
Easy Removal of Epiphytic Bacteria on Ulva (Ulvophyceae, Chlorophyta) by Vortex with Silica Sands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Samples
2.2. Sterilization and SEM Detection
2.3. Denaturing Gradient Gel Electrophoresis
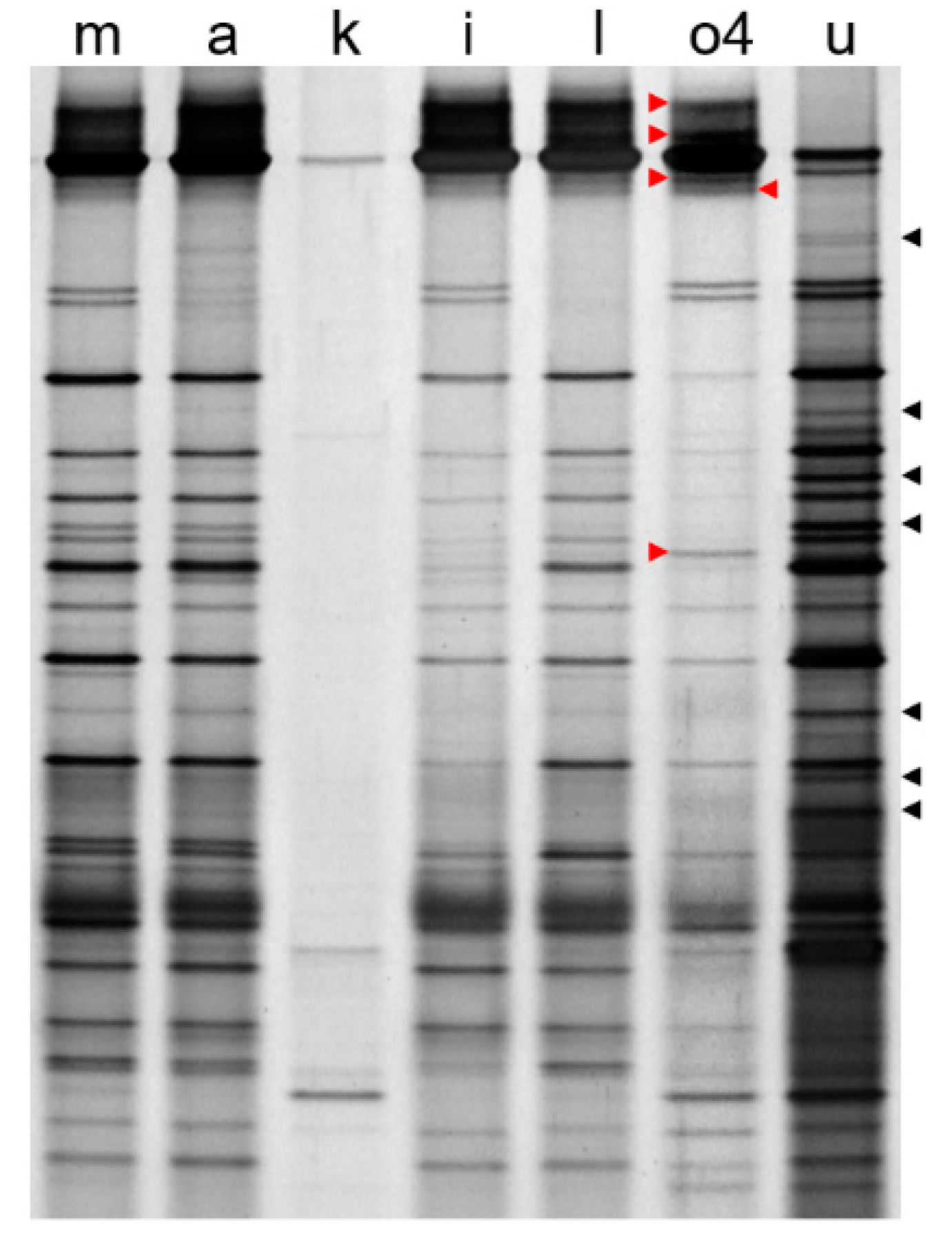
3. Results and Discussions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Chemical interactions between marine macroalgae and bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–299. [Google Scholar] [CrossRef]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin B-12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.M.; Repeta, D.J.; Laroche, J. Dissolved organic nitrogen hydrolysis rates in axenic cultures of Aureococcus anophagefferens (Pelagophyceae): Comparison with heterotrophic bacteria. Appl. Environ. Microbiol. 2002, 68, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobretsov, S.V.; Qian, P.Y. Effect of bacteria associated with the green alga Ulva reticulata on marine micro- and macrofouling. Biofouling 2002, 18, 217–228. [Google Scholar] [CrossRef]
- Ghaderiardakani, F.; Quartino, M.L.; Wichard, T. Microbiome-dependent adaptation of seaweeds under environmental stresses: A perspective. Front. Mar. Sci. 2020, 7, 575228. [Google Scholar] [CrossRef]
- Patel, P.; Callow, M.E.; Joint, I.; Callow, J.A. Specificity in the settlement—modifying response of bacterial biofilms towards zoospores of the marine alga Enteromorpha. Environ. Microbiol. 2003, 5, 338–349. [Google Scholar] [CrossRef]
- Matsuo, Y.; Imagawa, H.; Nishizawa, M.; Shizuri, Y. Isolation of an algal morphogenesis inducer from a marine bacterium. Science 2005, 307, 1598. [Google Scholar] [CrossRef]
- Marshall, K.; Joint, I.; Callow, M.E.; Callow, J.A. Effect of marine bacterial isolates on the growth and morphology of axenic plantlets of the green alga Ulva linza. Microbial. Ecol. 2006, 52, 302–310. [Google Scholar] [CrossRef]
- Spoerner, M.; Wichard, T.; Bachhuber, T.; Stratmann, J.; Oertel, W. Growth and thallus morphogenesis of Ulva mutabilis (Chlorophyta) depends on a combination of two bacterial species excreting regulatory factors. J. Phycol. 2012, 48, 1433–1447. [Google Scholar] [CrossRef]
- Wichard, T. Exploring bacteria-induced growth and morphogenesis in the green macroalga order Ulvales (Chlorophyta). Front. Plant Sci. 2015, 6, 86. [Google Scholar] [CrossRef]
- Wichard, T.; Charrier, B.; Mineur, F.; Bothwell, J.H.; De Clerck, O.; Coates, J.C. The green seaweed Ulva: A model system to study morphogenesis. Front. Plant Sci. 2015, 6, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsufyani, T.; Califano, G.; Deicke, M.; Grueneberg, J.; Weiss, A.; Engelen, A.H.; Kwantes, M.; Mohr, J.F.; Ulrich, J.F.; Wichard, T. Macroalgal–bacterial interactions: Identification and role of thallusin in morphogenesis of the seaweed Ulva (Chlorophyta). J. Exp. Bot. 2020, 71, 3340–3349. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.; Thomas, T.; Holmstrom, C.; Kjelleberg, S. Phylogenetic relationship and antifouling activity of bacterial epiphytes from the marine alga Ulva lactuca. Environ. Microbiol. 2000, 2, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Delbridge, L.; Coulburn, J.; Fagerberg, W.; Tisa, L.S. Community profiles of bacterial endosymbionts in four species of Caulerpa. Symbiosis 2004, 37, 335–344. [Google Scholar]
- Burke, C.; Thomas, T.; Lewis, M.; Steinberg, P.; Kjelleberg, S. Composition, uniqueness and variability of the epiphytic bacterial community of the green alga Ulva australis. ISME J. 2011, 5, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Aires, T.; Serrao, E.A.; Kendrick, G.; Duarte, C.M.; Arnaud-Haond, S. Invasion is a community affair: Clandestine followers in the bacterial community associated to green algae, Caulerpa racemosa, track the invasion source. PLoS ONE 2013, 8, e68429. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.H.; Jiang, P.; Zhao, J.; Wu, C.H. Comparative genomics of Pseudomonas sp. strain SI-3 associated with macroalga Ulva prolifera, the causative species for green tide in the Yellow Sea. Front. Microbiol. 2018, 9, 1458. [Google Scholar] [CrossRef]
- Mei, X.Y.; Wu, C.H.; Zhao, J.; Yan, T.; Jiang, P. Community structure of bacteria associated with drifting Sargassum horneri, the causative species of golden tide in the Yellow Sea. Front. Microbiol. 2019, 10, 1192. [Google Scholar] [CrossRef] [Green Version]
- Burke, C.; Kjelleberg, S.; Thomas, T. Selective extraction of bacterial DNA from the surfaces of macroalgae. Appl. Environ. Microbiol. 2009, 75, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.M.; Wilcox, L.W.; Gram, L.E. Molecular characterization of epiphytic bacterial communities on Charophycean green algae. Appl. Environ. Microbiol. 1998, 64, 4384–4389. [Google Scholar] [CrossRef] [Green Version]
- Hollants, J.; Leliaert, F.; De Clerck, O.; Willems, A. How endo- is endo-? Surface sterilization of delicate samples: A Bryopsis (Bryopsidales, Chlorophyta) case study. Symbiosis 2010, 51, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Aires, T.; Marba, N.; Serrao, E.A.; Duarte, C.M.; Arnaud-Haond, S. Selective elimination of chloroplastidial DNA for metagenomics of bacteria associated with the green alga Caulerpa taxifolia (Bryopsidophyceae). J. Phycol. 2012, 48, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kientz, B.; Thabard, M.; Cragg, S.M.; Pope, J.; Hellio, C. A new method for removing microflora from macroalgal surfaces: An important step for natural product discovery. Bot. Mar. 2011, 54, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.X.; Geng, Z.L.; Zeng, Y.; Shen, Y.M. Enriching plant microbiota for a metagenomic library construction. Environ. Microbiol. 2008, 10, 2684–2691. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.L. The occurrence of ‘‘green tides’’—A review. In Marine Benthic Vegetation: Recent Changes and the Effects of Eutrophication; Schramm, W., Nienhuis, P.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 7–43. [Google Scholar]
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Blomster, J.; Bäck, S.; Fewer, D.P.; Kiirikki, M.; Lehvo, A.; Maggs, C.A.; Stanhope, M. Novel morphology in Enteromorpha (Ulvophyceae) forming green tides. Am. J. Bot. 2002, 89, 1756–1763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Huo, Y.Z.; Zhang, Z.L.; Yu, K.F.; He, Q.; Zhang, L.H.; Yang, L.L.; Xu, R.; He, P.M. Variations of morphology and photosynthetic performances of Ulva prolifera during the whole green tide blooming process in the Yellow Sea. Mar. Environ. Res. 2013, 92, 35–42. [Google Scholar] [CrossRef]
- Gao, G.; Zhong, Z.; Zhou, X.; Xu, J. Changes in morphological plasticity of Ulva prolifera under different environmental conditions: A laboratory experiment. Harmful Algae 2016, 59, 51–58. [Google Scholar] [CrossRef]
- Qu, T.; Hou, C.; Zhao, X.; Zhong, Y.; Guan, C.; Lin, Z.; Tang, X.; Wang, Y. Bacteria associated with Ulva prolifera: A vital role in green tide formation and migration. Harmful Algae 2021, 108, 102104. [Google Scholar] [CrossRef]
- Zhao, J.; Jiang, P.; Liu, Z.Y.; Wei, W.; Lin, H.Z.; Li, F.C.; Wang, J.F.; Qin, S. The Yellow Sea green tides were dominated by one species, Ulva (Enteromorpha) prolifera, from 2007 to 2011. Chin. Sci. Bull. 2013, 58, 2298–2302. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Jiang, P.; Qin, S.; Liu, X.J.; Liu, Z.Y.; Lin, H.Z.; Li, F.C.; Chen, H.X.; Wu, C.H. Genetic analyses of floating Ulva prolifera in the Yellow Sea suggest a unique ecotype. Estuar. Coast. Shelf Sci. 2015, 163, 96–102. [Google Scholar] [CrossRef]
- Callow, M.E.; Evans, L.V.; Bolwell, G.P.; Callow, J.A. Fertilization in brown-algae. 1. SEM and other observations on Fucus serratus. J. Cell Sci. 1978, 32, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Muyzer, G.; Dewaal, E.C.; Uitterlinden, A.G. Profiling of complex microbial-populations by denaturing gradient gel-electrophoresis analysis of polymerase chain reaction-amplified genes-coding for 16S ribosomal-RNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, J.R.; Swerhone, G.D.W.; Leppard, G.G.; Araki, T.; Zhang, X.; West, M.M.; Hitchcock, A.P. Scanning transmission X-ray, laser scanning, and transmission electron microscopy mapping of the exopolymeric matrix of microbial biofilms. Appl. Environ. Microbiol. 2003, 69, 5543–5554. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Reddy, C.R.K. Seaweed-microbial interactions: Key functions of seaweed-associated bacteria. FEMS Microbiol. Ecol. 2014, 88, 213–230. [Google Scholar] [CrossRef]
- Barreto, M.; Meyer, J.J.M. The preservation of biofilms on macroalgae by osmium vapour. S. Afr. J. Bot. 2007, 73, 64–69. [Google Scholar] [CrossRef] [Green Version]
- de Beer, D.; Srinivasan, R.; Stewart, P.S. Direct measurement of chlorine penetration into biofilms during disinfection. Appl. Environ. Microbiol. 1994, 60, 4339–4344. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Wiese, J.; Weinberger, F.; Wahl, M. Rapid adaptation to controlling new microbial epibionts in the invaded range promotes invasiveness of an exotic seaweed. J. Ecol. 2016, 104, 969–978. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Type | Group No. | Sterilization Method | Extended Protocol |
|---|---|---|---|
| Control | a | no | fresh S1, control for groups from c to o4 |
| b | no | frozen S1, control for group o5 | |
| Enzymatic | c | lysozyme (AMRESCO, Solon, OH, USA) | S1 + 2 mg/mL lysozyme in SSW 1, 37 °C × 1 h |
| d | papain | S1 + 2 mg/mL papain in SSW, 55 °C × 30 min | |
| e | snailase | S1 + 2 mg/mL snailase in SSW, 37 °C × 1 h | |
| f | cellulase | S1 + 2 mg/mL cellulase in SSW, 25 °C × 2 h | |
| g | lysozyme + papain + snailase | S1 + mixture of lysozyme, papain, and snailase (2 mg/mL each) in SSW, 37 °C × 1 h | |
| h | rapid multienzyme cleaner | S1 + EDTA and filter-sterilized rapid multienzyme cleaner (3M, North Ryde, NSW, Australia) in CFASW 2, room temperature × 2 h | |
| i | proteinase K | S1 + 2 mg/mL proteinase K in SSW, 60 °C × 30 min | |
| Chemical | j | ethanol | S1 + 75% ethanol, room temperature × 5 min |
| k | bleach | S1 + 3% sodium hypochlorite, room temperature × 30 s | |
| l | UNSET buffer | S1 + 1 mL UNSET buffer, 55 °C × 15 min 3 | |
| Mechanical | m | ultrasonic bath sonication | S1 in ultrasonic cleaner, 40 kHz × 15 min |
| n | glass beads (Bio-Rad, Hercules, CA, USA)-vortex | 0.03 g S1 + 0.3 g glass beads (120–250 µm) in SSW, vortex in 1.5 mL tube at 3200 rpm, 2 × 20 min | |
| o1 | silica sand-vortex | 0.03 g S1 + 0.3 g silica sand (60–125 µm) in SSW, vortex in 1.5 mL tube at 3200 rpm for 2 × 1 h | |
| o2 | silica sand-vortex | same as group o1 except for size of silica sand (125–250 µm) | |
| o3 | silica sand-vortex | same as group o2 except for 2 × 30 min | |
| o4 | silica sand-vortex | same as group o2 except for 2 × 15 min | |
| o5 | silica sand-vortex | same as group o4 except for using frozen S1 | |
| p | silica sand-vortex | same as group o4 except for using frozen S3 | |
| q | silica sand-vortex | same as group o4 except for using S2 | |
| r | silica sand-vortex | same as group o4 except for using U. compressa | |
| s | silica sand-vortex | same as group o4 except for using U. linza |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhao, J.; Jiang, P. Easy Removal of Epiphytic Bacteria on Ulva (Ulvophyceae, Chlorophyta) by Vortex with Silica Sands. Microorganisms 2022, 10, 476. https://doi.org/10.3390/microorganisms10020476
Liu X, Zhao J, Jiang P. Easy Removal of Epiphytic Bacteria on Ulva (Ulvophyceae, Chlorophyta) by Vortex with Silica Sands. Microorganisms. 2022; 10(2):476. https://doi.org/10.3390/microorganisms10020476
Chicago/Turabian StyleLiu, Xiaojie, Jin Zhao, and Peng Jiang. 2022. "Easy Removal of Epiphytic Bacteria on Ulva (Ulvophyceae, Chlorophyta) by Vortex with Silica Sands" Microorganisms 10, no. 2: 476. https://doi.org/10.3390/microorganisms10020476