
Probiotic Potentiality from Versatile Lactiplantibacillus plantarum Strains as Resource to Enhance Freshwater Fish Health

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. LAB Strains
2.2. Antimicrobial Activity
2.3. Evaluation of Acid and Bile Tolerance
2.4. Cell Surface Properties
2.4.1. Bacterial Cultures
2.4.2. Auto-Aggregation
2.4.3. Hydrophobicity
2.5. Antioxidant Activity
2.6. Statistical Analysis
3. Results
3.1. Antimicrobial Activity
3.2. pH and Bile Resistance
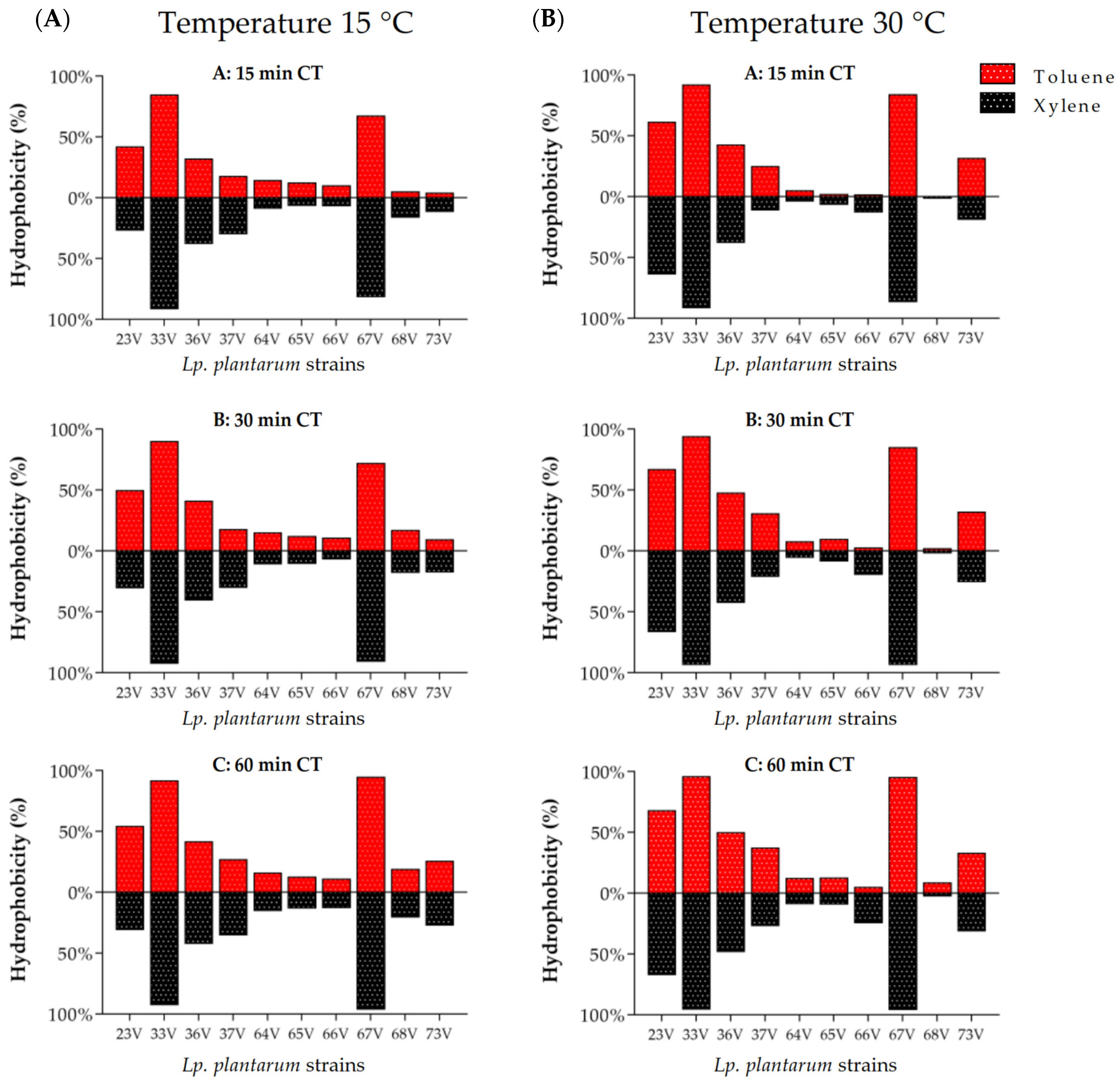
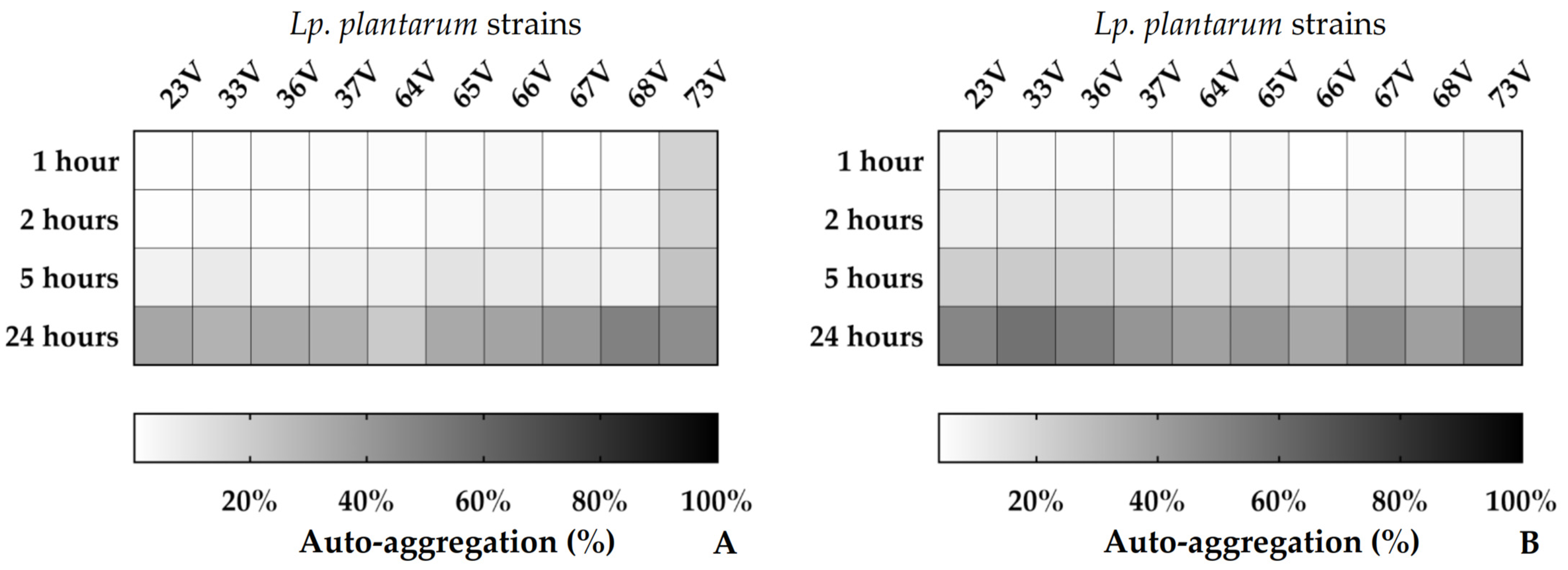
3.3. Cell Surface Properties: Hydrophobicity and Auto-Aggregation
3.4. Antioxidant Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Rodger, H.D. Fish Disease Causing Economic Impact in Global Aquaculture. In Fish Vaccines; Adams, A., Ed.; Springer: Basel, Switzerland, 2016; pp. 1–34. ISBN 978-3-0348-0980-1. [Google Scholar]
- Raja, R.A.; Jithendran, K.P. Aquaculture Disease Diagnosis and Health Management. In Advances in Marine and Brackishwater Aquaculture; Perumal, S., Thirunavukkarasu, A.R., Pachiappan, P., Eds.; Springer: New Delhi, India, 2015; pp. 247–255. ISBN 978-81-322-2271-2. [Google Scholar]
- Sharma, M.; Pandey, G. Overviews of the Treatment and Control of Common Fish Diseases. Int. Res. J. Pharm. 2012, 3, 123–127. [Google Scholar]
- Romero, J.; Feijoó, C.G.; Navarrete, P. Antibiotics in Aquaculture–Use, Abuse and Alternatives. Health Environ. Aquac. 2012, 159. [Google Scholar]
- Reith, M.E.; Singh, R.K.; Curtis, B.; Boyd, J.M.; Bouevitch, A.; Kimball, J.; Munholland, J.; Murphy, C.; Sarty, D.; Williams, J.; et al. The Genome of Aeromonas Salmonicida Subsp. Salmonicida A449: Insights into the Evolution of a Fish Pathogen. BMC Genom. 2008, 9, 427. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C. Heavy Use of Prophylactic Antibiotics in Aquaculture: A Growing Problem for Human and Animal Health and for the Environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food Safety Impacts of Antimicrobial Use and Their Residues in Aquaculture. Public Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef]
- Lulijwa, R.; Rupia, E.J.; Alfaro, A.C. Antibiotic Use in Aquaculture, Policies and Regulation, Health and Environmental Risks: A Review of the Top 15 Major Producers. Rev. Aquac. 2020, 12, 640–663. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, L.; Tiu, L.; Wang, H.H. Characterization of Antibiotic Resistance in Commensal Bacteria from an Aquaculture Ecosystem. Front. Microbiol. 2015, 6, 914. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sánchez, T.; Mora-Sánchez, B.; Balcázar, J.L. Biological Approaches for Disease Control in Aquaculture: Advantages, Limitations and Challenges. Trends Microbiol. 2018, 26, 896–903. [Google Scholar] [CrossRef]
- Gudding, R.; Van Muiswinkel, W.B. A History of Fish Vaccination: Science-Based Disease Prevention in Aquaculture. Fish Shellfish Immunol. 2013, 35, 1683–1688. [Google Scholar] [CrossRef]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Vet. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E. Probiotics in Shellfish Aquaculture. Aquac. Fish. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Chauhan, A.; Singh, R. Probiotics in Aquaculture: A Promising Emerging Alternative Approach. Symbiosis 2019, 77, 99–113. [Google Scholar] [CrossRef]
- Merrifield, D.; Bradley, G.; Baker, R.; Davies, S. Probiotic Applications for Rainbow Trout (Oncorhynchus Mykiss Walbaum) II. Effects on Growth Performance, Feed Utilization, Intestinal Microbiota and Related Health Criteria Postantibiotic Treatment. Aquac. Nutr. 2010, 16, 496–503. [Google Scholar] [CrossRef]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and Prebiotics Associated with Aquaculture: A Review. Fish Shellfish Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.-Z.; Wang, A.; Zhou, Z. Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Van Doan, H.; Lee, S.H.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, Lactic Acid Bacteria and Bacilli: Interesting Supplementation for Aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef] [Green Version]
- Merrifield, D.L.; Carnevali, O. Probiotic Modulation of the Gut Microbiota of Fish. In Aquaculture Nutrition; Wiley: Hoboken, NJ, USA, 2014; pp. 185–222. [Google Scholar]
- Li, X.; Ringø, E.; Hoseinifar, S.H.; Lauzon, H.L.; Birkbeck, H.; Yang, D. The Adherence and Colonization of Microorganisms in Fish Gastrointestinal Tract. Rev. Aquac. 2019, 11, 603–618. [Google Scholar] [CrossRef]
- Ringø, E.; Doan, H.V.; Lee, S.; Song, S.K. Lactic Acid Bacteria in Shellfish: Possibilities and Challenges. Rev. Fish. Sci. Aquac. 2020, 28, 139–169. [Google Scholar] [CrossRef]
- Piccolo, G.; Bovera, F.; Lombardi, P.; Mastellone, V.; Nizza, S.; Di Meo, C.; Marono, S.; Nizza, A. Effect of Lactobacillus Plantarum on Growth Performance and Hematological Traits of European Sea Bass (Dicentrarchus Labrax). Aquac. Int. 2015, 23, 1025–1032. [Google Scholar] [CrossRef]
- Dash, G.; Raman, R.P.; Pani Prasad, K.; Makesh, M.; Pradeep, M.A.; Sen, S. Evaluation of Paraprobiotic Applicability of Lactobacillus Plantarum in Improving the Immune Response and Disease Protection in Giant Freshwater Prawn, Macrobrachium Rosenbergii (de Man, 1879). Fish Shellfish Immunol. 2015, 43, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, H.; He, R.; Xu, W.; Mai, K.; He, G. Effects of Soybean Meal Fermentation by Lactobacillus Plantarum P8 on Growth, Immune Responses, and Intestinal Morphology in Juvenile Turbot (Scophthalmus Maximus L.). Aquaculture 2016, 464, 87–94. [Google Scholar] [CrossRef]
- Van Nguyen, N.; Onoda, S.; Van Khanh, T.; Hai, P.D.; Trung, N.T.; Hoang, L.; Koshio, S. Evaluation of Dietary Heat-Killed Lactobacillus Plantarum Strain L-137 Supplementation on Growth Performance, Immunity and Stress Resistance of Nile Tilapia (Oreochromis Niloticus). Aquaculture 2019, 498, 371–379. [Google Scholar] [CrossRef]
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.; Rademaker, J.L.; Starrenburg, M.J.; Kleerebezem, M.; Molenaar, D.; van Hylckama Vlieg, J.E. Phenotypic and Genomic Diversity of Lactobacillus Plantarum Strains Isolated from Various Environmental Niches. Environ. Microbiol. 2010, 12, 758–773. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Succi, M.; Sorrentino, E.; Petrarca, S.; De Cristofaro, A.; Coppola, R. Inter-and Intra-Species Diversity of Lactic Acid Bacteria in Apis Mellifera Ligustica Colonies. Microorganisms 2020, 8, 1578. [Google Scholar] [CrossRef]
- Iorizzo, M.; Albanese, G.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; D’Andrea, M.; Iaffaldano, N.; Coppola, R. Presence of Lactic Acid Bacteria in the Intestinal Tract of the Mediterranean Trout (Salmo Macrostigma) in Its Natural Environment. Life 2021, 11, 667. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile Resistance Mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Cui, Y.; Qu, X. Mechanisms and Improvement of Acid Resistance in Lactic Acid Bacteria. Arch. Microbiol. 2018, 200, 195–201. [Google Scholar] [CrossRef]
- Chang, M.-H.; Hong, S.-F.; Chen, J.-H.; Lin, M.-F.; Chen, C.-S.; Wang, S.-C. Antibacterial Activity Lactobacillus Plantarum Isolated from Fermented Vegetables and Investigation of the Plantaricin Genes. Afr. J. Microbiol. Res. 2016, 10, 796–803. [Google Scholar]
- Yadav, R.; Puniya, A.K.; Shukla, P. Probiotic Properties of Lactobacillus Plantarum RYPR1 from an Indigenous Fermented Beverage Raabadi. Front. Microbiol. 2016, 7, 1683. [Google Scholar] [CrossRef] [Green Version]
- Tremonte, P.; Pannella, G.; Succi, M.; Tipaldi, L.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial Activity of Lactobacillus Plantarum Strains Isolated from Different Environments: A Preliminary Study. Int. Food Res. J. 2017, 24, 852. [Google Scholar]
- Kaktcham, P.M.; Temgoua, J.-B.; Zambou, F.N.; Diaz-Ruiz, G.; Wacher, C.; de Lourdes Pérez-Chabela, M. In Vitro Evaluation of the Probiotic and Safety Properties of Bacteriocinogenic and Non-Bacteriocinogenic Lactic Acid Bacteria from the Intestines of Nile Tilapia and Common Carp for Their Use as Probiotics in Aquaculture. Probiotics Antimicrob. Proteins 2018, 10, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Kazuń, B.; Kazuń, K.; Żylińska, J.; Siwicki, A.K. In Vitro Study of Lactobacillus Plantarum Properties as a Potential Probiotic Strain and an Alternative Method to Antibiotic Treatment of Fish. Fish. Aquat. Life 2018, 26, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Foysal, M.J.; Fotedar, R.; Siddik, M.A.; Tay, A. Lactobacillus Acidophilus and L. Plantarum Improve Health Status, Modulate Gut Microbiota and Innate Immune Response of Marron (Cherax Cainii). Sci. Rep. 2020, 10, 5916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamad, N.; Manan, H.; Sallehhuddin, M.; Musa, N.; Ikhwanuddin, M. Screening of Lactic Acid Bacteria Isolated from Giant Freshwater Prawn (Macrobrachium Rosenbergii) as Potential Probiotics. Aquac. Rep. 2020, 18, 100523. [Google Scholar] [CrossRef]
- Maji, U.; Mohanty, S. Genotypic Characterization of Lactic Acid Bacteria in Gut Microbiome of Freshwater Fish. Microbiology 2017, 86, 276–285. [Google Scholar] [CrossRef]
- Kumaree, K.K.; Akbar, A.; Anal, A.K. Bioencapsulation and Application of Lactobacillus Plantarum Isolated from Catfish Gut as an Antimicrobial Agent and Additive in Fish Feed Pellets. Ann. Microbiol. 2015, 65, 1439–1445. [Google Scholar] [CrossRef]
- Bucio Galindo, A.; Hartemink, R.; Schrama, J.; Verreth, J.; Bucio, G.L.; Zwietering, M. Kinetics of Lactobacillus Plantarum 44a in the Faeces of Tilapia (Oreochromis Niloticus) after Its Intake in Feed. J. Appl. Microbiol. 2009, 107, 1967–1975. [Google Scholar] [CrossRef]
- Nathanailides, C.; Kolygas, M.; Choremi, K.; Mavraganis, T.; Gouva, E.; Vidalis, K.; Athanassopoulou, F. Probiotics Have the Potential to Significantly Mitigate the Environmental Impact of Freshwater Fish Farms. Fishes 2021, 6, 76. [Google Scholar] [CrossRef]
- Kaewchomphunuch, T.; Charoenpichitnunt, T.; Thongbaiyai, V.; Ngamwongsatit, N.; Kaeoket, K. Cell-Free Culture Supernatants of Lactobacillus Spp. and Pediococcus Spp. Inhibit Growth of Pathogenic Escherichia Coli Isolated from Pigs in Thailand. BMC Vet. Res. 2022, 18, 60. [Google Scholar] [CrossRef]
- Zapata, A.; Ramirez-Arcos, S. A Comparative Study of McFarland Turbidity Standards and the Densimat Photometer to Determine Bacterial Cell Density. Curr. Microbiol. 2015, 70, 907–909. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, K.M.G.M.M.; Yang, S.J.; Lee, N.-K.; Paik, H.-D. Probiotic Properties of Lactobacillus Brevis KU200019 and Synergistic Activity with Fructooligosaccharides in Antagonistic Activity against Foodborne Pathogens. Food Sci. Anim. Resour. 2020, 40, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; Ganassi, S.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A. Antimicrobial Activity against Paenibacillus Larvae and Functional Properties of Lactiplantibacillus Plantarum Strains: Potential Benefits for Honeybee Health. Antibiotics 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, C.; Testa, B.; Letizia, F.; Iorizzo, M.; Lombardi, S.J.; Ianiro, M.; Di Renzo, M.; Strollo, D.; Coppola, R. Effect of Exogenous Proline on the Ethanolic Tolerance and Malolactic Performance of Oenococcus Oeni. J. Food Sci. Technol. 2020, 57, 3973–3979. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Aarti, C.; Khusro, A.; Varghese, R.; Arasu, M.V.; Agastian, P.; Al-Dhabi, N.A.; Ilavenil, S.; Choi, K.C. In Vitro Studies on Probiotic and Antioxidant Properties of Lactobacillus Brevis Strain LAP2 Isolated from Hentak, a Fermented Fish Product of North-East India. LWT 2017, 86, 438–446. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer: Berlin/Heidelberg, Germany, 2007; Volume 26. [Google Scholar]
- Wuertz, S.; Schroeder, A.; Wanka, K.M. Probiotics in Fish Nutrition—Long-Standing Household Remedy or Native Nutraceuticals? Water 2021, 13, 1348. [Google Scholar] [CrossRef]
- Novoslavskij, A.; Terentjeva, M.; Eizenberga, I.; Valciņa, O.; Bartkevičs, V.; Bērziņš, A. Major Foodborne Pathogens in Fish and Fish Products: A Review. Ann. Microbiol. 2016, 66, 1–15. [Google Scholar] [CrossRef]
- Tripathy, S.; Kumar, N.; Mohanty, S.; Samanta, M.; Mandal, R.; Maiti, N. Characterisation of Pseudomonas Aeruginosa Isolated from Freshwater Culture Systems. Microbiol. Res. 2007, 162, 391–396. [Google Scholar] [CrossRef]
- Arfatahery, N.; Mirshafiey, A.; Abedimohtasab, T.P.; Zeinolabedinizamani, M. Study of the Prevalence of Staphylococcus Aureus in Marine and Farmed Shrimps in Iran Aiming the Future Development of a Prophylactic Vaccine. Procedia Vaccinol. 2015, 9, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Austin, B.; Stobie, M.; Robertson, P. Citrobacter Freundii: The Cause of Gastro-Enteritis Leading to Progressive Low Level Mortalities in Farmed Rainbow Trout, Oncorhynchus Mykiss Walbaum, in Scotland. Bull. Eur. Assoc. Fish Pathol. UK 1992, 12, 166–167. [Google Scholar]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; del Campo, R.; Hernández, P.E.; Herranz, C.; Cintas, L.M. Antimicrobial Activity, Antibiotic Susceptibility and Virulence Factors of Lactic Acid Bacteria of Aquatic Origin Intended for Use as Probiotics in Aquaculture. BMC Microbiol. 2013, 13, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilova, T.; Adzhieva, A.; Danilina, G.; Polyakov, N.; Soloviev, A.; Zhukhovitsky, V. Antimicrobial Activity of Supernatant of Lactobacillus Plantarum against Pathogenic Microorganisms. Bull. Exp. Biol. Med. 2019, 167, 751–754. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T.; Sorgeloos, P.; Bossier, P. Alternatives to Antibiotics for the Control of Bacterial Disease in Aquaculture. Curr. Opin. Microbiol. 2011, 14, 251–258. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.; Austin, B. Probiotics for Disease Control in Aquaculture. Front. Microbiol. 2017, 189–222. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant Defenses in Fish: Biotic and Abiotic Factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Wilhelm Filho, D. Reactive Oxygen Species, Antioxidants and Fish Mitochondria. Front Biosci. 2007, 12, 1229–1237. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2021, 29, 198–217. [Google Scholar] [CrossRef]
- Chowdhury, S.; Saikia, S. Oxidative Stress in Fish: A Review. J. Sci. Res. 2020, 12, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Lushchak, V.I. Environmentally Induced Oxidative Stress in Aquatic Animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as Potential Antioxidants: A Systematic Review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial Roles of Feed Additives as Immunostimulants in Aquaculture: A Review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Ringø, E.; Ángeles Esteban, M.; Dadar, M.; Dawood, M.A.; Faggio, C. Host-Associated Probiotics: A Key Factor in Sustainable Aquaculture. Rev. Fish. Sci. Aquac. 2020, 28, 16–42. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J. Oxidative Stress Tolerance and Antioxidant Capacity of Lactic Acid Bacteria as Probiotic: A Systematic Review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Genesis and Development of DPPH Method of Antioxidant Assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Chanput, W.; Krueyos, N.; Ritthiruangdej, P. Anti-Oxidative Assays as Markers for Anti-Inflammatory Activity of Flavonoids. Int. Immunopharmacol. 2016, 40, 170–175. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The Hydrophilic and Lipophilic Contribution to Total Antioxidant Activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary Postbiotic Lactobacillus Plantarum Improves Serum and Ruminal Antioxidant Activity and Upregulates Hepatic Antioxidant Enzymes and Ruminal Barrier Function in Post-Weaning Lambs. Antioxidants 2020, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Li, C.; He, Z.; Pan, F.; Pan, S.; Wang, Y. Probiotic Properties and Cellular Antioxidant Activity of Lactobacillus Plantarum MA2 Isolated from Tibetan Kefir Grains. Probiotics Antimicrob. Proteins 2018, 10, 523–533. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant Activity of Lactobacillus Plantarum Strains Isolated from Traditional Chinese Fermented Foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Goyal, A. Antioxidant Activity and γ-Aminobutyric Acid (GABA) Producing Ability of Probiotic Lactobacillus Plantarum DM5 Isolated from Marcha of Sikkim. LWT-Food Sci. Technol. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Grajek, K.; Sip, A.; Foksowicz-Flaczyk, J.; Dobrowolska, A.; Wita, A. Adhesive and Hydrophobic Properties of the Selected LAB Isolated from Gastrointestinal Tract of Farming Animals. Acta Biochim. Pol. 2016, 63, 311–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and Aggregation Ability of Probiotic Strain Lactobacillus Acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Darmastuti, A.; Hasan, P.N.; Wikandari, R.; Utami, T.; Rahayu, E.S.; Suroto, D.A. Adhesion Properties of Lactobacillus Plantarum Dad-13 and Lactobacillus Plantarum Mut-7 on Sprague Dawley Rat Intestine. Microorganisms 2021, 9, 2336. [Google Scholar] [CrossRef]
- Saboktakin-Rizi, M.; Behbahani, B.A.; Hojjati, M.; Noshad, M. Identification of Lactobacillus Plantarum TW29-1 Isolated from Iranian Fermented Cereal-Dairy Product (Yellow Zabol Kashk): Probiotic Characteristics, Antimicrobial Activity and Safety Evaluation. J. Food Meas. Charact. 2021, 15, 2615–2624. [Google Scholar] [CrossRef]
- Bucio, A.; Hartemink, R.; Schrama, J.W.; Verreth, J.; Rombouts, F.M. Survival of Lactobacillus Plantarum 44a after Spraying and Drying in Feed and during Exposure to Gastrointestinal Tract Fluids in Vitro. J. Gen. Appl. Microbiol. 2005, 51, 221–227. [Google Scholar] [CrossRef] [Green Version]
- He, E.; Wurtsbaugh, W.A. An Empirical Model of Gastric Evacuation Rates for Fish and an Analysis of Digestion in Piscivorous Brown Trout. Trans. Am. Fish. Soc. 1993, 122, 717–730. [Google Scholar] [CrossRef]
- Handeland, S.O.; Imsland, A.K.; Stefansson, S.O. The Effect of Temperature and Fish Size on Growth, Feed Intake, Food Conversion Efficiency and Stomach Evacuation Rate of Atlantic Salmon Post-Smolts. Aquaculture 2008, 283, 36–42. [Google Scholar] [CrossRef]
- Miegel, R.P.; Pain, S.J.; van Wettere, W.H.E.J.; Howarth, G.S.; Stone, D.A.J. Effect of Water Temperature on Gut Transit Time, Digestive Enzyme Activity and Nutrient Digestibility in Yellowtail Kingfish (Seriola Lalandi). Aquaculture 2010, 308, 145–151. [Google Scholar] [CrossRef]
- Volkoff, H.; Rønnestad, I. Effects of Temperature on Feeding and Digestive Processes in Fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Little, A.G.; Loughland, I.; Seebacher, F. What Do Warming Waters Mean for Fish Physiology and Fisheries? J. Fish Biol. 2020, 97, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; van Hylckama Vlieg, J.E. Genomic Diversity and Versatility of Lactobacillus Plantarum, a Natural Metabolic Engineer. Microb. Cell Factories 2011, 10, S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, K.E.; Farrell, A.P. Influence of Seasonal Temperature on the Repeat Swimming Performance of Rainbow Trout Oncorhynchus Mykiss. J. Exp. Biol. 2003, 206, 3569–3579. [Google Scholar] [CrossRef] [Green Version]
- López-Olmeda, J.; Sánchez-Vázquez, F. Thermal Biology of Zebrafish (Danio Rerio). J. Therm. Biol. 2011, 36, 91–104. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Indicator Strains | Lp. plantarum Strains | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 23V | 33V | 36V | 37V | 64V | 65V | 66V | 67V | 68V | 73V | |
| L. innocua | 0 ± 0 d | 0 ± 0 d | 0 ± 0 d | 0 ± 0 d | 0 ± 0 d | 0 ± 0 d | 7.1 ± 0.2 a | 5.0 ± 0.3 c | 5.5 ± 0.4 b | 5.9 ± 0.5 b |
| A. salmonicida | 7.0 ± 0.6 b | 7.0 ± 0.5 b | 7.7 ± 0.6 a | 7.5 ± 0.6 a | 8.5 ± 0.3 a | 7.3 ± 0.3 a | 6.6 ± 0.6 b | 7.5 ± 0.3 a | 8.0 ± 0.2 a | 7.7 ± 0.6 a |
| C. freundii | 7.2 ± 0.7 c | 7.2 ± 0.6 c | 8.0 ± 0.4 b | 6.4 ± 0.6 c | 9.9 ± 0.4 a | 7.0 ± 0.4 c | 10.5 ± 0.4 a | 9.0 ± 0.6 b | 10.2 ± 0.3 a | 10.3 ± 0.5 a |
| P. mirabilis | 9.2 ± 0.6 a | 7.4 ± 0.6 b | 8.3 ± 0.5 b | 9.7 ± 0.4 a | 9.5 ± 0.5 a | 9.0 ± 0.2 a | 9.6 ± 0.4 a | 10.1 ± 0.2 a | 9.5 ± 0.4 a | 7.3 ± 0.6 b |
| S. aureus | 0 ± 0 c | 0 ± 0 c | 0 ± 0 c | 0 ± 0 c | 0 ± 0 c | 0 ± 0 c | 6.5 ± 0.4 b | 7.1 ± 0.5 b | 8.9 ± 0.3 a | 6.9 ± 0.4 b |
| Ps. aeruginosa | 7.0 ± 0.3 a | 8.0 ± 0.3 a | 7.9 ± 0.6 a | 8.0 ± 0.7 a | 7.9 ± 0.6 a | 6.8 ± 0.3 a | 6.0 ± 0.1 b | 7.3 ± 0.4 a | 7.8 ± 0.4 a | 5.8 ± 0.2 b |
| E. coli | 8.8 ± 0.6 b | 9.9 ± 0.6 a | 9.6 ± 0.5 a | 10.1 ± 0.1 a | 9.8 ± 0.2 a | 6.9 ± 0.4 c | 7.0 ± 0.3 c | 7.8 ± 0.3 b | 8.9 ± 0.6 a | 7.1 ± 0.1 c |
| Indicator Strains | Lp. plantarum Strains | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 23V | 33V | 36V | 37V | 64V | 65V | 66V | 67V | 68V | 73V | |
| L. innocua | 7.9 ± 0.3 b | 7.9 ± 0.2 b | 7.0 ± 0.4 c | 8.0 ± 0.3 b | 9.0 ± 0.3 a | 9.0 ± 0.3 a | 5.9 ± 0.4 c | 10.0 ± 0.5 a | 8.2 ± 0.8 b | 9.0 ± 0.2 a |
| A. salmonicida | 7.8 ±0.6 a | 8.2 ± 0.6 a | 8.5 ± 0.5 a | 8.9 ± 0.3 a | 8.4 ± 0.5 a | 7.2 ± 0.8 b | 7.4 ± 1.0 a | 7.7 ± 0.5 a | 7.2 ± 0.4 b | 7.9 ± 0.2 a |
| C. freundii | 7.9 ± 0.7 b | 8.7 ± 0.8 b | 8.8 ± 0.9 b | 9.8 ± 1.2 b | 14.0 ± 0.8 a | 9.1 ± 0.8 b | 7.9 ± 0.3 b | 7.9 ± 0.4 b | 8.9 ± 0.6 b | 6.7 ± 0.8 c |
| P. mirabilis | 7.0 ± 0.5 a | 8.0 ± 0.3 a | 8.0 ± 0.4 a | 7.1 ± 0.6 a | 6.1 ± 0.8 b | 6.9 ± 0.6 a | 5.2 ± 0.5 b | 5.8 ± 0.4 b | 6.4 ± 0.5 b | 7.0 ± 0.6 a |
| S. aureus | 5.9 ± 0.7 b | 5.9 ± 0.6 b | 7.0 ± 0.4 a | 6.8 ± 0.2 a | 6.8 ± 0.6 a | 6.7 ± 0.2 a | 7.0 ± 0.6 a | 7.9 ± 0.6 a | 7.2 ± 0.4 a | 6.2 ± 0.6 b |
| Ps. aeruginosa | 7.2 ± 0.9 a | 7.0 ± 0.9 a | 6.4 ± 0.5 a | 6.8 ± 0.6 a | 6.9 ± 0.5 a | 6.0 ± 0.6 a | 5.8 ± 0.5 a | 7.4 ± 0.4 a | 7.0 ± 0.3 a | 6.0 ± 0.5 a |
| E. coli | 9.0 ± 0.5 a | 9.0 ± 0.6 a | 8.6 ± 0.4 a | 9.9 ± 1.0 a | 9.9 ± 1.0 a | 10.2 ± 1.0 a | 8.0 ± 0.8 b | 7.9 ± 0.5 b | 8.0 ± 0.7 b | 7.0 ± 0.6 b |
| Antioxidant Assay | Lp. plantarum Strains | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 23V | 33V | 36V | 37V | 64V | 65V | 66V | 67V | 68V | 73V | |
| ABTS | 24.1 ± 0.3 d | 30.9 ± 0.7 b | 31.5 ± 1.1 b | 22.6 ± 0.2 d | 31.0 ± 1.1 b | 38.0 ± 0.7 a | 29.1 ± 0.9 b | 31.2 ± 1.4 b | 27.4 ± 2.3 c | 25.9 ± 0.6 c |
| DPPH | 1.0 ± 0.7 b | 1.3 ± 0.3 a | 1.6 ± 0.1 a | 0.8 ± 0.4 b | 1.3 ± 0.1 a | 1.5 ± 0.6 a | 1.0 ± 0.4 b | 2.2 ± 0.1 a | 2.1 ± 0.3 a | 1.2 ± 0.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iorizzo, M.; Albanese, G.; Letizia, F.; Testa, B.; Tremonte, P.; Vergalito, F.; Lombardi, S.J.; Succi, M.; Coppola, R.; Sorrentino, E. Probiotic Potentiality from Versatile Lactiplantibacillus plantarum Strains as Resource to Enhance Freshwater Fish Health. Microorganisms 2022, 10, 463. https://doi.org/10.3390/microorganisms10020463
Iorizzo M, Albanese G, Letizia F, Testa B, Tremonte P, Vergalito F, Lombardi SJ, Succi M, Coppola R, Sorrentino E. Probiotic Potentiality from Versatile Lactiplantibacillus plantarum Strains as Resource to Enhance Freshwater Fish Health. Microorganisms. 2022; 10(2):463. https://doi.org/10.3390/microorganisms10020463
Chicago/Turabian StyleIorizzo, Massimo, Gianluca Albanese, Francesco Letizia, Bruno Testa, Patrizio Tremonte, Franca Vergalito, Silvia Jane Lombardi, Mariantonietta Succi, Raffaele Coppola, and Elena Sorrentino. 2022. "Probiotic Potentiality from Versatile Lactiplantibacillus plantarum Strains as Resource to Enhance Freshwater Fish Health" Microorganisms 10, no. 2: 463. https://doi.org/10.3390/microorganisms10020463