
Transcriptomic Analysis of Degradative Pathways for Azo Dye Acid Blue 113 in Sphingomonas melonis B-2 from the Dye Wastewater Treatment Process

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Microorganisms Efficient for Azo Dye Biodegradation and Their Identification
2.2. Experimental Culture Setup for Biodegradation of the Azo Dye Acid Blue 113
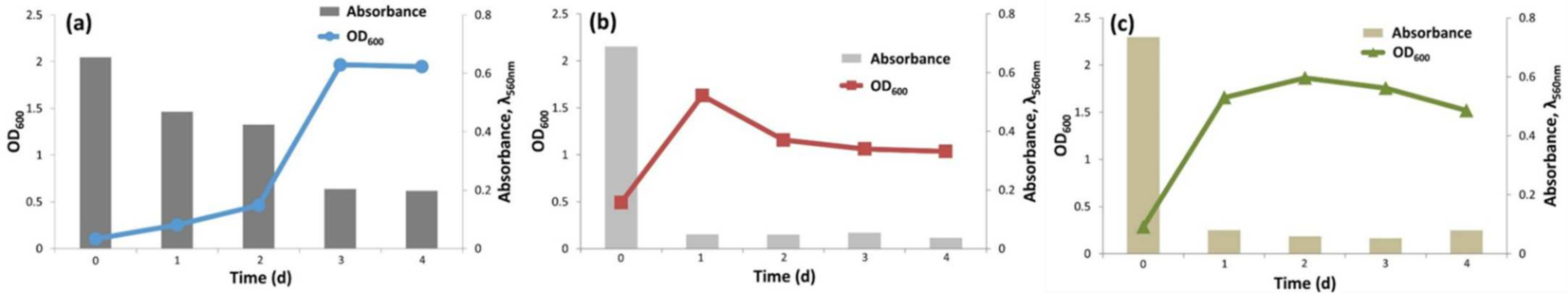
2.3. Monitoring the Cell Growth and Dye Decolorization
2.4. RNA Extraction and cDNA Library Sequencing of the Transcriptome
2.5. De Novo Assembly of the S. melonis B-2 Transcriptome
2.6. Analysis of Differential Gene Expression Involved in Azo Dye Degradation
3. Results
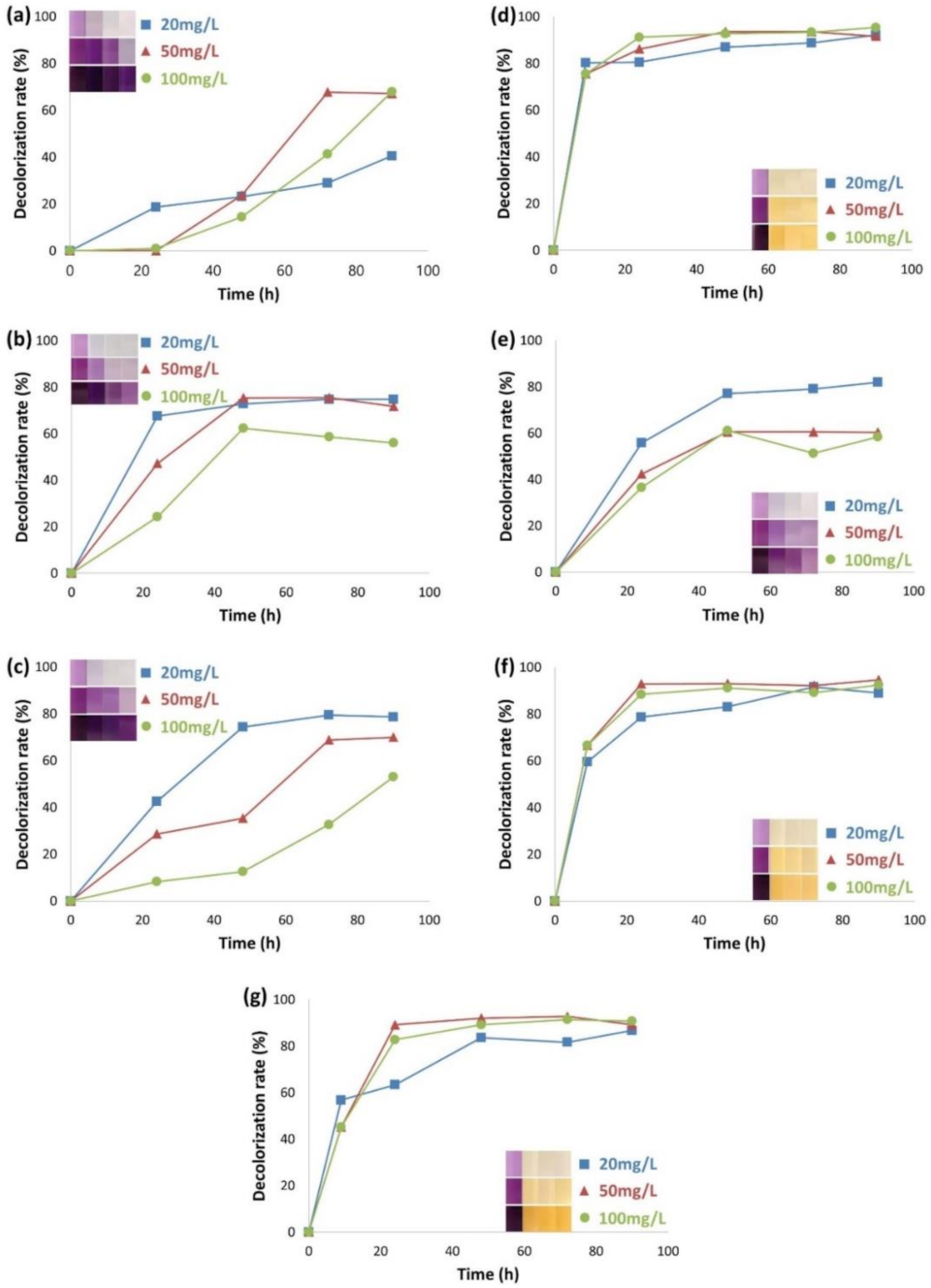
3.1. Time Courses of Dye Decolorization by Various Cultures
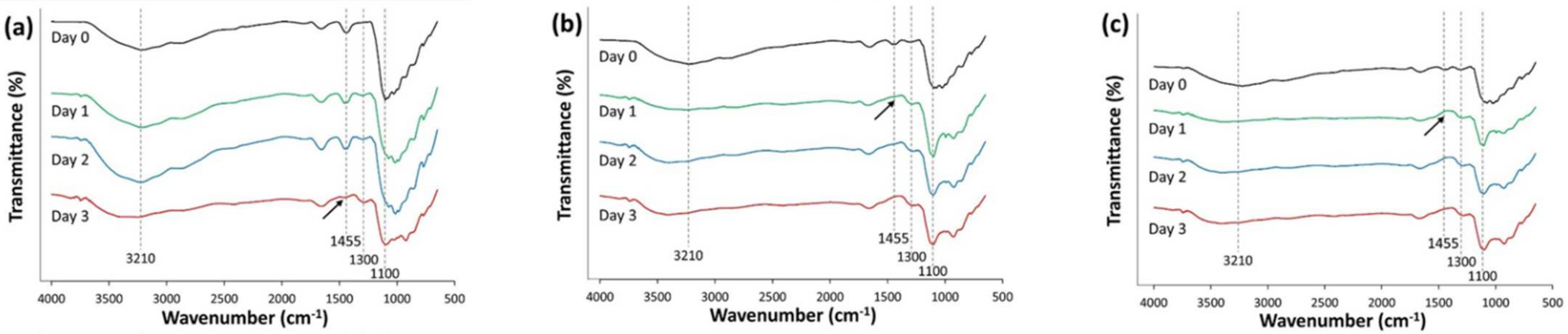
3.2. Monitoring of Organics and Biodegradation Process by Microbial Consortia for the Azo Dye AB113
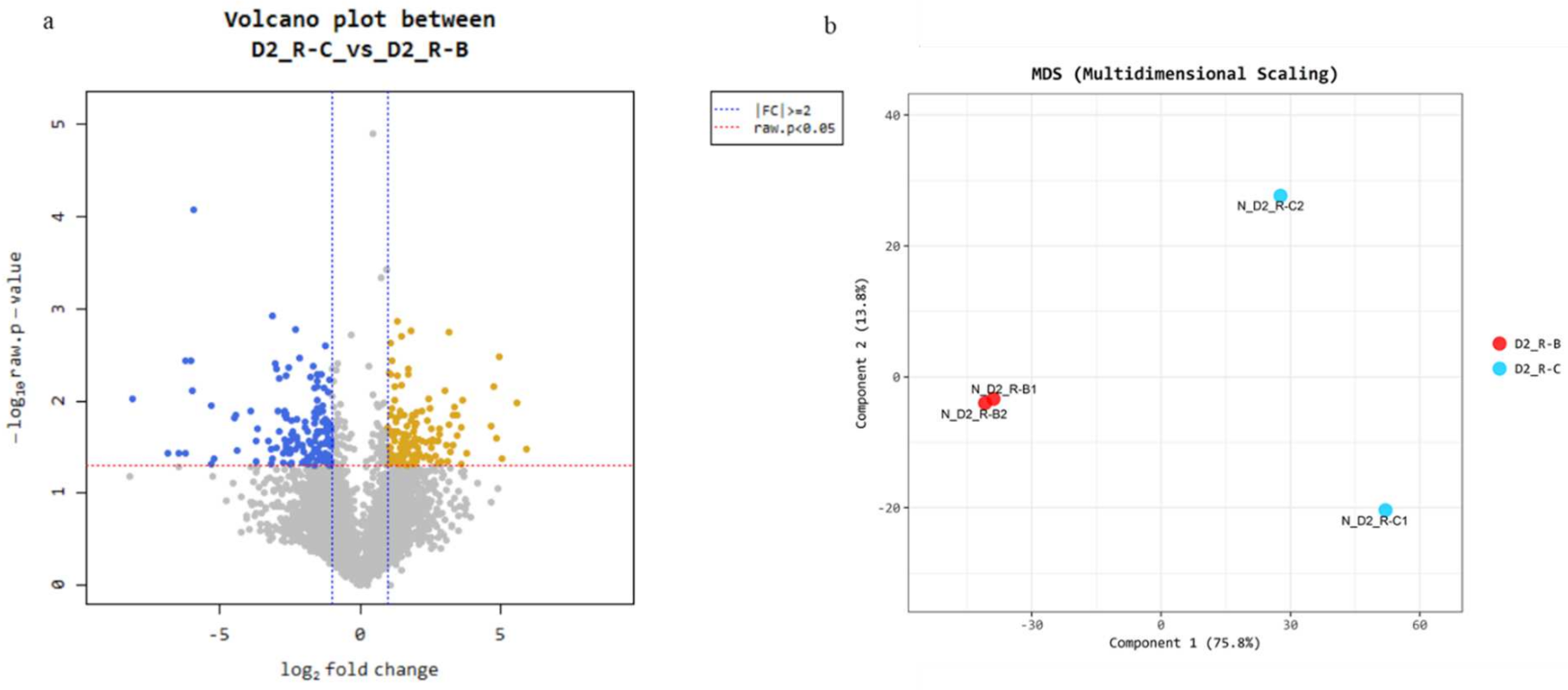
3.3. Results of Transcriptome Data and Quality Control Evaluation
3.4. Gene Expression Pattern Analysis and Clustering of DEGs
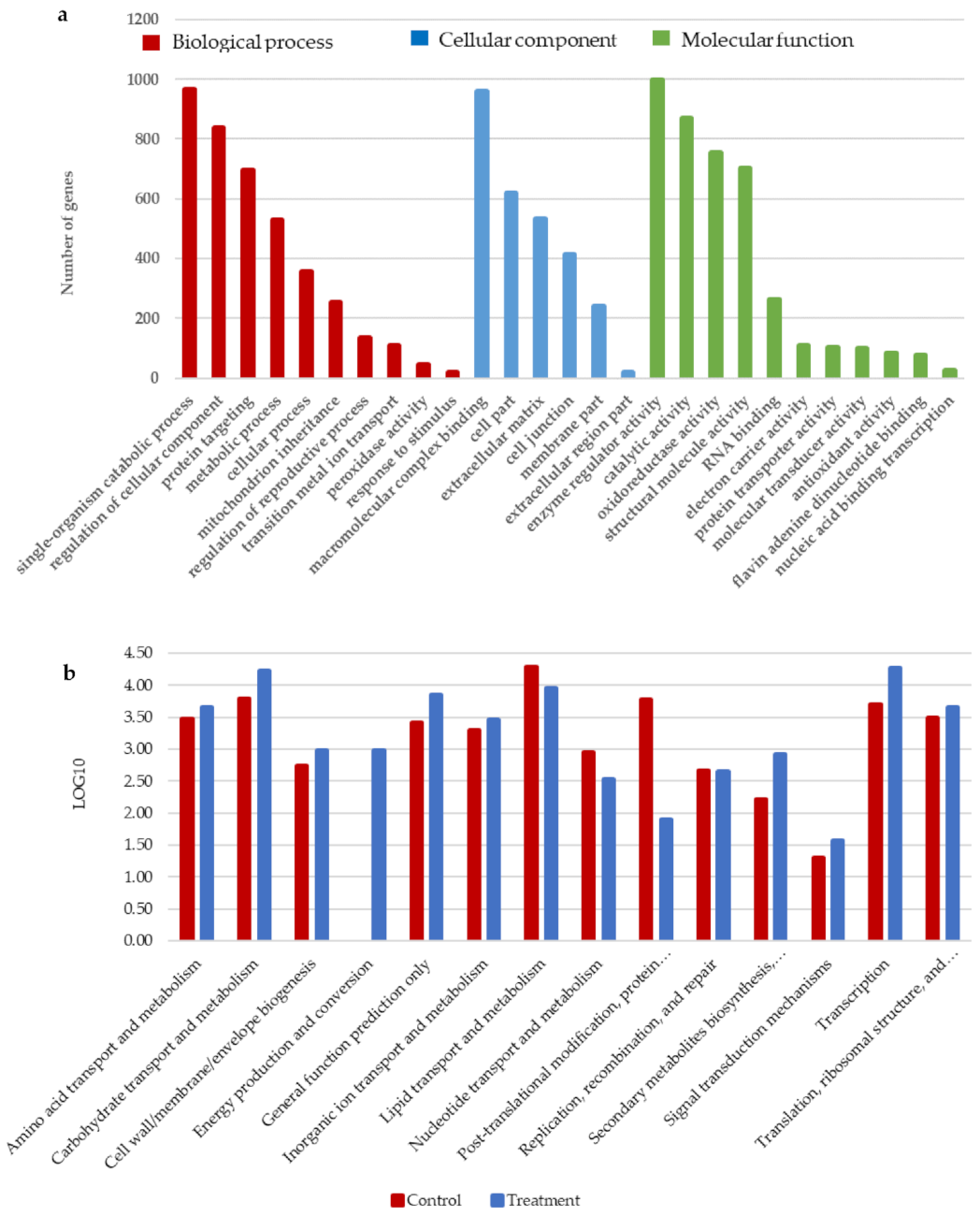
3.5. Differential Grouping of DEGs in the GO Database Analysis
3.6. Differential Grouping of DEGs and Their Annotation on EggNOG Database
3.7. Gene Expression Pattern Analysis and Clustering of DEGs
3.8. Expression Patterns of Azo Dye Degradative Enzymes and Their Metabolic Pathway Connections in the Degradation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ngulube, T.; Gumbo, J.R.; Masindi, V.; Maity, A.; Ngulube, T.; Gumbo, J.R.; Masindi, V.; Maity, A. An update on synthetic dyes adsorption onto clay based minerals: A state-of-art review. J. Environ. Manag. 2017, 191, 35–57. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Arora, S. Removal of Synthetic Textile Dyes from Wastewaters: A Critical Review on Present Treatment Technologies. Crit. Rev. Environ. Sci. Technol. 2011, 41, 807–878. [Google Scholar] [CrossRef]
- Saratale, R.; Saratale, G.D.; Chang, J.; Govindwar, S. Bacterial decolorization and degradation of azo dyes: A review. J. Taiwan Inst. Chem. Eng. 2011, 42, 138–157. [Google Scholar] [CrossRef]
- Robinson, T.; McMullan, G.; Marchant, R.; Nigam, P. Remediation of dyes in textile effluent: A critical review on current treatment technologies with a proposed alternative. Bioresour. Technol. 2001, 77, 247–255. [Google Scholar] [CrossRef]
- Khehra, M.S.; Saini, H.S.; Sharma, D.K.; Chadha, B.S.; Chimni, S. Decolorization of various azo dyes by bacterial consortium. Dye. Pigment. 2005, 67, 55–61. [Google Scholar] [CrossRef]
- Solís, M.; Solís, A.; Pérez, H.I.; Manjarrez, N.; Flores, M. Microbial decolouration of azo dyes: A review. Process Biochem. 2012, 47, 1723–1748. [Google Scholar] [CrossRef]
- Almeida, E.; Corso, C.R. Comparative study of toxicity of azo dye Procion Red MX-5B following biosorption and biodegradation treatments with the fungi Aspergillus niger and Aspergillus terreus. Chemosphere 2014, 112, 317–322. [Google Scholar] [CrossRef]
- de Miranda, R.d.C.M.; de Barros Gomes, E.; Pereira, N.; Marin-Morales, M.A.; Machado, K.M.G.; de Gusmão, N.B. Biotreatment of textile effluent in static bioreactor by Curvularia lunata URM 6179 and Phanerochaete chrysosporium URM 6181. Bioresour. Technol. 2013, 142, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Carliell, C.M.; Barclay, S.J.; Shaw, C.; Wheatley, A.D.; Buckley, C.A. The Effect of Salts Used in Textile Dyeing on Microbial Decolourisation of a Reactive Azo Dye. Environ. Technol. 1998, 19, 1133–1137. [Google Scholar] [CrossRef]
- Ayed, L.; El Ksibi, I.; Charef, A.; El Mzoughi, R. Hybrid coagulation-flocculation and anaerobic-aerobic biological treatment for industrial textile wastewater: Pilot case study. J. Text. Inst. 2020, 112, 200–206. [Google Scholar] [CrossRef]
- Amin, S.; Rastogi, R.P.; Chaubey, M.G.; Jain, K.; Divecha, J.; Desai, C.; Madamwar, D. Degradation and Toxicity Analysis of a Reactive Textile Diazo Dye-Direct Red 81 by Newly Isolated Bacillus sp. DMS2. Front. Microbiol. 2020, 11, 2280. [Google Scholar] [CrossRef]
- Alhassani, H.; Rauf, M.; Ashraf, S. Efficient microbial degradation of Toluidine Blue dye by Brevibacillus sp. Dye. Pigment. 2007, 75, 395–400. [Google Scholar] [CrossRef]
- Rajkumar, D.; Kim, J.G. Oxidation of various reactive dyes with in situ electro-generated active chlorine for textile dyeing industry wastewater treatment. J. Hazard. Mater. 2006, 136, 203–212. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Titus, A.; Gnanamani, A.; Mandal, A.; Sekaran, G. Treatment of textile wastewater by homogeneous and heterogeneous Fenton oxidation processes. Desalination 2011, 281, 438–445. [Google Scholar] [CrossRef]
- Asghar, A.; Bello, M.M.; Raman, A.A.A.; Daud, W.M.A.W.; Ramalingam, A.; Zain, S.B.M. Predicting the degradation potential of Acid blue 113 by different oxidants using quantum chemical analysis. Heliyon 2019, 5, e02396. [Google Scholar] [CrossRef] [Green Version]
- Hamza, M.; Altaf, A.A.; Kausar, S.; Murtaza, S.; Rasool, N.; Gul, R.; Badshah, A.; Zaheer, M.; Shah, S.A.A.; Zakaria, Z.A. Catalytic Removal of Alizarin Red Using Chromium Manganese Oxide Nanorods: Degradation and Kinetic Studies. Catalysts 2020, 10, 1150. [Google Scholar] [CrossRef]
- Sarkar, S.; Banerjee, A.; Halder, U.; Biswas, R.; Bandopadhyay, R. Degradation of Synthetic Azo Dyes of Textile Industry: A Sustainable Approach Using Microbial Enzymes. Water Conserv. Sci. Eng. 2017, 2, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.U.; Hinsu, A.T.; Kotadiya, R.J.; Rank, J.K.; Andharia, K.N.; Kothari, R.K. Decolorization and biodegradation of textile di-azo dye Acid Blue 113 by Pseudomonas stutzeri AK6. 3 Biotech 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Dhanve, R.S.; Shedbalkar, U.U.; Jadhav, J.P. Biodegradation of diazo reactive dye Navy blue HE2R (Reactive blue 172) by an isolated Exiguobacterium sp. RD3. Biotechnol. Bioprocess Eng. 2008, 13, 53–60. [Google Scholar] [CrossRef]
- Pourbabaee, A.; Malekzadeh, F.; Sarbolouki, M.; Najafi, F. Aerobic Decolorization and Detoxification of a Disperse Dye in Textile Effluent by a New Isolate of Bacillus sp. Biotechnol. Bioeng. 2006, 93, 631–635. [Google Scholar] [CrossRef]
- Bulgariu, L.; Escudero, L.B.; Bello, O.S.; Iqbal, M.; Nisar, J.; Adegoke, K.A.; Alakhras, F.; Kornaros, M.; Anastopoulos, I. The utilization of leaf-based adsorbents for dyes removal: A review. J. Mol. Liq. 2018, 276, 728–747. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cheng, X.; Sun, D. Autocatalysis in Reactive Black 5 biodecolorization by Rhodopseudomonas palustris W1. Appl. Microbiol. Biotechnol. 2008, 80, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, L.; Feng, L.; Chen, G.; Wang, Y.; Zhai, Z.; Zhang, Q. Exploration of the key functional strains from an azo dye degradation microbial community by DGGE and high-throughput sequencing technology. Environ. Sci. Pollut. Res. 2019, 26, 24658–24671. [Google Scholar] [CrossRef]
- Santhanarajan, A.; Sul, W.-J.; Yoo, K.-J.; Seong, H.-J.; Kim, H.-G.; Koh, S.-C. Metagenomic Insight of a Full Scale Eco-Friendly Treatment System of Textile Dye Wastewater Using Bioaugmentation of the Composite Culture CES-1. Microorganisms 2021, 9, 1503. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, S.A.; Sul, W.J.; Seong, H.J.; Rhee, C.; Ekpheghere, K.I.; Kim, I.-S.; Kim, H.-G.; Koh, S.-C. Metagenomic analysis of relationships between the denitrification process and carbon metabolism in a bioaugmented full-scale tannery wastewater treatment plant. World J. Microbiol. Biotechnol. 2019, 35, 149. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Shao, Y.; Ning, S.; Tan, L. Performance of a newly isolated salt-tolerant yeast strain Pichia occidentalis G1 for degrading and detoxifying azo dyes. Bioresour. Technol. 2017, 233, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.H.-J.; Beiko, R.G. Transfer of energy pathway genes in microbial enhanced biological phosphorus removal communities. BMC Genom. 2015, 16, 526. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Ning, S.; Zhang, X.; Shi, S. Aerobic decolorization and degradation of azo dyes by growing cells of a newly isolated yeast Candida tropicalis TL-F1. Bioresour. Technol. 2013, 138, 307–313. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Ning, S.; Shi, S.; Tan, L. Improving Azo Dye Decolorization Performance and Halotolerance of Pichia occidentalis A2 by Static Magnetic Field and Possible Mechanisms Through Comparative Transcriptome Analysis. Front. Microbiol. 2020, 11, 712. [Google Scholar] [CrossRef]
- Cheriaa, J.; Bakhrouf, A. Triphenylmethanes, malachite green and crystal violet dyes decolourisation by Sphingomonas paucimobilis. Ann. Microbiol. 2009, 59, 57–61. [Google Scholar] [CrossRef]
- Coughlin, M.F.; Kinkle, B.K.; Bishop, P.L. Degradation of azo dyes containing aminonaphthol by Sphingomonas sp. strain 1CX. J. Ind. Microbiol. Biotechnol. 1999, 23, 341–346. [Google Scholar] [CrossRef]
- Senthilvelan, T.; Kanagaraj, J.; Panda, R.C. Enzyme-Mediated Bacterial Biodegradation of an Azo Dye (C.I. Acid Blue 113): Reuse of Treated Dye Wastewater in Post-Tanning Operations. Appl. Biochem. Biotechnol. 2014, 174, 2131–2152. [Google Scholar] [CrossRef] [PubMed]
- FastQC. Available online: https://qubeshub.org/resources/fastqc (accessed on 24 June 2015).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Ali, S.S.; Jiao, H.; Mustafa, A.M.; Koutra, E.; El-Sapagh, S.; Kornaros, M.; Elsamahy, T.; Khalil, M.; Bulgariu, L.; Sun, J. Construction of a novel microbial consortium valued for the effective degradation and detoxification of creosote-treated sawdust along with enhanced methane production. J. Hazard. Mater. 2021, 418, 126091. [Google Scholar] [CrossRef]
- Zhang, J.; Chi, Y.; Feng, L. The mechanism of degradation of alizarin red by a white-rot fungus Trametes gibbosa. BMC Biotechnol. 2021, 21, 1–17. [Google Scholar] [CrossRef]
- Russ, R.; Rau, J.; Stolz, A. The Function of Cytoplasmic Flavin Reductases in the Reduction of Azo Dyes by Bacteria. Appl. Environ. Microbiol. 2000, 66, 1429–1434. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, S.; Khalid, A.; Arshad, M.; Mahmood, T.; Crowley, D. Detoxification of azo dyes by bacterial oxidoreductase enzymes. Crit. Rev. Biotechnol. 2015, 36, 639–651. [Google Scholar] [CrossRef]
- Sekar, S.; Mahadevan, S.; Shanmugam, B.K.; Mandal, A.B. Bioenergetics and pathway of acid blue 113 degradation by Staphylococcus lentus. Biotechnol. Prog. 2012, 28, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Guo, J.; Sun, G. Biodegradation of textile azo dye by Shewanella decolorationis S12 under microaerophilic conditions. Appl. Microbiol. Biotechnol. 2007, 76, 719–726. [Google Scholar] [CrossRef]
- Matsuo, M.; Endo, T.; Asada, K. Isolation of a novel NAD(P)H-quinone oxidoreductase from the cyanobacterium Synechocystis PCC6803. Plant Cell Physiol. 1998, 39, 751–755. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.M.; Wright, C.M.; John, G.H. Identification, Isolation and characterization of a novel azoreductase from Clostridium perfringens. Anaerobe 2012, 18, 229–234. [Google Scholar] [CrossRef]
- Gong, Y.; Liu, W.; Huang, X.; Hao, L.; Li, Y.; Sun, S. Antifungal Activity and Potential Mechanism of N-Butylphthalide Alone and in Combination with Fluconazole Against Candida albicans. Front. Microbiol. 2019, 10, 1461. [Google Scholar] [CrossRef]
- Bürger, S.; Stolz, A. Characterisation of the flavin-free oxygen-tolerant azoreductase from Xenophilus azovorans KF46F in comparison to flavin-containing azoreductases. Appl. Microbiol. Biotechnol. 2010, 87, 2067–2076. [Google Scholar] [CrossRef]
- Chen, H.; Feng, J.; Kweon, O.; Xu, H.; E Cerniglia, C. Identification and molecular characterization of a novel flavin-free NADPH preferred azoreductase encoded by azoB in Pigmentiphaga kullae K24. BMC Biochem. 2010, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Wang, R.-F.; E Cerniglia, C. Molecular cloning, overexpression, purification, and characterization of an aerobic FMN-dependent azoreductase from Enterococcus faecalis. Protein Expr. Purif. 2004, 34, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Demanèche, S.; Meyer, C.; Micoud, J.; Louwagie, M.; Willison, J.C.; Jouanneau, Y. Identification and Functional Analysis of Two Aromatic-Ring-Hydroxylating Dioxygenases from a Sphingomonas Strain That Degrades Various Polycyclic Aromatic Hydrocarbons. Appl. Environ. Microbiol. 2004, 70, 6714–6725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leys, N.M.E.J.; Ryngaert, A.; Bastiaens, L.; Verstraete, W.; Top, E.M.; Springael, D. Occurrence and Phylogenetic Diversity of Sphingomonas Strains in Soils Contaminated with Polycyclic Aromatic Hydrocarbons. Appl. Environ. Microbiol. 2004, 70, 1944–1955. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.B.; Shaikh, S.; Jain, K.R.; Desai, C.; Madamwar, D. Polycyclic Aromatic Hydrocarbons: Sources, Toxicity, and Remediation Approaches. Front. Microbiol. 2020, 11, 2675. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Nwinyi, O.C.; Ajayi, O.; Amund, O.O. Degradation of polynuclear aromatic hydrocarbons by two strains of Pseudomonas. Braz. J. Microbiol. 2016, 47, 551–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Tao, X.; Jiang, Y.; Khan, A.; Wu, Q.; Xuanyu, T.; Wu, D.; Chen, Y.; Ling, Z.; Liu, P.; et al. The naphthalene catabolic protein NahG plays a key role in hexavalent chromium reduction in Pseudomonas brassicacearum LZ-4. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Costa, D.M.; Gómez, S.V.; de Araújo, S.S.; Pereira, M.S.; Alves, R.B.; Favaro, D.C.; Hengge, A.C.; Nagem, R.A.; Brandão, T.A. Catalytic mechanism for the conversion of salicylate into catechol by the flavin-dependent monooxygenase salicylate hydroxylase. Int. J. Biol. Macromol. 2019, 129, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Díaz, E.; Ferrández, A.; Prieto, M.A.; García, J.L. Biodegradation of Aromatic Compounds by Escherichia coli. Microbiol. Mol. Biol. Rev. 2001, 65, 523–569. [Google Scholar] [CrossRef] [Green Version]
- Travkin, V.M.; Solyanikova, I.P. Salicylate or Phthalate: The Main Intermediates in the Bacterial Degradation of Naphthalene. Processes 2021, 9, 1862. [Google Scholar] [CrossRef]
- Mishra, A.K.; Baek, K.-H. Salicylic Acid Biosynthesis and Metabolism: A Divergent Pathway for Plants and Bacteria. Biomolecules 2021, 11, 705. [Google Scholar] [CrossRef]
- Higson, F.K.; Focht, D.D. Degradation of 2-methylbenzoic acid by Pseudomonas cepacia MB2. Appl. Environ. Microbiol. 1992, 58, 194–200. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Identification | Best Match (Closest Strain) | |||||
|---|---|---|---|---|---|---|---|
| Name | Strain | Authors | Accession | Similarity (%) | Mismatch/Total nt | ||
| B-1 | Mesorhisobium sp. | AVBN_s (Mesorhisobium sp.) | NBIMC_P2-C1 | Lo, S.C et al., 2014 | KF040403, AVBN01000018 | 100 | 0/1406 |
| B-3 | Mesorhisobium haukuii | IAM 14158 | (Chen et al., 1991) Jarvis et al., 1997 | D12797 | 98.42 | 22/1399 | |
| B-2 | Sphingomonas melonis B-2 | Sphingomonas melonis | DAPP-PG 224 | Buonaurio at al., 2002 | KB900605 | 100 | 0/1409 |
| Y-1 | Apiotrichum mycotoxinivorans | Apiotrichum mycotoxinivorans | CBS: 10094 | Vu, D at al., 2016 | KY109958 | 99 | 2/603 |
| Y-2 | Meyerozyma guilliermondi | Meyerozyma guilliermondi | Gj8-2 | Han, S. 2016 | KU316708 | 99 | 1/578 |
| Cultures * | Species of Cultures Grown in Culture | Inoculation Amount (%) | Applied Conc. of AB 113 (mg L−1) |
|---|---|---|---|
| B-1 | Mesorhizobium sp. | 5 | 20, 50, 100 |
| B-2 | Sphingomonas melonis | 5 | 20, 50, 100 |
| Y-1 | Apiotrichum mycotoxinivarans | 5 | 20, 50, 100 |
| Y-2 | Meyerozyma guillermondi | 5 | 20, 50, 100 |
| B-C a | Mesorhizobium sp. | 2.5 | 20, 50, 100 |
| Sphingomonas melonis | 2.5 | ||
| Y-C b | Apiotrichum mycotoxinivarans | 2.5 | 20, 50, 100 |
| Meyerozyma guillermondi | 2.5 | ||
| T-C c | Mesorhizobium sp. | 1.25 | 20, 50, 100 |
| Sphingomonas melonis | 1.25 | ||
| Apiotrichum mycotoxinivarans | 1.25 | ||
| Meyerozyma guillermondi | 1.25 |
| GO Term | Enzymes | Gene Id | Up-Regulated | Coverage (%) | E-Value |
|---|---|---|---|---|---|
| Catalytic activity | catalase HPII | gene2971 | 4.99 | 98.13 | 0 |
| Oxidoreductase activity | NADH quinone oxidoreductase | gene2085 | 5.38 | 94 | 7 × 10−1 |
| gene2088 | 8.30 | 95.68 | 1 × 10−8 | ||
| gene2090 | 4.88 | 96.57 | 5 × 10−3 | ||
| gene2095 | 10.52 | 98.04 | 8 × 10−3 | ||
| gene2099 | 11.64 | 89.64 | 0 | ||
| Catalytic activity | N-acetyltransferase | gene285 | 8.83 | 84.88 | 5 × 10−1 |
| gene680 | 2.06 | 94.4 | 9 × 10−4 | ||
| gene990 | 2.31 | 94.66 | 0 | ||
| gene1312 | 3.11 | 84.75 | 2 × 10−6 | ||
| gene1850 | 3.84 | 99.31 | 0 | ||
| gene2426 | 5.32 | 99.55 | 0 | ||
| Metabolic process | aromatic ring-hydroxylating dioxygenase | gene1111 | 10.69 | 98.12 | 5 × 10−5 |
| gene2173 | 11.85 | 87.36 | 1 × 10−9 | ||
| gene2473 | 8.88 | 85.01 | 3 × 10−1 | ||
| GO Term | Enzymes | Gene Id | Down-Regulated | Coverage | E-Value |
| Heme binding | cytochrome P450 | gene3695 | 3.17 | 35.58 | 8 × 10−1 |
| gene3489 | 2.10 | 98.29 | 6 × 10−6 | ||
| gene1577 | 3.44 | 96.36 | 1 × 10−1 | ||
| Oxidoreductase activity | NADH flavin reductase | gene1341 | 2.89 | 99.52 | 3 × 10−7 |
| Catalytic activity | 3-hydroxybutyryl-CoA dehydrogenase | gene953 | 2.48 | 93.1 | 2 × 10−9 |
| salicylate hydroxylase | gene3343 | 2.86 | 49.36 | 0.0003 | |
| salicylaldehyde dehydrogenase | gene420 | 3.56 | 96.61 | 8 × 10−7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santhanarajan, A.-E.; Rhee, C.; Sul, W.J.; Yoo, K.; Seong, H.J.; Kim, H.-G.; Koh, S.-C. Transcriptomic Analysis of Degradative Pathways for Azo Dye Acid Blue 113 in Sphingomonas melonis B-2 from the Dye Wastewater Treatment Process. Microorganisms 2022, 10, 438. https://doi.org/10.3390/microorganisms10020438
Santhanarajan A-E, Rhee C, Sul WJ, Yoo K, Seong HJ, Kim H-G, Koh S-C. Transcriptomic Analysis of Degradative Pathways for Azo Dye Acid Blue 113 in Sphingomonas melonis B-2 from the Dye Wastewater Treatment Process. Microorganisms. 2022; 10(2):438. https://doi.org/10.3390/microorganisms10020438
Chicago/Turabian StyleSanthanarajan, Aalfin-Emmanuel, Chaeyoung Rhee, Woo Jun Sul, Keunje Yoo, Hoon Je Seong, Hong-Gi Kim, and Sung-Cheol Koh. 2022. "Transcriptomic Analysis of Degradative Pathways for Azo Dye Acid Blue 113 in Sphingomonas melonis B-2 from the Dye Wastewater Treatment Process" Microorganisms 10, no. 2: 438. https://doi.org/10.3390/microorganisms10020438