
Identification, Characterization, and Virulence Gene Expression of Marine Enterobacteria in the Upper Gulf of Thailand


Abstract
:1. Introduction
2. Materials and Methods
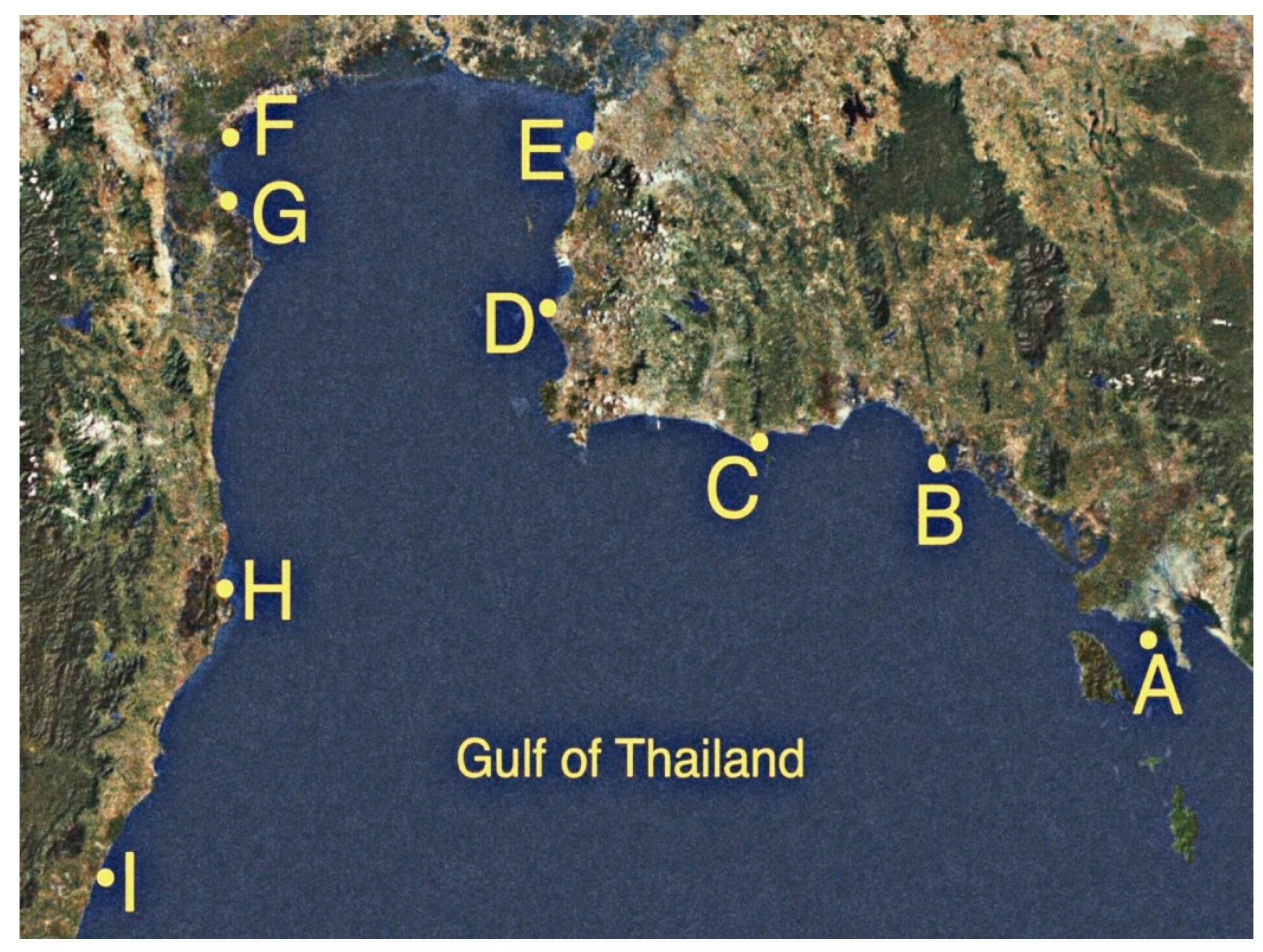
2.1. Isolation of Enterobacteria from Seawater Samples
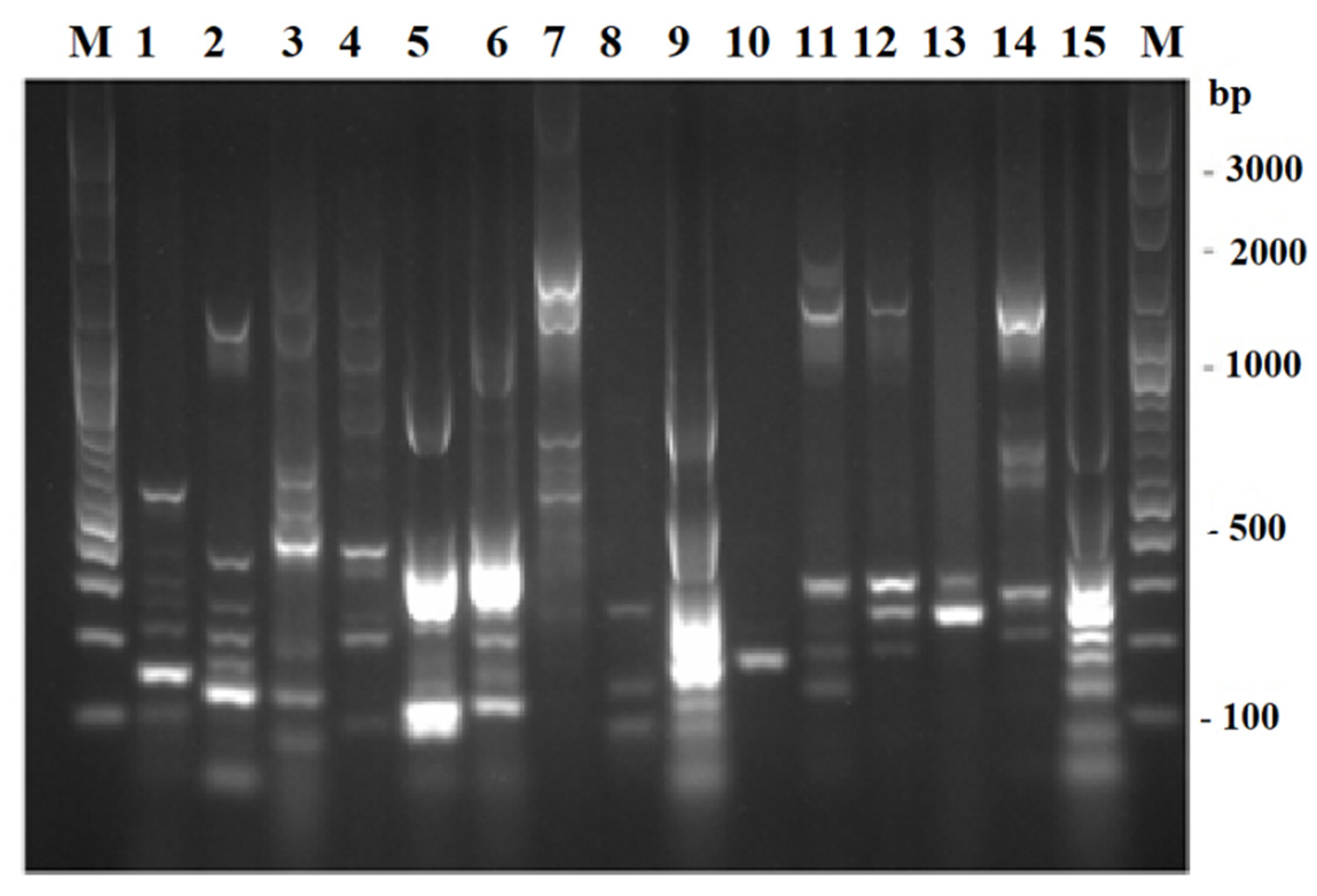
2.2. ERIC-PCR Fingerprinting of Presumptive Enterobacterial Isolates
2.3. TP-RAPD Fingerprinting of Presumptive Enterobacterial Strains and Species Identification by Using the Vitek® MS System
2.4. Examination on Antibiotic Resistance of Enterobacterial Strains
2.5. Detection and Sequence Analysis of Antibiotic Resistance and Virulence Genes in Enterobacterial Strains
2.6. Statistical Analysis for Association between Antibiotic Resistance Genes and Virulence Genes
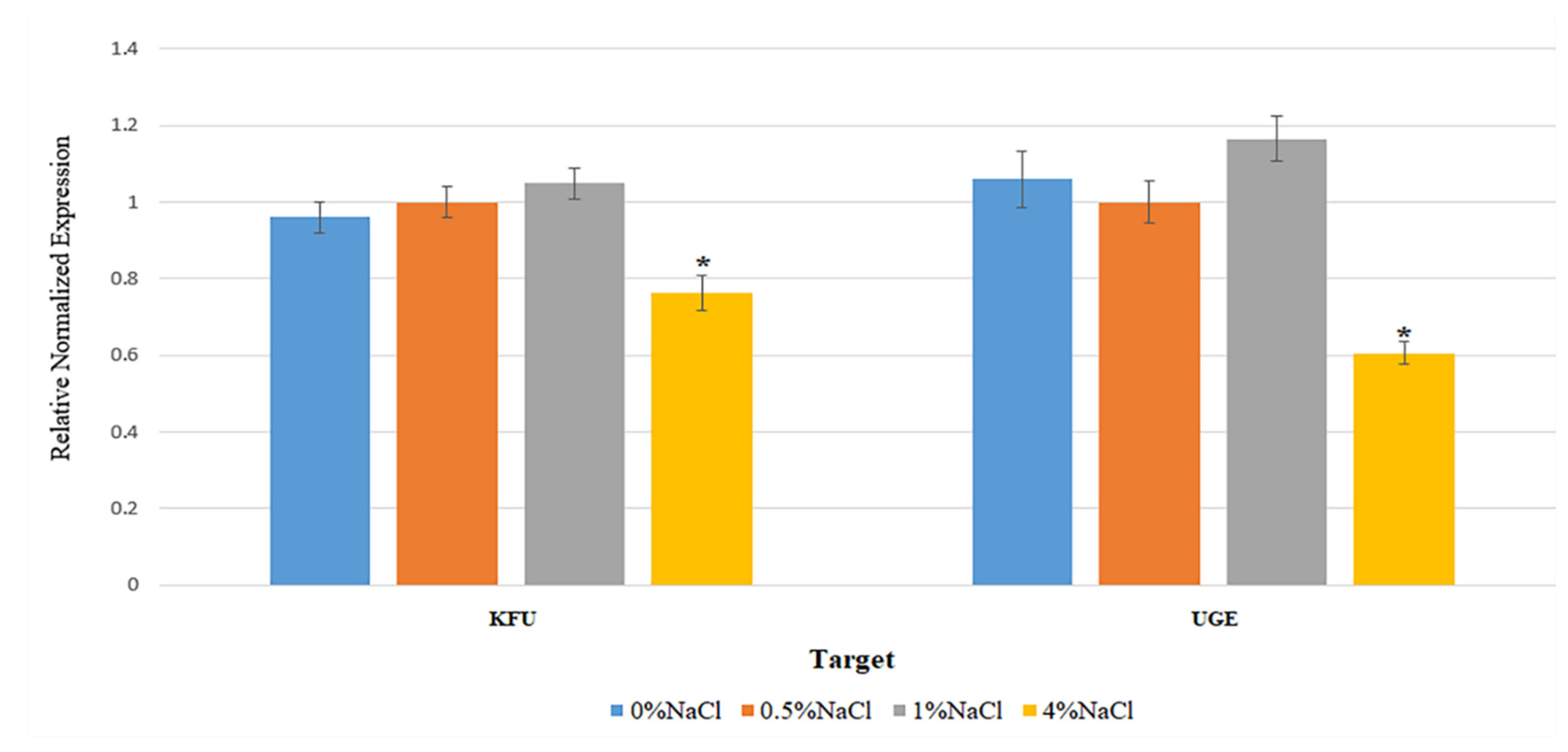
2.7. Examination on Factors Affecting the Expression of Virulence Genes
2.8. Examination on Nanoparticle Synthesis by Enterobacterial Strains
3. Results
3.1. Prevalence of Enterobacteria in Seawater along the Upper Gulf of Thailand
3.2. ERIC-PCR Fingerprinting of Presumptive Enterobacterial Isolates
3.3. Distinguishing Presumptive Enterobacterial Species Based on TP-RAPD Fingerprinting
3.4. Species Identification of Presumptive Enterobacterial Strains Based on the Vitek® MS System
3.5. Prevalence of Antibiotic Resistance among Enterobacterial Strains
3.6. Detected Antibiotic Resistance Genes in Enterobacterial Strains
3.7. Detected Virulence Genes in Enterobacterial Strains
3.8. Association of Antibiotic Resistance Genes and Virulence Genes
3.9. Synthesis of Silver and Gold Nanoparticles by Enterobacterial Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nimnoi, P.; Pongsilp, N. Marine bacterial communities in the upper gulf of Thailand assessed by Illumina next-generation sequencing platform. BMC Microbiol. 2020, 20, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [PubMed]
- Morales-López, S.; Yepes, J.A.; Prada-Herrera, J.C.; Torres-Jiménez, A. Enterobacteria in the 21st century: A review focused on taxonomic changes. J. Infect. Dev. Ctries 2019, 13, 265–273. [Google Scholar] [CrossRef]
- Baldelli, V.; Scaldaferri, F.; Putignani, L.; Del Chierico, F. The role of Enterobacteriaceae in gut microbiota dysbiosis in inflammatory bowel diseases. Microorganisms 2021, 9, 697. [Google Scholar] [CrossRef]
- Biran, D.; Ron, E.Z. Extraintestinal pathogenic Escherichia coli. Curr. Top. Microbiol. Immunol. 2018, 416, 149–161. [Google Scholar] [PubMed]
- Sarkar, S.; Kimono, D.; Albadrani, M.; Seth, R.K.; Busbee, P.; Alghetaa, H.; Porter, D.E.; Scott, G.I.; Brooks, B.; Nagarkatti, M.; et al. Environmental microcystin targets the microbiome and increases the risk of intestinal inflammatory pathology via NOX2 in underlying murine model of nonalcoholic fatty liver disease. Sci. Rep. 2019, 9, 8742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, P.; Messina, N.; Mohn, W.W.; Finlay, B.B.; Curtis, N. Association between the intestinal microbiota and allergic sensitization, eczema, and asthma: A systematic review. J. Allergy Clin. Immunol. 2019, 143, 467–485. [Google Scholar] [CrossRef]
- Wen, X.; Cgen, F.; Lin, Y.; Zhu, H.; Yuan, F.; Kuang, D.; Jia, Z.; Yuan, Z. Microbial indicators and their use for monitoring drinking water quality: A review. Sustainability 2020, 12, 2249. [Google Scholar] [CrossRef] [Green Version]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Gonzalez-Martinez, A.; Gonzalez-Lopez, J.; Munoz-Palazon, B.; Fenice, M. Persistence of Enterobacteriaceae drawn into a marine saltern (Saline di Tarquinia, Italy) from the adjacent coastal zone. Water 2021, 13, 1443. [Google Scholar] [CrossRef]
- Cohen, R.; Paikin, S.; Astrahan, P. 1619 carbapenemase producing Enterobacteriaceae in river estuaries and coastal water of Netanya, Israel. Open Forum Infect. Dis. 2019, 6, S590–S591. [Google Scholar] [CrossRef]
- Mahon, B.M.; Brehony, C.; Cahill, N.; McGrath, E.; O’Conor, L.; Varley, A.; Cormican, M.; Ryan, S.; Hickey, P.; Keane, S.; et al. Detection of OXA-48-like-producing Enterobacterales in Irish recreational water. Sci. Total Environ. 2019, 690, 1–6. [Google Scholar] [CrossRef]
- Mahon, B.M.; Brehony, C.; McGrath, E.; Killeen, J.; Cormican, M.; Hickey, P.; Keane, S.; Hanahoe, B.; Dolan, A.; Morris, D. Indistinguishable NDM-producing Escherichia coli isolated from recreational waters, sewage, and a clinical specimen in Ireland, 2016 to 2017. Euro. Surveill. 2017, 22, 30513. [Google Scholar] [CrossRef] [Green Version]
- Paschoal, R.P.; Campana, E.H.; Corrêa, L.L.; Montezzi, L.F.; Barrueto, L.R.L.; da Silva, I.R.; Bonelli, R.R.; Castro, L.S.; Picao, R.C. Concentration and variety of carbapenemase producers in recreational coastal waters showing distinct levels of pollution. Antimicrob. Agents Chemother. 2017, 61, e01963-17. [Google Scholar] [CrossRef] [Green Version]
- Abdelraouf, A.E.; Philippe, H.; Kamal, J.E.; Adnan, A.; Samir, A.; Fatma, E.; Alaa, R.A. Antimicrobial resistance of Staphylococcus aureus, fecal streptococci, Enterobacteriaceae and Pseudomonas aeruginosa isolated from the coastal water of the Gaza strip-Palestine. Int. Arab. J. Antimicrob. Agents. 2016, 6, 3. [Google Scholar] [CrossRef]
- Farkas, A.; Tarco, E.; Butiuc-Keul, A. Antibiotic resistance profiling of pathogenic Enterobacteriaceae from Cluj-Napoca, Romania. Germs 2019, 9, 17–27. [Google Scholar] [CrossRef]
- Santos, A.L.; Dos Santos, A.P.; Ito, C.R.M.; Queiroz, P.H.P.; de Almeida, J.A.; de Carvalho Júnior, M.A.B.; de Oliveira, C.Z.; Avelino, M.A.G.; Wastowski, I.J.; Gomes, G.P.L.A.; et al. Profile of enterobacteria resistant to beta-lactams. Antibiotics 2020, 9, 410. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.S.; Pereira, A.; Araujo, S.M.; Castro, B.B.; Correia, A.C.M.; Henriques, I. Seawater is a reservoir of multi-resistant Escherichia coli, including strains hosting plasmid-mediated quinolones resistance and extended-spectrum beta-lactamases genes. Front. Microbiol. 2014, 5, 426. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Paikin, S.; Rokney, A.; Rubin-Blum, M.; Astrahan, P. Multidrug-resistant enterobacteriaceae in coastal water: An emerging threat. Antimicrob. Resist. Infect. Control 2020, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Maloo, A.; Borade, S.; Dhawde, R.; Gajbhiye, S.N.; Dastager, S.G. Occurrence and distribution of multiple antibiotic-resistantbacteria of Enterobacteriaceae family in waters of Veraval coast, India. Environ. Exp. Bot. 2014, 12, 43–50. [Google Scholar]
- Partridge, S.R. Resistance mechanisms in Enterobacteriaceae. Pathology 2015, 47, 270–284. [Google Scholar] [CrossRef]
- Nappier, S.P.; Liguori, K.; Ichida, A.M.; Stewart, J.R.; Jones, K.R. Antibiotic resistance in recreational waters: State of the science. Int. J. Environ. Res. Pub. Health 2020, 17, 8034. [Google Scholar] [CrossRef]
- Gupta, M.; Naik, A.K.; Singh, S.K. Bacteriological profile and antimicrobial resistance patterns of burn wound infections in a tertiary care hospital. Heliyon 2019, 5, e02956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Gao, H.; Zheng, L.; Liu, S.; Cao, Y.; Zhu, S.; Wu, Z.; Ren, H.; Mao, D.; Luo, Y. Antibiotic resistance and virulence of extraintestinal pathogenic Escherichia coli (ExPEC) vary according to molecular types. Front. Microbiol. 2020, 11, 598305. [Google Scholar] [CrossRef] [PubMed]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizon, S.; de Roode, J.C.; Michalakis, Y. Multiple infections and the evolution of virulence. Ecol. Lett. 2013, 16, 556–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-PLoSkonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Wassenaar, T.M.; Gaastra, W. Bacterial virulence: Can we draw the line? FEMS Microbiol. Lett. 2001, 201, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, K.; Garai, P.; Chatterjee, J.; Chakravortty, D. Peptide transporter YjiY influences the expression of the virulence gene mgtC to regulate biofilm formation in Salmonella. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Nimnoi, P.; Pongsilp, N. Distribution and expression of virulence genes in potentially pathogenic bacteria isolated from seafood in Thailand. CyTA J. Food. 2020, 18, 753–763. [Google Scholar] [CrossRef]
- Paytubi, S.; Cansado, C.; Madrid, C.; Balsalobre, C. Nutrient composition promotes switching between pellicle and bottom biofilm in Salmonella. Front. Microbiol. 2017, 8, 2160. [Google Scholar] [CrossRef]
- Gerstel, U.; Romling, U. Oxygen tension and nutrient starvation are major signals that regulate agfD promoter activity and expression of the multicellular morphotype in Salmonella typhimurium. Environ. Microbiol. 2001, 3, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Jubelin, G.; Vianney, A.; Beloin, C.; Ghigo, J.M.; Lazzaroni, J.C.; Lejeune, P.; Dorel, C. CpxR/OmpR interplay regulates curli gene expression in response to osmolarity in Escherichia coli. J. Bacteriol. 2005, 187, 2038–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, N.; Jespersen, L. Expression of virulence-related genes in Listeria monocytogenes grown on Danish hard cheese as affected by NaCl content. Foodborne Pathog. Dis. 2015, 12, 536–544. [Google Scholar] [CrossRef]
- Prignent-Combaret, C.; Brombacher, E.; Vidal, O.; Ambert, A.; Lejeune, P.; Landini, P.; Dorel, C. Complex regulatory network controls initial adhesion and biofilm formation in Escherichia coli via regulation of the csgD gene. J. Bacteriol. 2001, 183, 7213–7223. [Google Scholar] [CrossRef] [Green Version]
- Valdes, K.M.; Sundar, G.S.; Belew, A.T.; Islam, E.; El-Sayed, N.M.; Breton, Y.L.; Mclver, K.S. Glucose levels alter the Mga virulence regulon in the group A. Streptococcus. Sci. Rep. 2018, 8, 4971. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Li, L.J.; Shi, Q.S.; Ouyang, Y.S.; Chen, Y.B.; Hu, W.F. Effects of nutritional and environmental conditions on planktonic growth and biofilm formation of Citrobacter werkmanii BF-6. J. Microbiol. Biotechnol. 2013, 23, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Melson, E.M.; Kendall, M.M. The sRNA DicF integrates oxygen sensing to enhance enterohemorrhagic Escherichia coli virulence via distinctive RNA control mechanisms. Proc. Natl. Acad. Sci. USA 2019, 116, 14210–14215. [Google Scholar] [CrossRef] [Green Version]
- Guijarro, J.A.; Cascales, D.; Garcia-Torrico, A.I.; Garcia-Dominguez, M.; Mendez, J. Temperature-dependent expression of virulence genes in fish-pathogenic bacteria. Front. Microbiol. 2015, 6, 700. [Google Scholar] [CrossRef]
- Koohsari, H.; Ghaemi, E.A.; Mozaffari, N.A.; Moradi, A.; Sadegh-Sheshpoli, M.; Javid, S.N. The effect of adding blood on the virulence genes expression of Staphylococcus aureus in exponential and stationary growth phase. Jundishapur J. Microbiol. 2017, 10, e14380. [Google Scholar] [CrossRef]
- Ogasawara, H.; Yamada, K.; Kori, A.; Yamamoto, K.; Ishihama, A. Regulation of the Escherichia coli csgD promoter: Interplay between five transcription factors. Microbiology 2010, 156, 2470–2483. [Google Scholar] [CrossRef] [Green Version]
- Ferrando, M.L.; van Baarlen, P.; Orru, G.; Piga, R.; Bongers, R.S.; Wels, M.; De Greeff, A.; Smith, H.E.; Wells, J.M. Carbohydrate availability regulates virulence gene expression in Streptococcus suis. PLoS ONE 2014, 9, e89334. [Google Scholar] [CrossRef] [Green Version]
- Kentache, T.; Milochanic, E.; Cao, T.N.; Mokhtari, A.; Ake, F.M.; Ma Pham, Q.M.; Joyet, P.; Deutscher, J. Transport and catabolism of pentitols by Listeria monocytogenes. J. Mol. Microbiol. Biotechnol. 2016, 26, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Makthal, N.; VanderWal, A.R.; Saavedra, M.O.; Olsen, R.J.; Musser, J.M.; Kumaraswami, M. Environmental pH and peptide signaling control virulence of Streptococcus pyogenes via a quorum-sensing pathway. Nat. Commun. 2019, 10, 2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joffre, E.; Nicklasson, M.; Alvarez-Carretero, S.; Xiao, X.; Sun, L.; Nookaew, I.; Zhu, B.; Sjoling, A. The bile salt glycocholate induces global changes in gene and protein expression and activates virulence in enterotoxigenic Escherichia coli. Sci. Rep. 2019, 9, 108. [Google Scholar] [CrossRef]
- Haber, A.; Friedman, S.; Lobel, L.; Burg-Golani, T.; Sigal, N.; Rose, J.; Livnat-Levanon, N.; Lewinson, O.; Herskovits, A.A. L-glutamate induces expression of Listeria monocytogenes virulence genes. PLoS Pathog. 2017, 13, e1006161. [Google Scholar] [CrossRef] [PubMed]
- Ayesha, A. Bacterial synthesis and applications of nanoparticles. Nano. Sci. Nano. Technol. 2017, 11, 119. [Google Scholar]
- Koul, B.; Poonia, A.K.; Yadav, D.; Jin, J.O. Microbe-mediated biosynthesis of nanoparticles: Applications and future prospects. Biomolecules 2021, 11, 886. [Google Scholar] [CrossRef]
- Sadeghi-Aghbash, M.; Rahimnejad, M.; Pourali, S.M. Bio-mediated synthesis and characterization of zinc phosphate nanoparticles using Enterobacter aerogenes cells for antibacterial and anticorrosion applications. Curr. Pharm. Biotechnol. 2020, 21, 1232–1241. [Google Scholar] [CrossRef]
- Srinath, B.S.; Rai, V.R. Biosynthesis of highly monodispersed, spherical gold nanoparticles of size 4-10 nm from spent cultures of Klebsiella pneumoniae. 3 Biotech 2015, 5, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Honary, S.; Gharaei-Fathabad, E.; Paji, Z.K.; Eslamifar, M. A novel biological synthesis of gold nanoparticle by Enterobacteriaceae family. Trop. J. Pharm. Res. 2012, 11, 887–891. [Google Scholar] [CrossRef] [Green Version]
- Baltazar-Encarnación, E.; Escárcega-González, C.E.; Vasto-Anzaldo, X.G.; Cantú-Cárdenas, M.E.; Morones-Ramírez, J.R. Silver nanoparticles synthesized through green methods using Escherichia coli Top 10 (Ec-Ts) growth culture medium exhibit antimicrobial properties against nongrowing bacterial strains. J. Nanomater. 2019, 2019, 4637325. [Google Scholar] [CrossRef] [Green Version]
- Javaid, A.; Oloketuyi, S.; Khan, M.M.; Khan, F. Diversity of bacterial synthesis of silver nanoparticles. BioNanoSci. 2018, 8, 43–59. [Google Scholar] [CrossRef]
- Song, D.; Li, X.; Cheng, Y.; Xiao, X.; Lu, Z.; Wang, Y.; Wang, F. Aerobic biogenesis of selenium nanoparticles by Enterobacter cloacae Z0206 as a consequence of fumarate reductase mediated selenite reduction. Sci. Rep. 2017, 7, 3239. [Google Scholar] [CrossRef]
- El-Shanshoury, A.R.R.; Elsilk, S.E.; Ateya, P.S.; Ebeid, E.M. Synthesis of lead nanoparticles by Enterobacter sp. and avirulent Bacillus anthracis PS2010. Ann. Microbiol. 2012, 62, 1803–1810. [Google Scholar] [CrossRef]
- Wang, Q.F.; Li, Q.; Lin, Y.; Hou, Y.; Deng, Z.; Liu, W.; Wang, H.; Xia, Z. Biochemical and genetic basis of cadmium biosorption by Enterobacter ludwigii LY6, isolated from industrial contaminated soil. Environ. Pollut. 2020, 264, 114637. [Google Scholar] [CrossRef]
- Standard Microbiological Water Sampling Technique. Available online: https://ww2.health.wa.gov.au/~/media/Files/Corporate/general-documents/water/envwater/other-publications/PDF/Env-waters-micro-sampling-technique.pdf (accessed on 11 January 2022).
- Da Silva, N.; Taniwaki, M.H.; Junqueira, V.C.A.; Silveira, N.; Da Silva Do Nascimento, M.; Gomes, R.A.R. Microbiology Examination Methods of Food and Water: A Laboratory Manual, 1st ed.; CRC Press: London, UK, 2012; pp. 83–88. [Google Scholar]
- Walker, K.E.; Mahon, C.R.; Lehman, D.C.; Manuselis, G. Enterobacteriaceae. In Textbook of Diagnostic Microbiology, 5th ed.; Mahon, C.R., Lehman, D.C., Manuselis, G., Eds.; Saunders: Philadelphia, PA, USA, 2015; pp. 420–454. [Google Scholar]
- American Public Health Association (APHA). Standard Method for the Examination of Water and Wastewater, 22nd ed.; APHA: Washington, DC, USA, 2012; Volume 9221B. [Google Scholar]
- Ogutcu, H.; Adiguzel, A.; Gulluce, M.; Karadayi, M.; Sahin, F. Molecular characterization of Rhizobium strains isolated from wild chickpeas collected from high altitudes in Erzurum-Turkey. Rom. Biotechnol. Lett. 2009, 14, 4294–4300. [Google Scholar]
- Rivas, R.; Velazquez, E.; Valverde, A.; Mateos, P.F.; Martinez-Molina, E. A two primers random amplified polymorphic DNA procedure to obtain polymerase chain reaction fingerprints of bacterial species. Electrophoresis 2001, 22, 1086–1089. [Google Scholar] [CrossRef]
- Antimicrobial Susceptibility Testing EUCAST Disk Diffusion Method Version 2.1. Available online: https://asmsig.files.wordpress.com/2013/08/manual_v_2-1_eucast_disk_test.pdf (accessed on 15 December 2021).
- Antimicrobial Susceptibility Systems. Available online: https://www.himedialabs.com/HML/images/literature/pdf/100000027/68.pdf (accessed on 15 December 2021).
- Hanson, N.D.; Thomson, K.S.; Moland, E.S.; Sanders, C.C.; Berthold, G.; Penn, R.G. Molecular characterization of a multiply resistant Klebsiella pneumoniae encoding ESBLs and a plasmid-mediated AmpC. J. Antimicrob. Chemother. 1999, 44, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Mulvey, M.R.; Grant, J.M.; Plewes, K.; Roscoe, D.; Boyd, D.A. New Delhi metallo-β-lactamase in Klebsiella pneumoniae and Escherichia coli, Canada. Emerg. Infect. Dis. 2011, 17, 103–106. [Google Scholar] [CrossRef]
- Jang, J.; Suh, Y.S.; Di, D.Y.; Unno, T.; Sadowsky, M.J.; Hur, H.G. Pathogenic Escherichia coli strains producing extended-spectrum β-lactamase in the Yeongsan River basin of South Korea. Environ. Sci. Technol. 2013, 47, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.E.; Christensen, H.; Aarestrup, F.M. Diversity and evolution of blaZ from Staphylococcus aureus and coagulase-negative staphylococci. J. Antimicrob. Chemother. 2006, 57, 450–460. [Google Scholar] [CrossRef]
- Duran, N.; Ozer, B.; Duran, G.G.; Onlen, Y.; Demir, C. Antibiotic resistance genes & susceptibility patterns in staphylococci. Indian J. Med. Res. 2012, 135, 389–396. [Google Scholar]
- Pass, M.A.; Odedra, R.; Batt, R.M. Multiplex PCRs for identification of Escherichia coli virulence genes. J. Clin. Microbiol. 2000, 38, 2001–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, M.; Bakhshi, B.; Peerayeh, S.N.; Behmanesh, M. Detection of curli biogenesis genes among Enterobacter cloacae isolated from blood cultures. Int. J. Enteric Pathog. 2015, 3, e28413. [Google Scholar] [CrossRef] [Green Version]
- Sandner, L.; Eguiarte, L.E.; Navarro, A.; Cravioto, A.; Souza, V. The elements of the locus of enterocyte effacement in human and wild mammal isolates of Escherichia coli: Evolution by assemblage or disruption? Microbiology 2001, 147, 3149–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aher, T.; Roy, A.; Kumar, P. Molecular detection of virulence genes associated with pathogenicity of Klebsiella spp. isolated from the respiratory tract of apparently healthy as well as sick goats. Isr. J. Vet. Med. 2012, 67, 249–252. [Google Scholar]
- Gomes, A.É.I.; Stuchi, L.P.; Siqueira, N.M.G.; Henrique, J.B.; Vicentini, R.; Ribeiro, M.L.; Darrieux, M.; Ferraz, L.F.C. Selection and validation of reference genes for gene expression studies in Klebsiella pneumoniae using reverse transcription quantitative real-time PCR. Sci. Rep. 2018, 8, 9001. [Google Scholar] [CrossRef] [PubMed]
- Matei, A.; Matei, S.; Matei, G.M.; Cogălniceanu, G.; Cornea, C.P. Biosynthesis of silver nanoparticles mediated by culture filtrate of lactic acid bacteria, characterization and antifungal activity. EuroBiotech J. 2020, 4, 97–103. [Google Scholar] [CrossRef]
- Chang, C.C.; Chen, C.P.; Wu, T.H.; Yang, C.H.; Lin, C.W.; Chen, C.Y. Gold nanoparticle-based colorimetric strategies for chemical and biological sensing applications. Nanomaterials 2019, 9, 861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson- Liljequist, B.; et al. Multidrig-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Karbasdehi, V.N.; Dobaradaran, S.; Nabipour, I.; Ostovar, A.; Arfaeinia, H.; Vazirizadeh, A.; Mirahmadi, R.; Keshtkar, M.; Ghasemi, F.F.; Khalifeim, F. Indicator bacteria community in seawater and coastal sediment: The Persian Gulf as a case. J. Environ. Health Sci. Eng. 2017, 15, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valverde, A.; Igual, J.M.; Peix, A.; Cervantes, E.; Velázquez, E. Rhizobium lusitanum sp. nov. a bacterium that nodulates Phaseolus vulgaris. Int. J. Syst. Evol. Microbiol. 2006, 56, 2631–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, D.; Grare, M.; Prere, M.F.; Segonds, C.; Marty, N.; Oswald, E. Performances of the Vitek MS matrix-assisted laser desorption ionization-time of flight mass spectrometry system for rapid identification of bacteria in routine clinical microbiology. J. Clin. Microbiol. 2012, 50, 2568–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Siu, G.K.H.; Yeung, A.S.F.; Chen, J.H.K.; Ho, P.L.; Leung, K.W.; Tsang, J.L.Y.; Cheng, V.C.C.; Guo, L.; Yang, J.; et al. Performance of the VITEK MS matrix-assisted laser desorption ionization-time of flight mass spectrometry system for rapid bacterial identification in two diagnostic centres in China. J. Med. Microbiol. 2015, 64, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibi, S.; Ferjani, A.; Boukadida, J.; de Alegría, C.R.; Martínez-Martínez, L.; Navas, J. Evaluation of the VITEK-MS matrix-assisted laser desorption/ionization time-of-flight mass spectrometry system for the identification of clinical Corynebacterium species. Rev. Esp. Quimioter. 2017, 30, 57–58. [Google Scholar]
- Maravic, A.; Skocibusic, M.; Cvjetan, S.; Samanic, I.; Fredotovic, Z.; Puizina, J. Prevalence and diversity of extended-spectrum-β-lactamase-producing Enterobacteriaceae from marine beach waters. Mar. Pollut. Bull. 2015, 90, 60–67. [Google Scholar] [CrossRef]
- Zango, U.U.; Ibrahim, M.; Shawai, S.A.A.; Shamsuddin, I.M. A review on β-lactam antibiotic drug resistance. MOJ Drug Des. Develop. Ther. 2019, 3, 52–58. [Google Scholar]
- Pongsilp, N.; Nimnoi, P. Diversity and antibiotic resistance patterns of enterobacteria isolated from seafood in Thailand. CyTA J. Food. 2018, 16, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, K.D.; Wang, Y.; Musicha, P.; Hansen, E.G.; Palmer, M.B.; Herman, D.J.; Feasy, N.A.; White, A.P. Parallel evolution leading to impaired biofilm formation in invasive Salmonella strains. PLoS Genet. 2019, 15, e1008233. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.M.; Hassman, H.M.; Orabi, A.; Abdelhafez, A.S.T. Phenotypic, antimicrobial susceptibility profile and virulence factors of Klebsiella pneumoniae isolated from buffalo and cow mastitic milk. Pathog. Glob. Health 2014, 108, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Tomar, R.S.; Agarwal, M.; Jyoti, A. Determination of drug resistance and virulent gene signatures in potable water isolates of Escherichia coli in Gwalior city. J. Pharm. Sci. Res. 2015, 7, 967–971. [Google Scholar]
- Cornick, N.A.; Booher, S.L.; Moon, H.W. Intimin facilitates colonization by Escherichia coli O157-H7 in adult ruminants. Infect. Immun. 2002, 70, 2704–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutin, L.; Krause, G.; Zimmermann, S.; Kaulfuss, S.; Gleier, K. Characterization of Shiga toxin-producing Escherichia coli strains isolated from human patients in Germany over a 3-year period. J. Clin. Microbiol. 2004, 42, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Karama, M.; Mainga, A.O.; Cenci-Goga, B.T.; Malahlela, M.; El-Ashram, S.; Kalake, A. Molecular profiling and antimicrobial resistance of Shiga toxin-producing Escherichia coli O26, O45, O103, O121, O145 and O157 isolates from cattle on cow-calf operations in South Africa. Sci. Rep. 2019, 9, 11930. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Chuang, Y.C.; Yu, W.L.; Lee, N.Y.; Chang, C.M.; Ko, N.Y.; Wang, L.R.; Ko, W.C. Clinical implications of hypermucoviscosity phenotype in Klebsiella pneumoniae isolates: Association with invasive syndrome in patients with community-acquired bacteraemia. J. Intern. Med. 2006, 259, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.J.; Wang, J.T.; Callegan, M.C. Contribution of mucoviscosity-associated gene A (magA) to virulence in experimental Klebsiella pneumoniae endophthalmitis. Invest. Ophthalmol. Vis. Sci. 2011, 52, 6860–6866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdanpour, Z.; Tadjrobehkar, O.; Shahkhah, M. Significant association between genes encoding virulence factors with antibiotic resistance and phylogenetic groups in community acquired uropathogenic Escherichia coli isolates. BMC Microbiol. 2020, 20, 241. [Google Scholar] [CrossRef]
- Haq, K.U.; Noreen, S.; Sehgal, S.A.; Tahir, R.A.; Essa, A.; Ihsan, A.; Hassan, M.U.; Iqbal, W.; Hussain, M.; Hussain, H.; et al. Association of virulence genes with antibiotic resistance in Pakistani uropathogenic E. coli isolates. Comb. Chem. High Throughput Screen 2020, 23, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Baky, R.M.; Ibrahim, R.A.; Mohamed, D.S.; Ahmed, E.F.; Hashem, Z.S. Prevalence of virulence genes and their association with antimicrobial resistance among pathogenic E. coli isolated from Egyptian patients with different clinical infections. Infect. Drug Resist. 2020, 13, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Mukherjee, N.; Pal, C.; Sulaiman, I.M.; Jiang, Y.; Hanna, S.; Dunn, J.R.; Katmaus, W.; Banerjee, P. Association of virulence and antibiotic resistance in Salmonella-statistical and computational insights into a selected set of clinical isolates. Microorganisms 2020, 8, 1465. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef]
- Remya, P.A.; Shanthi, M.; Sekar, U. Characterisation of virulence genes associated with pathogenicity in Klebsiella pneumoniae. Indian J. Med. Microbiol. 2019, 37, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Aljanaby, A. Role of rmpA, wabG, uge, Ycfm, fimh1, EntB, Ybt-irp2 and kfu genes in pathogenicity of Klebsiella pneumoniae: An overview. Int. J. Chemtech. Res. 2017, 10, 391–398. [Google Scholar]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef]
- Rivera, L.; Lopez-Patino, M.A.; Milton, D.L.; Nieto, T.P.; Farto, R. Effective qPCR methodology to quantify the expression of virulence genes in Aeromonas salmonicida subsp. salmonicida. J. Appl. Microbiol. 2015, 118, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.S.; Wigneshweraraj, S. Regulation of virulence gene expression. Virulence 2014, 5, 832–834. [Google Scholar] [CrossRef] [PubMed]
- Chakroun, I.; Mahdhi, A.; Morcillo, P.; Cordero, H.; Cuesta, A.; Bakhrouf, A.; Mahdouani, K.; Esteban, M.A. Motility, biofilm formation, apoptotic effect and virulence gene expression of atypical Salmonella Typhimurium outside and inside Caco-2 cells. Microb. Pathog. 2018, 114, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Liu, Z.G.; Shen, W.; Gurunathan, S. Silver nanoparticles: Synthesis, characterization, properties, applications, and therapeutic approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhang, Y.; Ding, T.; Liu, J.; Zhao, H. Multifunctional gold nanoparticles: A novel nanomaterial for various medical applications and biological activities. Front. Bioeng. Biotechnol. 2020, 8, 990. [Google Scholar] [CrossRef] [PubMed]
- Liebana, E.; Carattoli, A.; Coque, T.M.; Hasman, H.; Magiorakos, A.P.; Mevius, D.; Peixe, L.; Poirel, L.; Schuepbach-Regula, G.; Torneke, K.; et al. Public health risks of enterobacterial isolates producing extended-spectrum β-lactamases or AmpC β-lactamases in food and food-producing animals: An EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 2013, 56, 1030–1037. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Site | Sampling Date | Place | District, Province | Latitude | Longitude | Distance (km) | Land Use |
|---|---|---|---|---|---|---|---|
| A | 11 October 2018 | Black Sand Beach | Laemngop, Trat | 12.169° N | 102.406° E | 0.00 | Mangrove forest |
| B | 11 October 2018 | Kungkrabaen Bay | Thamai, Chanthaburi | 12.573° N | 101.902° E | 123.57 | Mangrove forest |
| C | 11 October 2018 | Suanson Beach | Mueang, Rayong | 12.458° N | 101.473° E | 190.51 | Tourist site |
| D | 11 October 2018 | Pattaya Beach | Banglamoong, Chonburi | 12.936° N | 100.883° E | 326.22 | Tourist site |
| E | 11 October 2018 | Angsila old market | Mueang, Chonburi | 13.341° N | 100.926° E | 388.60 | Aquaculture site |
| F | 12 January 2018 | Donhoylhod | Mueang, Samutsongkhram | 13.361° N | 100.022° E | 527.64 | Aquaculture site |
| G | 12 January 2018 | Bangtaboon Bay | Banlaem, Phetchaburi | 13.264° N | 99.945° E | 548.77 | Aquaculture site |
| H | 12 January 2018 | Pranburi forest park | Pranburi, Prachuapkhirikhan | 12.412° N | 99.981° E | 659.23 | Mangrove forest |
| I | 12 January 2018 | Wanakorn Beach | Thubsakae, Prachuapkhirikhan | 11.635° N | 99.703° E | 769.97 | Tourist site |
| Gene | Nucleotide Sequence of Primer | Size of the PCR Product (bp) |
|---|---|---|
| kfu | KFU-RTf 5′ CGA CCG GTT TCT GGG CGT TA 3′ KFU-RTr 5′ GGC GTT TCA AAA CCG GCG AG 3′ | 293 |
| uge | UGE-RTf 5′ CTC TCA ACG GTC CAG TCG GC 3′ UGE-RTr 5′ CCT GTA TGC CGC CAC CAA GA 3′ | 288 |
| recA | RECA-RTf 5′ TTA AAC AGG CCG AAT TCC AG 3′ RECA-RTr 5′ CCG CTT TCT CAA TCA GCT TC 3′ | 99 |
| Site | Average Number of Presumptive Enterobacteria (CFU/mL) * | Number of Total Coliform Bacteria (MPN Index/100 mL) |
|---|---|---|
| A, Black Sand Beach | 6.22 ± 2.68 | 3500 |
| B, Kungkrabaen Bay | 0.44 ± 0.73 | 9.3 |
| C, Suanson Beach | 17.00 ± 3.97 | 540 |
| D, Pattaya Beach | 1.11 ± 1.05 | 1600 |
| E, Angsila old market | 6.22 ± 4.58 | 350 |
| F, Donhoylhod | 12.33 ± 6.80 | 700 |
| G, Bangtaboon Bay | 6.00 ± 1.41 | 920 |
| H, Pranburi forest park | 1.89 ± 1.36 | 700 |
| I, Wanakorn Beach | 0.22 ± 0.44 | 4.5 |
| Genus/Species | Number of Strain | Number of Strain Possessing Antibiotic Gene | ||
|---|---|---|---|---|
| ampC | blaSHV | blaTEM | ||
| E. cloacae | 13 | 4 | 1 | 1 |
| E. hormaechei | 7 | 1 | 1 | 0 |
| E. coli | 16 | 4 | 3 | 6 |
| K. aerogenes | 4 | 1 | 0 | 0 |
| K. pneumoniae | 48 | 31 | 12 | 5 |
| K. variicola | 12 | 7 | 4 | 0 |
| Pantoea sp. | 1 | 0 | 1 | 0 |
| Total | 101 | 48 | 22 | 12 |
| Genus/Species | Number of Strain | Number of Strain Possessing Virulence Gene | |||||
|---|---|---|---|---|---|---|---|
| csgD | eaeA | kfu | LTI | magA | uge | ||
| E. cloacae | 13 | 8 | 2 | 0 | 0 | 0 | 0 |
| E. hormaechei | 7 | 4 | 0 | 0 | 0 | 0 | 1 |
| E. coli | 16 | 9 | 7 | 0 | 1 | 0 | 0 |
| K. aerogenes | 4 | 1 | 0 | 0 | 0 | 0 | 0 |
| K. pneumoniae | 48 | 12 | 7 | 15 | 2 | 11 | 23 |
| K. variicola | 12 | 4 | 1 | 5 | 2 | 2 | 7 |
| Pantoea sp. | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Total | 101 | 38 | 17 | 20 | 6 | 13 | 32 |
| Virulence Gene | Antibiotic Resistance Gene | |||||
|---|---|---|---|---|---|---|
| ampC | blaSHV | blaTEM | ||||
| r(s) | p | r(s) | p | r(s) | p | |
| csgD | −0.203 | 0.042 * | −0.231 | 0.020 * | 0.141 | 0.161 |
| eaeA | −0.004 | 0.967 | −0.109 | 0.277 | 0.244 | 0.014 * |
| kfu | 0.179 | 0.074 | 0.068 | 0.502 | −0.171 | 0.087 |
| LTI | 0.012 | 0.920 | 0.07 | 0.485 | 0.037 | 0.712 |
| magA | 0.285 | 0.004 * | −0.06 | 0.554 | −0.141 | 0.159 |
| uge | 0.332 | 0.001 * | 0.259 | 0.009 * | −0.119 | 0.238 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nimnoi, P.; Pongsilp, N. Identification, Characterization, and Virulence Gene Expression of Marine Enterobacteria in the Upper Gulf of Thailand. Microorganisms 2022, 10, 511. https://doi.org/10.3390/microorganisms10030511
Nimnoi P, Pongsilp N. Identification, Characterization, and Virulence Gene Expression of Marine Enterobacteria in the Upper Gulf of Thailand. Microorganisms. 2022; 10(3):511. https://doi.org/10.3390/microorganisms10030511
Chicago/Turabian StyleNimnoi, Pongrawee, and Neelawan Pongsilp. 2022. "Identification, Characterization, and Virulence Gene Expression of Marine Enterobacteria in the Upper Gulf of Thailand" Microorganisms 10, no. 3: 511. https://doi.org/10.3390/microorganisms10030511