Brown, S.; D’Amico, D.J. Application of bioactive glycolipids to control Listeria monocytogenes biofilms and as post-lethality contaminants in milk and cheese. Food Microbiol. 2021, 95, 103683. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence | Locus | Protein/Function | Reference | Efficiency 1 |
|---|---|---|---|---|---|---|
| 16S rRNA | Forward | 5′-TGGCGGACGGGTGAGTA-3′ | lmor01 | Reference gene | Upadhyay et al. [30] | 2.05 |
| Reverse | 5′-CCGGAGTTATCCCCAACTTACA-3′ | |||||
| hly | Forward | 5′-TCTCCGCCTGCAAGTCCTA-3′ | lmo0202 | Listeriolysin O | Upadhyay et al. [30] | 2.06 |
| Reverse | 5′-TCGATTTCATCCGCGTGTT-3′ | |||||
| iap | Forward | 5′-CTACAGCTGGGATTGCGGTAA-3′ | lmo0582 | Invasion-associated protein | Upadhyay et al. [30] | 1.99 |
| Reverse | 5′-TGCTTGCGGATGCGATT-3′ | |||||
| lmo1666 | Forward | 5′-TGGAGTGGGCACGTGTTGT-3′ | lmo1666 | Adhesion protein B | Upadhyay et al. [30] | 2.03 |
| Reverse | 5′-TTGTCAGCTGCATATTGTGAATTG-3′ | |||||
| plcA | Forward | 5′-TCGGACCATTGTAGTCATCTTGA-3′ | lmo0201 | Phospholipase C | Upadhyay et al. [30] | 1.97 |
| Reverse | 5′-CACAAATTCGGCATGCAGTT-3′ | |||||
| sigB | Forward | 5’-GATGATGGATTTGAACGTGTGAA-3’ | lmo0895 | Biofilm formation and disinfectant resistance | Li et al. [31] | 1.99 |
| Reverse | 5’-CGCTCATCTAAAACAGGAGAAC-3’ | |||||
| flaA | Forward | 5’-CTGGTATGAGTCGCCTTAG-3’ | lmo0690 | Flagellin | Li et al. [31] | 1.96 |
| Reverse | 5’-CATTTGCGGTGTTTGGTTTG-3’ |
| Concentration | pH |
|---|---|
| 100% | 3.27 ± 0.03 a 1 |
| 50% | 3.86 ± 0.04 b |
| 25% | 4.24 ± 0.02 c |
| 12.5% | 4.69 ± 0.04 d |
| 6.25% | 5.40 ± 0.01 e |
| 3.12% | 6.37 ± 0.02 f |
| 1.56% | 6.85 ± 0.05 g |
| 0.78% | 7.09 ± 0.01 h |
| Ohelo Berry Juice (%) | Hydrophobicity (%) | Auto-Aggregation (%) | Swimming Motility 2 (%) | Swarming Motility 2 (%) | Biofilm Formation 2 (%) |
|---|---|---|---|---|---|
| 0 | 7.71 ± 0.85 a | 16.02 ± 2.83 a | 100 a | 100 a | 100 a |
| 3.12 | 2.77 ± 0.46 b | 23.95 ± 3.83 a | 41.29 ± 3.00 b | 22.80 ± 0.31 b | 54.33 ± 0.39 b |
| 6.25 | 2.14 ± 0.87 b | 67.98 ± 10.92 b | 4.39 ± 0.17 c | 14.71 ± 0.23 b | 23.00 ± 0.53 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, B.; Liu, X.; Nakamoto, S.T.; Wall, M.; Li, Y. Antimicrobial Activity of Ohelo Berry (Vaccinium calycinum) Juice against Listeria monocytogenes and Its Potential for Milk Preservation. Microorganisms 2022, 10, 548. https://doi.org/10.3390/microorganisms10030548
Wu B, Liu X, Nakamoto ST, Wall M, Li Y. Antimicrobial Activity of Ohelo Berry (Vaccinium calycinum) Juice against Listeria monocytogenes and Its Potential for Milk Preservation. Microorganisms. 2022; 10(3):548. https://doi.org/10.3390/microorganisms10030548
Chicago/Turabian StyleWu, Biyu, Xiaohan Liu, Stuart T. Nakamoto, Marisa Wall, and Yong Li. 2022. "Antimicrobial Activity of Ohelo Berry (Vaccinium calycinum) Juice against Listeria monocytogenes and Its Potential for Milk Preservation" Microorganisms 10, no. 3: 548. https://doi.org/10.3390/microorganisms10030548