
Bacterial Biofilms Utilize an Underlying Extracellular DNA Matrix Structure That Can Be Targeted for Biofilm Resolution


{kind=link}
Abstract
:1. Introduction
2. eDNA
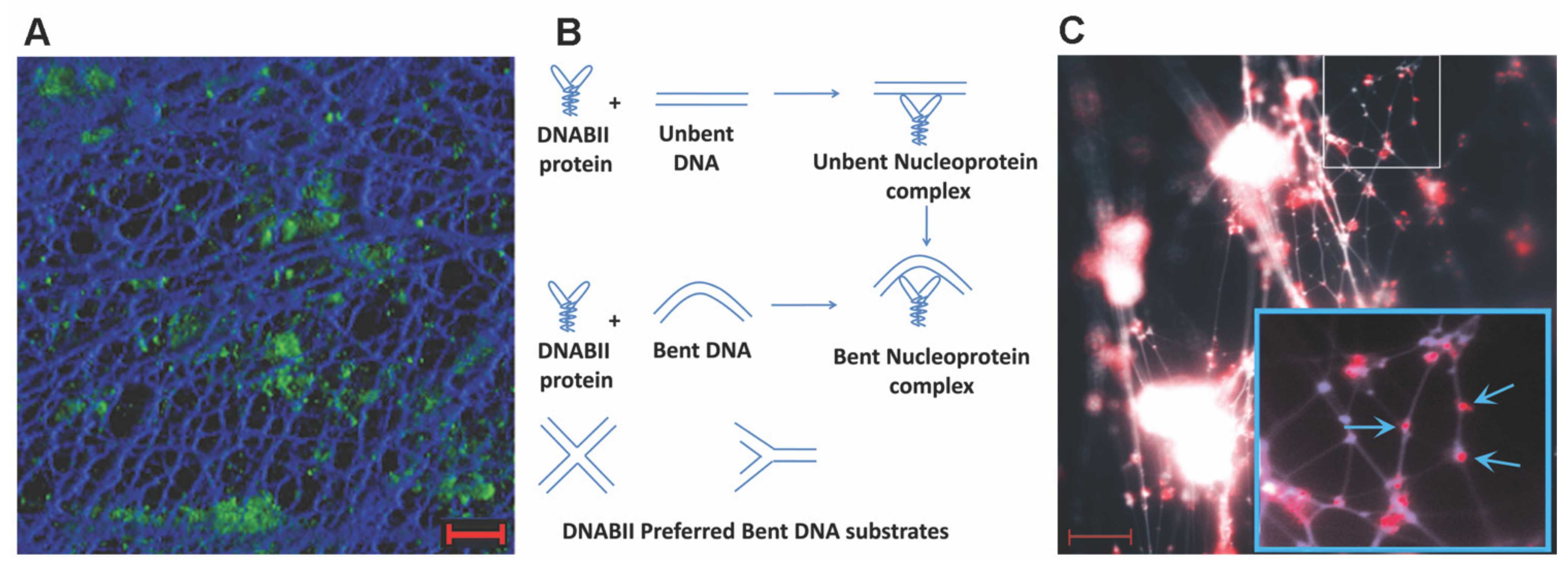
3. DNABII Proteins
4. Mechanisms of eDNA and DNABII Release
5. Immunological Depletion of DNABII Disrupts Biofilms In Vitro
6. Immunological Depletion of DNABII Disrupts Biofilms In Vivo: Clinical Relevance and Pre-Clinical Data
7. eDNA-Dependent Structure Relies on HJs
8. eDNA Is Important throughout Biofilm Development but Recalcitrant to Degradation by Various Nucleases
9. The Nature of the Nuclease-Resistant Structure of Biofilm eDNA
10. Inclusivity vs. Exclusivity and Future Work
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall-Stoodley, L.; Stoodley, P.; Kathju, S.; Hoiby, N.; Moser, C.; Costerton, J.W.; Moter, A.; Bjarnsholt, T. Towards diagnostic guidelines for biofilm-associated infections. FEMS Immunol Med. Microbiol. 2012, 65, 127–145. [Google Scholar] [CrossRef] [Green Version]
- Stewart, P.S. Biophysics of biofilm infection. Pathog. Dis. 2014, 70, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Stinson, M.W.; Bergey, E.J. Isolation of heart- and kidney-binding protein from group A streptococci. Infect. Immun. 1982, 35, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Winters, B.D.; Ramasubbu, N.; Stinson, M.W. Isolation and characterization of a Streptococcus pyogenes protein that binds to basal laminae of human cardiac muscle. Infect. Immun. 1993, 61, 3259–3264. [Google Scholar] [CrossRef] [Green Version]
- Shimoji, Y.; Ng, V.; Matsumura, K.; Fischetti, V.A.; Rambukkana, A. A 21-kDa surface protein of Mycobacterium leprae binds peripheral nerve laminin-2 and mediates Schwann cell invasion. Proc. Natl Acad Sci USA 1999, 96, 9857–9862. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Weeks, D.L.; Shin, J.M.; Scott, D.R.; Young, M.K.; Sachs, G. Proteins released by Helicobacter pylori in vitro. J. Bacteriol. 2002, 184, 6155–6162. [Google Scholar] [CrossRef] [Green Version]
- Boleij, A.; Schaeps, R.M.; de Kleijn, S.; Hermans, P.W.; Glaser, P.; Pancholi, V.; Swinkels, D.W.; Tjalsma, H. Surface-exposed histone-like protein a modulates adherence of Streptococcus gallolyticus to colon adenocarcinoma cells. Infect. Immun. 2009, 77, 5519–5527. [Google Scholar] [CrossRef] [Green Version]
- Goodman, S.D.; Obergfell, K.P.; Jurcisek, J.A.; Novotny, L.A.; Downey, J.S.; Ayala, E.A.; Tjokro, N.; Li, B.; Justice, S.S.; Bakaletz, L.O. Biofilms can be dispersed by focusing the immune system on a common family of bacterial nucleoid-associated proteins. Mucosal Immunol. 2011, 4, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Nur, A.; Hirota, K.; Yumoto, H.; Hirao, K.; Liu, D.; Takahashi, K.; Murakami, K.; Matsuo, T.; Shu, R.; Miyake, Y. Effects of extracellular DNA and DNA-binding protein on the development of a Streptococcus intermedius biofilm. J. Appl. Microbiol. 2013, 115, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Dey, D.; Nagaraja, V.; Ramakumar, S. Structural and evolutionary analyses reveal determinants of DNA binding specificities of nucleoid-associated proteins HU and IHF. Mol. Phylogenet. Evol. 2017, 107, 356–366. [Google Scholar] [CrossRef]
- Avery, O.T.; Macleod, C.M.; McCarty, M. Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types: Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated from Pneumococcus Type Iii. J. Exp. Med. 1944, 79, 137–158. [Google Scholar] [CrossRef]
- Li, Y.H.; Lau, P.C.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. Natural genetic transformation of Streptococcus mutans growing in biofilms. J. Bacteriol. 2001, 183, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Jurcisek, J.A.; Brockman, K.L.; Novotny, L.A.; Goodman, S.D.; Bakaletz, L.O. Nontypeable Haemophilus influenzae releases DNA and DNABII proteins via a T4SS-like complex and ComE of the type IV pilus machinery. Proc. Natl Acad Sci USA 2017, 114, E6632–E6641. [Google Scholar] [CrossRef] [Green Version]
- Smithies, W.R.; Gibbons, N.E. The deoxyribose nucleic acid slime layer of some halophilic bacteria. Can. J. Microbiol. 1955, 1, 614–621. [Google Scholar] [CrossRef]
- Murakawa, T. Slime production by Pseudomonas aeruginosa. IV. Chemical analysis of two varieties of slime produced by Pseudomonas aeruginosa. Jpn. J. Microbiol. 1973, 17, 513–520. [Google Scholar] [CrossRef]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Jurcisek, J.A.; Bakaletz, L.O. Biofilms formed by nontypeable Haemophilus influenzae in vivo contain both double-stranded DNA and type IV pilin protein. J. Bacteriol. 2007, 189, 3868–3875. [Google Scholar] [CrossRef] [Green Version]
- Bockelmann, U.; Janke, A.; Kuhn, R.; Neu, T.R.; Wecke, J.; Lawrence, J.R.; Szewzyk, U. Bacterial extracellular DNA forming a defined network-like structure. FEMS Microbiol. Lett. 2006, 262, 31–38. [Google Scholar] [CrossRef]
- Swinger, K.K.; Rice, P.A. IHF and HU: Flexible architects of bent DNA. Curr. Opin. Struct. Biol. 2004, 14, 28–35. [Google Scholar] [CrossRef]
- Pontiggia, A.; Negri, A.; Beltrame, M.; Bianchi, M.E. Protein HU binds specifically to kinked DNA. Mol. Microbiol. 1993, 7, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Bonnefoy, E.; Takahashi, M.; Yaniv, J.R. DNA-binding parameters of the HU protein of Escherichia coli to cruciform DNA. J. Mol. Biol. 1994, 242, 116–129. [Google Scholar] [CrossRef]
- Kamashev, D.; Balandina, A.; Rouviere-Yaniv, J. The binding motif recognized by HU on both nicked and cruciform DNA. EMBO J. 1999, 18, 5434–5444. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, A.; Novotny, L.A.; Robledo-Avila, F.H.; Buzzo, J.R.; Mashburn-Warren, L.; Jurcisek, J.A.; Tjokro, N.O.; Partida-Sanchez, S.; Bakaletz, L.O.; Goodman, S.D. The extracellular innate-immune effector HMGB1 limits pathogenic bacterial biofilm proliferation. J. Clin. Investig. 2021, 131, e140527. [Google Scholar] [CrossRef]
- Justice, S.S.; Li, B.; Downey, J.S.; Dabdoub, S.M.; Brockson, M.E.; Probst, G.D.; Ray, W.C.; Goodman, S.D. Aberrant community architecture and attenuated persistence of uropathogenic Escherichia coli in the absence of individual IHF subunits. PLoS ONE 2012, 7, e48349. [Google Scholar] [CrossRef] [Green Version]
- Gustave, J.E.; Jurcisek, J.A.; McCoy, K.S.; Goodman, S.D.; Bakaletz, L.O. Targeting bacterial integration host factor to disrupt biofilms associated with cystic fibrosis. J. Cyst. Fibros. 2013, 12, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Novotny, L.A.; Amer, A.O.; Brockson, M.E.; Goodman, S.D.; Bakaletz, L.O. Structural stability of Burkholderia cenocepacia biofilms is reliant on eDNA structure and presence of a bacterial nucleic acid binding protein. PLoS ONE 2013, 8, e67629. [Google Scholar] [CrossRef]
- Idicula, W.K.; Jurcisek, J.A.; Cass, N.D.; Ali, S.; Goodman, S.D.; Elmaraghy, C.A.; Jatana, K.R.; Bakaletz, L.O. Identification of biofilms in post-tympanostomy tube otorrhea. Laryngoscope 2016, 126, 1946–1951. [Google Scholar] [CrossRef]
- Devaraj, A.; Buzzo, J.; Rocco, C.J.; Bakaletz, L.O.; Goodman, S.D. The DNABII family of proteins is comprised of the only nucleoid associated proteins required for nontypeable Haemophilus influenzae biofilm structure. Microbiologyopen 2018, 7, e00563. [Google Scholar] [CrossRef] [Green Version]
- Claverys, J.P.; Martin, B.; Havarstein, L.S. Competence-induced fratricide in streptococci. Mol. Microbiol. 2007, 64, 1423–1433. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Bakaletz, L.O.; Wozniak, D.J. What’s on the Outside Matters: The Role of the Extracellular Polymeric Substance of Gram-negative Biofilms in Evading Host Immunity and as a Target for Therapeutic Intervention. J. Biol. Chem. 2016, 291, 12538–12546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, J.; Mokrzan, E.; Lakhani, V.; Rosas, L.; Jurcisek, J.A.; Ray, W.C.; Bakaletz, L.O. Extracellular DNA and Type IV Pilus Expression Regulate the Structure and Kinetics of Biofilm Formation by Nontypeable Haemophilus influenzae. mBio 2017, 8, e01466-17. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.; Jurcisek, J.; Lakhani, V.; Snedden, A.; Ray, W.C.; Mokrzan, E.M.; Bakaletz, L.O.; Das, J. In Silico Modeling of Biofilm Formation by Nontypeable Haemophilus influenzae In Vivo. mSphere 2019, 4, e00254-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaraj, A.; Buzzo, J.R.; Mashburn-Warren, L.; Gloag, E.S.; Novotny, L.A.; Stoodley, P.; Bakaletz, L.O.; Goodman, S.D. The extracellular DNA lattice of bacterial biofilms is structurally related to Holliday junction recombination intermediates. Proc. Natl Acad Sci USA 2019, 116, 25068–25077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockson, M.E.; Novotny, L.A.; Mokrzan, E.M.; Malhotra, S.; Jurcisek, J.A.; Akbar, R.; Devaraj, A.; Goodman, S.D.; Bakaletz, L.O. Evaluation of the kinetics and mechanism of action of anti-integration host factor-mediated disruption of bacterial biofilms. Mol. Microbiol. 2014, 93, 1246–1258. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, A.; Justice, S.S.; Bakaletz, L.O.; Goodman, S.D. DNABII proteins play a central role in UPEC biofilm structure. Mol. Microbiol. 2015, 96, 1119–1135. [Google Scholar] [CrossRef] [Green Version]
- Rocco, C.J.; Davey, M.E.; Bakaletz, L.O.; Goodman, S.D. Natural antigenic differences in the functionally equivalent extracellular DNABII proteins of bacterial biofilms provide a means for targeted biofilm therapeutics. Mol. Oral Microbiol. 2017, 32, 118–130. [Google Scholar] [CrossRef]
- Rocco, C.J.; Bakaletz, L.O.; Goodman, S.D. Targeting the HUbeta Protein Prevents Porphyromonas gingivalis from Entering into Preexisting Biofilms. J. Bacteriol. 2018, 200, e00790-17. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, A.; Gonzalez, J.F.; Eichar, B.; Thilliez, G.; Kingsley, R.A.; Baker, S.; Allard, M.W.; Bakaletz, L.O.; Gunn, J.S.; Goodman, S.D. Enhanced biofilm and extracellular matrix production by chronic carriage versus acute isolates of Salmonella Typhi. PLoS Pathog. 2021, 17, e1009209. [Google Scholar] [CrossRef]
- Mokrzan, E.M.; Novotny, L.A.; Brockman, K.L.; Bakaletz, L.O. Antibodies against the Majority Subunit (PilA) of the Type IV Pilus of Nontypeable Haemophilus influenzae Disperse Moraxella catarrhalis from a Dual-Species Biofilm. mBio 2018, 9, e02423-18. [Google Scholar] [CrossRef] [Green Version]
- Mokrzan, E.M.; Ahearn, C.P.; Buzzo, J.R.; Novotny, L.A.; Zhang, Y.; Goodman, S.D.; Bakaletz, L.O. Nontypeable Haemophilus influenzae newly released (NRel) from biofilms by antibody-mediated dispersal versus antibody-mediated disruption are phenotypically distinct. Biofilm 2020, 2, 100039. [Google Scholar] [CrossRef]
- Sauer, K.; Cullen, M.C.; Rickard, A.H.; Zeef, L.A.; Davies, D.G.; Gilbert, P. Characterization of nutrient-induced dispersion in Pseudomonas aeruginosa PAO1 biofilm. J. Bacteriol. 2004, 186, 7312–7326. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.R.; Davidson, B.A.; Knight, P.R.; Hakansson, A.P. Interkingdom signaling induces Streptococcus pneumoniae biofilm dispersion and transition from asymptomatic colonization to disease. mBio 2013, 4, e00438-13. [Google Scholar] [CrossRef] [Green Version]
- Chua, S.L.; Liu, Y.; Yam, J.K.; Chen, Y.; Vejborg, R.M.; Tan, B.G.; Kjelleberg, S.; Tolker-Nielsen, T.; Givskov, M.; Yang, L. Dispersed cells represent a distinct stage in the transition from bacterial biofilm to planktonic lifestyles. Nat. Commun. 2014, 5, 4462. [Google Scholar] [CrossRef]
- Chambers, J.R.; Cherny, K.E.; Sauer, K. Susceptibility of Pseudomonas aeruginosa Dispersed Cells to Antimicrobial Agents Is Dependent on the Dispersion Cue and Class of the Antimicrobial Agent Used. Antimicrob. Agents Chemother. 2017, 61, e00846-17. [Google Scholar] [CrossRef] [Green Version]
- Goodwine, J.; Gil, J.; Doiron, A.; Valdes, J.; Solis, M.; Higa, A.; Davis, S.; Sauer, K. Pyruvate-depleting conditions induce biofilm dispersion and enhance the efficacy of antibiotics in killing biofilms in vitro and in vivo. Sci Rep. 2019, 9, 3763. [Google Scholar] [CrossRef] [Green Version]
- Rood, K.M.; Buhimschi, I.A.; Jurcisek, J.A.; Summerfield, T.L.; Zhao, G.; Ackerman, W.E.; Wang, W.; Rumpf, R.W.; Thung, S.F.; Bakaletz, L.O.; et al. Skin Microbiota in Obese Women at Risk for Surgical Site Infection After Cesarean Delivery. Sci. Rep. 2018, 8, 8756. [Google Scholar] [CrossRef]
- Barron, C.L.; Kamel-Abusalha, L.B.; Sethia, R.; Goodman, S.D.; Elmaraghy, C.A.; Bakaletz, L.O. Identification of essential biofilm proteins in middle ear fluids of otitis media with effusion patients. Laryngoscope 2020, 130, 806–811. [Google Scholar] [CrossRef]
- Novotny, L.A.; Jurcisek, J.A.; Goodman, S.D.; Bakaletz, L.O. Monoclonal antibodies against DNA-binding tips of DNABII proteins disrupt biofilms in vitro and induce bacterial clearance in vivo. EBioMedicine 2016, 10, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Brandstetter, K.A.; Jurcisek, J.A.; Goodman, S.D.; Bakaletz, L.O.; Das, S. Antibodies directed against integration host factor mediate biofilm clearance from Nasopore. Laryngoscope 2013, 123, 2626–2632. [Google Scholar] [CrossRef] [Green Version]
- Martyn, L.; Sethia, R.; Chon, R.; Novotny, L.; Goodman, S.D.; Elmaraghy, C.; Bakaletz, L.O. Antibodies against the DNABII protein integration host factor (IHF) inhibit sinus implant biofilms. Laryngoscope 2020, 130, 1364–1371. [Google Scholar] [CrossRef]
- Freire, M.O.; Devaraj, A.; Young, A.; Navarro, J.B.; Downey, J.S.; Chen, C.; Bakaletz, L.O.; Zadeh, H.H.; Goodman, S.D. A bacterial-biofilm-induced oral osteolytic infection can be successfully treated by immuno-targeting an extracellular nucleoid-associated protein. Mol. Oral Microbiol. 2017, 32, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Novotny, L.A.; Goodman, S.D.; Bakaletz, L.O. Redirecting the immune response towards immunoprotective domains of a DNABII protein resolves experimental otitis media. NPJ Vaccines 2019, 4, 43. [Google Scholar] [CrossRef]
- Novotny, L.A.; Clements, J.D.; Goodman, S.D.; Bakaletz, L.O. Transcutaneous Immunization with a Band-Aid Prevents Experimental Otitis Media in a Polymicrobial Model. Clin. Vaccine Immunol. 2017, 24, e00563-16. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.T.; Lauber, C.L.; Novotny, L.A.; Goodman, S.D.; Bakaletz, L.O. Immunization with a Biofilm-Disrupting Nontypeable Haemophilus influenzae Vaccine Antigen Did Not Alter the Gut Microbiome in Chinchillas, Unlike Oral Delivery of a Broad-Spectrum Antibiotic Commonly Used for Otitis Media. mSphere 2020, 5, e00296-20. [Google Scholar] [CrossRef] [Green Version]
- Nijland, R.; Hall, M.J.; Burgess, J.G. Dispersal of biofilms by secreted, matrix degrading, bacterial DNase. PLoS ONE 2010, 5, e15668. [Google Scholar] [CrossRef] [Green Version]
- Shields, R.C.; Mokhtar, N.; Ford, M.; Hall, M.J.; Burgess, J.G.; ElBadawey, M.R.; Jakubovics, N.S. Efficacy of a marine bacterial nuclease against biofilm forming microorganisms isolated from chronic rhinosinusitis. PLoS ONE 2013, 8, e55339. [Google Scholar] [CrossRef] [Green Version]
- Seviour, T.; Winnerdy, F.R.; Wong, L.L.; Shi, X.; Mugunthan, S.; Foo, Y.H.; Castaing, R.; Adav, S.S.; Subramoni, S.; Kohli, G.S.; et al. The biofilm matrix scaffold of Pseudomonas aeruginosa contains G-quadruplex extracellular DNA structures. NPJ Biofilms Microbiomes 2021, 7, 27. [Google Scholar] [CrossRef]
- Zhou, H.; Hintze, B.J.; Kimsey, I.J.; Sathyamoorthy, B.; Yang, S.; Richardson, J.S.; Al-Hashimi, H.M. New insights into Hoogsteen base pairs in DNA duplexes from a structure-based survey. Nucleic Acids Res. 2015, 43, 3420–3433. [Google Scholar] [CrossRef]
- Yau, H.C.L.; Malekpour, A.K.; Momin, N.G.; Morales-Garcia, A.L.; Willats, W.G.T.; Lant, N.J.; Jones, C.Y. Removal of eDNA from fabrics using a novel laundry DNase revealed using high-resolution imaging. Sci. Rep. 2021, 11, 21542. [Google Scholar] [CrossRef]
- Buzzo, J.R.; Devaraj, A.; Gloag, E.S.; Jurcisek, J.A.; Robledo-Avila, F.; Kesler, T.; Wilbanks, K.; Mashburn-Warren, L.; Balu, S.; Wickham, J.; et al. Z-form extracellular DNA is a structural component of the bacterial biofilm matrix. Cell 2021, 184, 5740–5758.e5717. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.B.; Mashburn-Warren, L.; Bakaletz, L.O.; Bailey, M.T.; Goodman, S.D. Enhanced Probiotic Potential of Lactobacillus reuteri When Delivered as a Biofilm on Dextranomer Microspheres That Contain Beneficial Cargo. Front. Microbiol. 2017, 8, 489. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goodman, S.D.; Bakaletz, L.O. Bacterial Biofilms Utilize an Underlying Extracellular DNA Matrix Structure That Can Be Targeted for Biofilm Resolution. Microorganisms 2022, 10, 466. https://doi.org/10.3390/microorganisms10020466
Goodman SD, Bakaletz LO. Bacterial Biofilms Utilize an Underlying Extracellular DNA Matrix Structure That Can Be Targeted for Biofilm Resolution. Microorganisms. 2022; 10(2):466. https://doi.org/10.3390/microorganisms10020466
Chicago/Turabian StyleGoodman, Steven D., and Lauren O. Bakaletz. 2022. "Bacterial Biofilms Utilize an Underlying Extracellular DNA Matrix Structure That Can Be Targeted for Biofilm Resolution" Microorganisms 10, no. 2: 466. https://doi.org/10.3390/microorganisms10020466