
Enhanced Production of (R)-3-Hydroxybutyrate Oligomers by Coexpression of Molecular Chaperones in Recombinant Escherichia coli Harboring a Polyhydroxyalkanoate Synthase Derived from Bacillus cereus YB-4


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Culture Conditions for 3HBO Production
2.3. Quantification of Extracellular 3HBOs/3HB, Extracellular Ethanol, Intracellular 3HBOs and P(3HB)
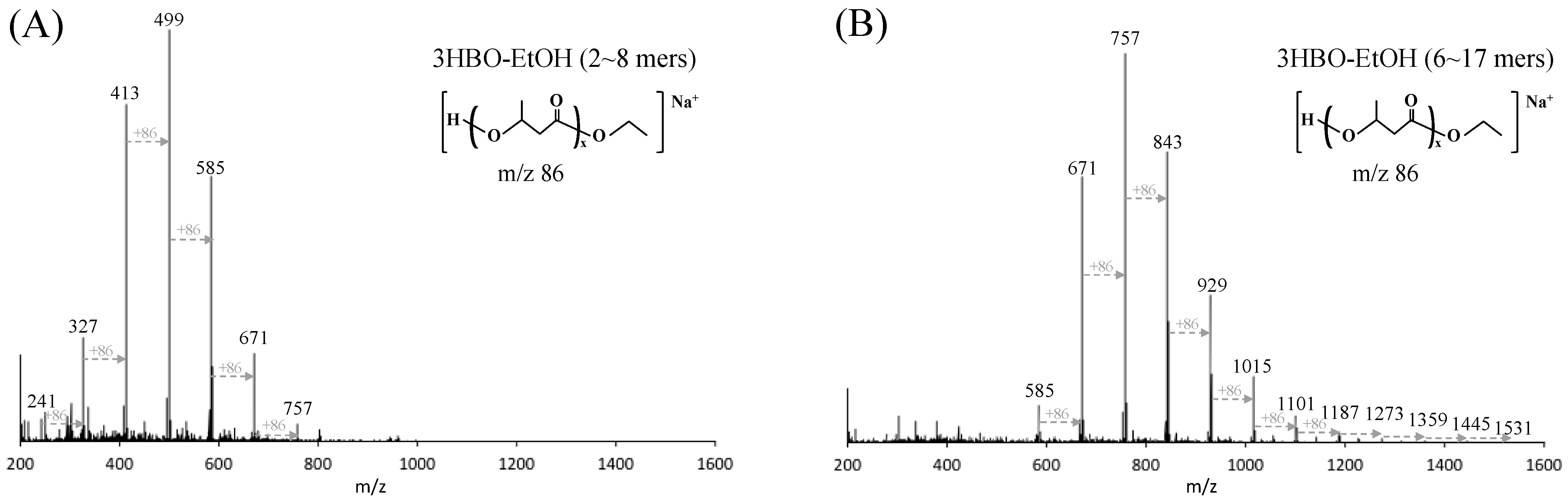
2.4. Extraction of 3HBOs and ESI-TOF-MS Analysis
2.5. Purification of P(3HB) and GPC Analysis
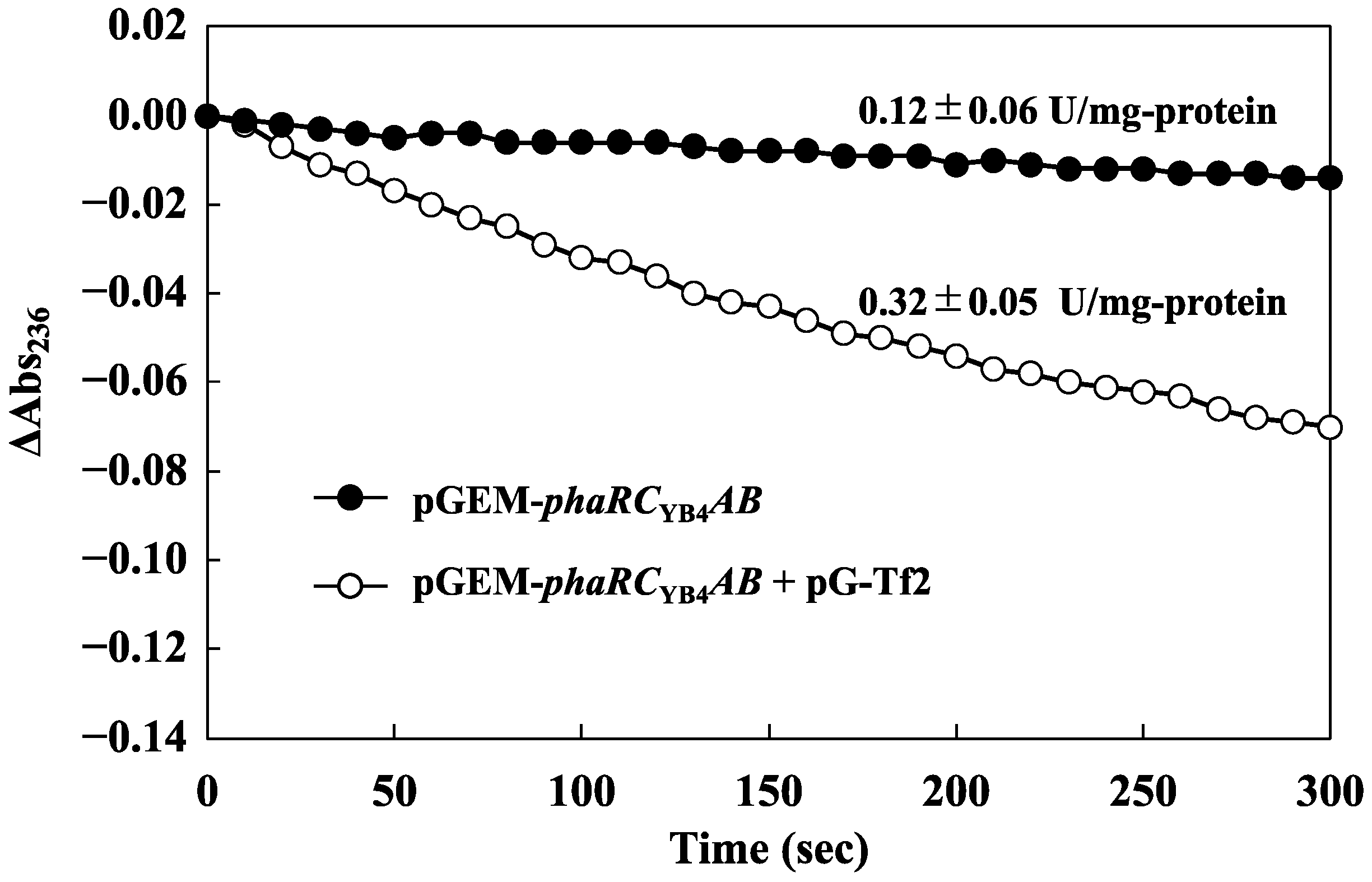
2.6. PHA Synthase Activity Assay
2.7. Expression of PhaRYB4 and PhaCYB4 with Molecular Chaperones
3. Results
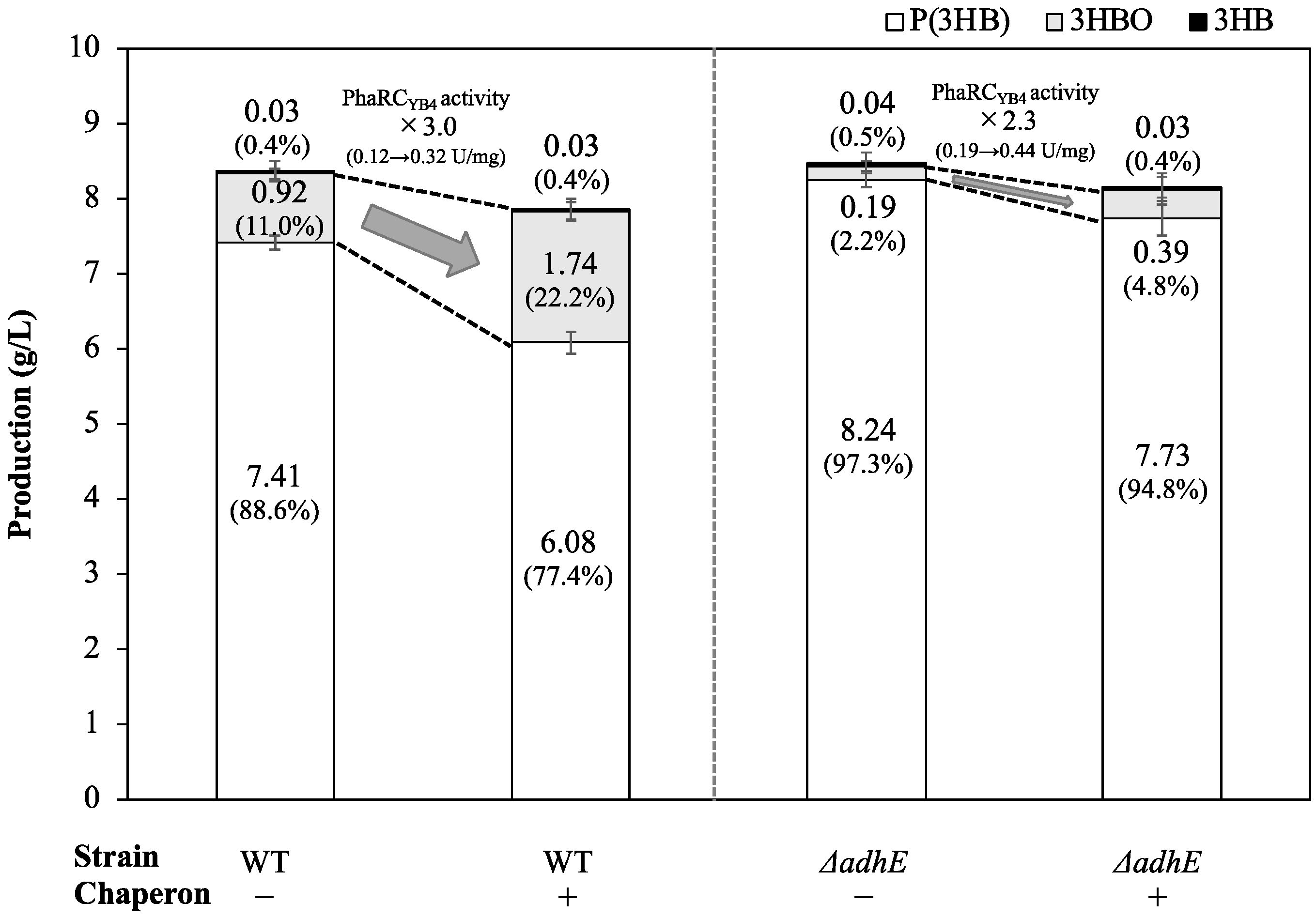
3.1. Effect of Chaperones on 3HBO Production
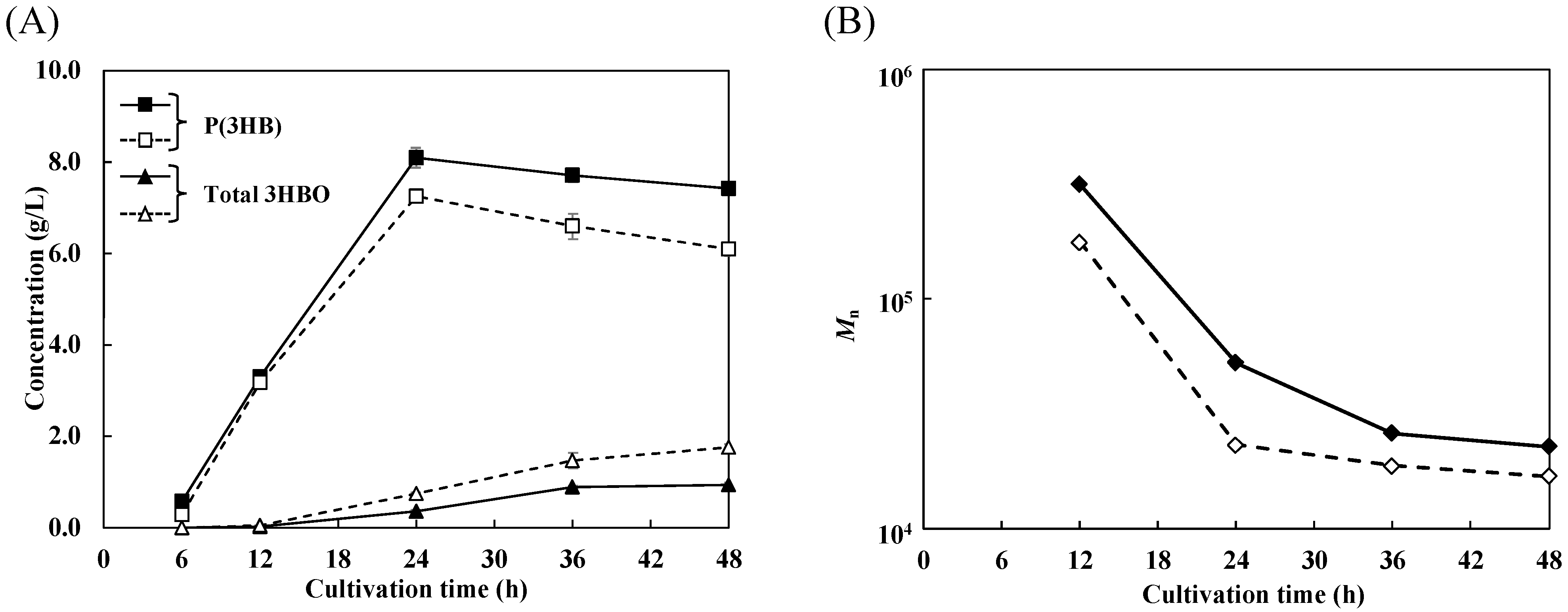
3.2. Time Course Pattern of 3HBO Production with Coexpression of Molecular Chaperones
3.3. Production of 3HBOs in Non-Ethanol-Producing E. coli with Coexpression of Molecular Chaperones
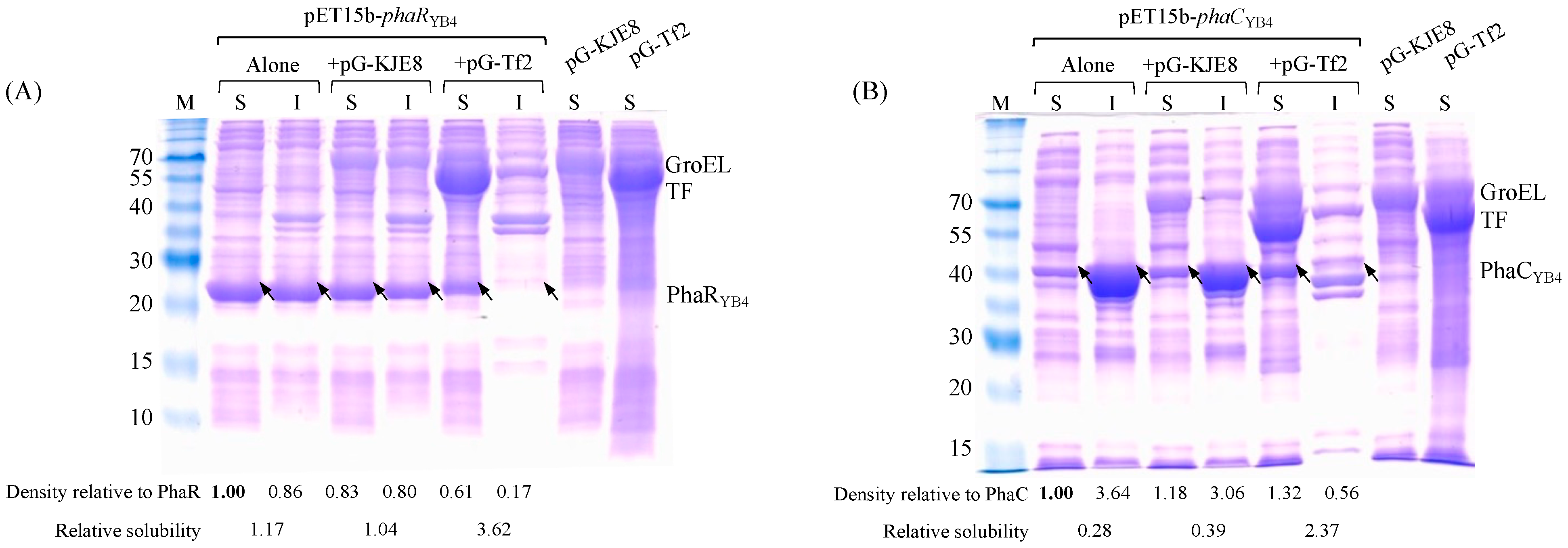
3.4. Effect of Chaperones on PhaRYB4 and PhaCYB4 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinbüchel, A.; Valentin, H.E. Diversity of bacterial polyhydroxyalkanoic acids. FEMS Microbiol. Lett. 1995, 128, 219–228. [Google Scholar] [CrossRef]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Chanprateep, S. Current trends in biodegradable polyhydroxyalkanoates. J. Biosci. Bioeng. 2010, 110, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Rehm, B.H.A. Biogenesis of microbial polyhydroxyalkanoate granules: A platform technology for the production of tailor-made bioparticles. Curr. Issues Mol. Biol. 2007, 9, 41–62. [Google Scholar]
- Tsuge, T.; Hyakutake, M.; Mizuno, K. Class IV polyhydroxyalkanoate (PHA) synthases and PHA-producing Bacillus. Appl. Microbiol. Biotechnol. 2015, 99, 6231–6240. [Google Scholar] [CrossRef] [PubMed]
- Nomura, C.T.; Taguchi, S. PHA synthase engineering toward superbiocatalysts for custom-made biopolymers. Appl. Microbiol. Biotechnol. 2007, 73, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Shi, M.; Zhang, T.; Li, L.; Li, L.; Xian, M. Natural and engineered polyhydroxyalkanoate (PHA) synthase: Key enzyme in biopolyester production. Appl. Microbiol. Biotechnol. 2017, 101, 7417–7426. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, Y.; Wu, Q.; Wang, Y.; Chen, G.Q. Synthetic biology and genome-editing tools for improving PHA metabolic engineering. Trends Biotechnol. 2020, 38, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Maruyama, H.; Fujiki, T.; Matsumoto, K. Regulation of 3-hydroxyhexanoate composition in PHBH synthesized by recombinant Cupriavidus necator H16 from plant oil by using butyrate as a co-substrate. J. Biosci. Bioeng. 2015, 120, 246–251. [Google Scholar] [CrossRef]
- Matsusaki, H.; Abe, H.; Doi, Y. Biosynthesis and properties of poly(3-hydroxybutyrate-co-3-hydroxyalkanoates) by recombinant strains of Pseudomonas sp. 61-3. Biomacromolecules 2000, 1, 17–22. [Google Scholar] [CrossRef]
- Yamada, M.; Matsumoto, K.; Uramoto, S.; Motohashi, R.; Abe, H.; Taguchi, S. Lactate fraction dependent mechanical properties of semitransparent poly(lactate-co-3-hydroxybutyrate)s produced by control of lactyl-CoA monomer fluxes in recombinant Escherichia coli. J. Biotechnol. 2011, 154, 255–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroe, A.; Ishii, N.; Ishii, D.; Kabe, T.; Abe, H.; Iwata, T.; Tsuge, T. Uniformity of monomer composition and material properties of medium-chain-length polyhydroxyalkanoates biosynthesized from pure and crude fatty acids. ACS Sustain. Chem. Eng. 2016, 4, 6905–6911. [Google Scholar] [CrossRef]
- Utsunomia, C.; Matsumoto, K.; Taguchi, S. Microbial secretion of D-lactate-based oligomers. ACS Sustain. Chem. Eng. 2017, 5, 2360–2367. [Google Scholar] [CrossRef]
- Utsunomia, C.; Matsumoto, K.; Date, S.; Hori, C.; Taguchi, S. Microbial secretion of lactate-enriched oligomers for efficient conversion into lactide: A biological shortcut to polylactide. J. Biosci. Bioeng. 2017, 124, 204–208. [Google Scholar] [CrossRef]
- Utsunomia, C.; Hori, C.; Matsumoto, K.; Taguchi, S. Investigation of the Escherichia coli membrane transporters involved in the secretion of D-lactate-based oligomers by loss-of-function screening. J. Biosci. Bioeng. 2017, 124, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, Y.; Hiroe, A.; Tsuge, T.; Taguchi, S. Microbial secretion platform for 3-hydroxybutyrate oligomer and its end-capped forms using chain transfer reaction-mediated polyhydroxyalkanoate synthases. Biotechnol. J. 2019, 14, e1900201. [Google Scholar] [CrossRef] [PubMed]
- Hiroe, A.; Sakurai, T.; Mizuno, S.; Miyahara, Y.; Goto, S.; Yamada, M.; Tsuge, T.; Taguchi, S. Microbial oversecretion of (R)-3-hydroxybutyrate oligomer with diethylene glycol terminal as a macromonomer for polyurethane synthesis. Int. J. Biol. Macromol. 2021, 167, 1290–1296. [Google Scholar] [CrossRef]
- Nduko, J.M.; Taguchi, S. Microbial production of biodegradable lactate-based polymers and oligomeric building blocks from renewable and waste resources. Front. Bioeng. Biotechnol. 2021, 8, 618077. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Konishi, H.; Shimao, M.; Sakazawa, C. Production of 3-hydroxybutyric acid trimer by Bacillus megaterium B-124. J. Ferment. Bioeng. 1992, 73, 246–247. [Google Scholar] [CrossRef]
- Schulz, S.; Toft, S. Identification of a sex pheromone from a spider. Science 1993, 260, 1635–1637. [Google Scholar] [CrossRef]
- Ogita, N.; Hashidoko, Y.; Limin, S.H.; Tahara, S. Linear 3-hydroxybutyrate tetramer (HB4) produced by Sphingomonas sp. is characterized as a growth promoting factor for some rhizomicrofloral composers. Biosci. Biotechnol. Biochem. 2006, 70, 2325–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskimäki, J.J.; Kajula, M.; Hokkanen, J.; Ihantola, E.L.; Kim, J.H.; Hautajärvi, H.; Hankala, E.; Suokas, M.; Pohjanen, J.; Podolich, O.; et al. Methyl-esterified 3-hydroxybutyrate oligomers protect bacteria from hydroxyl radicals. Nat. Chem. Biol. 2016, 12, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, Z.; Li, J.; Yang, X.; Fei, B.; Leung, P.H.M.; Tao, X. A new antimicrobial agent: Poly(3-hydroxybutyric acid) oligomer. Macromol. Biosci. 2019, 19, 1970014. [Google Scholar] [CrossRef] [Green Version]
- Utsunomia, C.; Saito, T.; Matsumoto, K.; Hori, C.; Isono, T.; Satoh, T.; Taguchi, S. Synthesis of lactate (LA)-based poly(ester-urethane) using hydroxyl-terminated LA-based oligomers from a microbial secretion system. J. Polym. Res. 2017, 24, 167. [Google Scholar] [CrossRef]
- Hyakutake, M.; Tomizawa, S.; Mizuno, K.; Abe, H.; Tsuge, T. Alcoholytic cleavage of polyhydroxyalkanoate chains by Class IV synthases induced by endogenous and exogenous ethanol. Appl. Environ. Microbiol. 2014, 80, 1421–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyakutake, M.; Tomizawa, S.; Mizuno, K.; Hisano, T.; Abe, H.; Tsuge, T. A common active site of polyhydroxyalkanoate synthase from Bacillus cereus YB-4 is involved in polymerization and alcoholysis reactions. Appl. Microbiol. Biotechnol. 2015, 99, 4701–4711. [Google Scholar] [CrossRef]
- Tsuge, T. Fundamental factors determining the molecular weight of polyhydroxyalkanoate during biosynthesis. Polym. J. 2016, 48, 1051–1057. [Google Scholar] [CrossRef]
- Hiroe, A.; Ushimaru, K.; Tsuge, T. Characterization of polyhydroxyalkanoate (PHA) synthase derived from Delftia acidovorans DS-17 and the influence of PHA production in Escherichia coli. J. Biosci. Bioeng. 2013, 115, 633–638. [Google Scholar] [CrossRef]
- Thomson, N.M.; Saika, A.; Ushimaru, K.; Sangiambut, S.; Tsuge, T.; Summers, D.K.; Sivaniah, E. Efficient production of active polyhydroxyalkanoate synthase in Escherichia coli by coexpression of molecular chaperones. Appl. Environ. Microbiol. 2013, 79, 1948–1955. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.; Bukau, B.; Kramer, G. Structure and function of the molecular chaperone Trigger Factor. Biochim. Biophys. Acta 2010, 1803, 650–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Thomas, J.G.; Ayling, A.; Baneyx, F. Molecular chaperones, folding catalysts, and the recovery of active recombinant proteins from E. coli. Appl. Biochem. Biotechnol. 1997, 66, 197–238. [Google Scholar] [CrossRef] [PubMed]
- Kerner, M.J.; Naylor, D.J.; Ishihama, Y.; Maier, T.; Chang, H.C.; Stines, A.P.; Georgopoulos, C.; Frishman, D.; Hayer-Hartl, M.; Mann, M.; et al. Proteome-wide analysis of chaperonin-dependent protein folding in Escherichia coli. Cell 2005, 122, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomizawa, S.; Hyakutake, M.; Saito, Y.; Agus, J.; Mizuno, K.; Abe, H.; Tsuge, T. Molecular weight change of polyhydroxyalkanoate (PHA) caused by the PhaC subunit of PHA synthase from Bacillus cereus YB-4 in recombinant Escherichia coli. Biomacromolecules 2011, 12, 2660–2666. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [Green Version]
- Kato, M.; Bao, H.J.; Kang, C.K.; Fukui, T.; Doi, Y. Production of a novel copolyester of 3-hydroxybutyric acid and medium-chain-length 3-hydroxyalkanoic acids by Pseudomonas sp. 61-3 from sugars. Appl. Microbiol. Biotechnol. 1996, 45, 363–370. [Google Scholar] [CrossRef]
- Fukui, T.; Yoshimoto, A.; Matsumoto, M.; Hosokawa, S.; Saito, T.; Nishikawa, H.; Tomita, K. Enzymatic synthesis of poly-β-hydroxybutyrate in Zoogloea ramigera. Arch. Microbiol. 1976, 110, 149–156. [Google Scholar] [CrossRef]
- Kihara, T.; Hiroe, A.; Ishii-Hyakutake, M.; Mizuno, K.; Tsuge, T. Bacillus cereus-type polyhydroxyalkanoate biosynthetic gene cluster contains R-specific enoyl-CoA hydratase gene. Biosci. Biotechnol. Biochem. 2017, 81, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Niwa, T.; Kanamori, T.; Ueda, T.; Taguchi, H. Global analysis of chaperone effects using a reconstituted cell-free translation system. Proc. Natl. Acad. Sci. USA 2012, 109, 8937–8942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, T.; Ying, B.W.; Saito, K.; Jin, W.; Takada, S.; Ueda, T.; Taguchi, H. Bimodal protein solubility distribution revealed by an aggregation analysis of the entire ensemble of Escherichia coli proteins. Proc. Natl. Acad. Sci. USA 2009, 106, 4201–4206. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Relevant Characteristics | Reference/Source |
|---|---|---|
| Strains | ||
| Escherichia coli BW25113 | rrnBT14 ΔlacZWJ16 hsdR514 ΔaraBADAH33 | [35] |
| E. coli BW25113 ΔadhE | BW25113, ΔadhE::FRT-Km-FRT | [35] |
| E. coli BL21(DE3) | F− ompT hsdSB(rB−mB−) gal dcm | Novagen |
| Plasmids | ||
| pGEM-phaRCYB4AB | pGEM-T derivative; phaRe promoter, phaRCYB4 from B. cereus YB-4, and phaABRe from Ralstonia eutropha (Cupriavidus necator), Apr | [34] |
| pG-Tf2 | Expression vector for GroEL/GroES/Tf; pzt1 promoter; Cmr | Takara Bio. |
| pG-KJE8 | Expression vector for GroEL/GroES with pzt1 promoter; DnaK, DnaJ, GrpE with araB promoter; Cmr | Takara Bio. |
| pET15b-phaRYB4 | pET15b derivative; expression of N-terminal His-tagged phaRYB4 from B. cereus YB-4, Apr | [25] |
| pET15b-phaCYB4 | pET15b derivative; expression of N-terminal His-tagged phaCYB4 from B. cereus YB-4, Apr | [25] |
| Plasmid | Dry Cell Weight (g/L) | Residual Cell Mass (g/L) | Intracellular P(3HB) (g/L) | Intracellular 3HBOs (g/L) | Extracellular 3HBOs (g/L) | Extracellular 3HB (g/L) | Molecular Weight of P(3HB) | ||
|---|---|---|---|---|---|---|---|---|---|
| Mn (×104) | Mw (×104) | Mw/Mn | |||||||
| pGEM-phaRCYB4AB | 9.40 ± 0.05 | 1.72 ± 0.01 | 7.41 ± 0.09 | 0.27 ± 0.03 | 0.65 ± 0.04 | 0.03 ± 0.00 | 2.3 ± 0.0 | 5.0 ± 0.1 | 2.2 |
| pGEM-phaRCYB4AB + pG-Tf2 | 8.81 ± 0.08 | 2.22 ± 0.12 | 6.08 ± 0.15 | 0.50 ± 0.04 | 1.24 ± 0.08 | 0.03 ± 0.00 | 1.7 ± 0.1 | 3.5 ± 0.0 | 2.2 |
| Plasmid | Dry Cell Weight (g/L) | Residual Cell Mass (g/L) | Intracellular P(3HB) (g/L) | Intracellular 3HBOs (g/L) | Extracellular 3HBOs (g/L) | Extracellular 3HB (g/L) | Molecular Weight of P(3HB) | ||
|---|---|---|---|---|---|---|---|---|---|
| Mn (×104) | Mw (×104) | Mw /Mn | |||||||
| pGEM-phaRCYB4AB | 9.67 ± 0.02 | 1.42 ± 0.09 | 8.24 ± 0.07 | 0.01 ± 0.00 | 0.18 ± 0.01 | 0.04 ± 0.00 | 20.4 ± 1.3 | 151.9 ± 5.4 | 7.5 |
| pGEM-phaRCYB4AB + pG-Tf2 | 9.77 ± 0.02 | 2.03 ± 0.23 | 7.73 ± 0.21 | 0.02 ± 0.00 | 0.37 ± 0.02 | 0.03 ± 0.00 | 9.4 ± 0.5 | 76.5 ± 5.8 | 8.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goto, S.; Miyahara, Y.; Taguchi, S.; Tsuge, T.; Hiroe, A. Enhanced Production of (R)-3-Hydroxybutyrate Oligomers by Coexpression of Molecular Chaperones in Recombinant Escherichia coli Harboring a Polyhydroxyalkanoate Synthase Derived from Bacillus cereus YB-4. Microorganisms 2022, 10, 458. https://doi.org/10.3390/microorganisms10020458
Goto S, Miyahara Y, Taguchi S, Tsuge T, Hiroe A. Enhanced Production of (R)-3-Hydroxybutyrate Oligomers by Coexpression of Molecular Chaperones in Recombinant Escherichia coli Harboring a Polyhydroxyalkanoate Synthase Derived from Bacillus cereus YB-4. Microorganisms. 2022; 10(2):458. https://doi.org/10.3390/microorganisms10020458
Chicago/Turabian StyleGoto, Saki, Yuki Miyahara, Seiichi Taguchi, Takeharu Tsuge, and Ayaka Hiroe. 2022. "Enhanced Production of (R)-3-Hydroxybutyrate Oligomers by Coexpression of Molecular Chaperones in Recombinant Escherichia coli Harboring a Polyhydroxyalkanoate Synthase Derived from Bacillus cereus YB-4" Microorganisms 10, no. 2: 458. https://doi.org/10.3390/microorganisms10020458